1 Схема Структурные компоненты бактериальной клетки Обязательные
 Скачать 4.86 Mb. Скачать 4.86 Mb.
|

|
133.Препарат «Лецитиназная активность стафилококка» 1 - бактериологический метод, получение изолированных колоний на плотной питательной среде (1 этап); 1этап 1день:материал-гной больного,проводят высев исследуемого материала в чашку петри с МПА с целью получения изолированных колоний(37с,24ч).2 день:изучение изолир.колоний выросших на чашке Петри с МПА,и пересев грам - колоний на скошенный агар для получения чистых аэробных культур пробирку поместить в термостат(37с) 3день:проверяем чистая ли культура выросла:макроскопически оцениваем однородность роста культуры на скошенном агаре и микроскопически для этого берут материал из разных мест выросшей на агаре культуры,готовят мазок и окрашивают по ГРаму,если клетки одинаковые по форме и окраске то это чистая культура 2 - желточно-солевой агар; 3 - образование «радужных» ореолов, зон преломления света вокруг колоний; 4 – биохимическая(делают посев чистой культуры уколом в столбик с маннитом через 18-24 часа измен окраска индикатора) и по ферментам агрессии;выполняют посев на цитратную плазму,после икубации 18-24 часа плазма коагулировалась->коагулаза+;наличие гемолизина появление гемолиза 5 –идентификация чистой культуры по ферментам агрессси: расщепление лецитина и вителлина под действием ферментативного комплекса лецитовителлазы (лицетиназы) 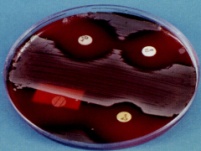 1.34.Антнбиотикограмма, способ дисков на кровяном агаре 1 - бактериологический метод на этапе изучения чистой культуры; 2 - диффузионный способ определения чувствительности к дискам с антибиотиками на плотной среде; 3 - размеры зоны задержки роста культуры или её отсутствие; 4 - можно воспользоваться определением минимальной подавляющей концентрации препаратов (МПК), использовать др. Способы определения чувствительности к лекарственным препаратам, проводить определение генов резистентности с помощью ПЦР; 5 - посев выполнен на кровяном агаре; подобная постановка исследования используется при опенке чувствительности гемофильных и анаэробных бактерий, не растущих на обычных питательных средах  1.35. Антибиотикограмма, кассетный микрометод 1 - бактериологический метод на этапе изучения чистой культуры или первичный посев материала (экспресс-методика); 2 - диффузионный способ определения чувствительности к антибиотикам на плотной среде, которая распределена в лунки пластиковой кассеты с разными препаратами и/или разными концентрациями препаратов; 3 - отсутствие признаков роста культуры или её рост; 4 - можно воспользоваться способом дисков, использовать др. Способы определения чувствительности к лекарственным препаратам, проводить определение генов резистентности с помощью ПЦР; 5 - модифицированная методика позволяет определять как чувствительность отдельных штаммов, так и проводить определение МПК; в данном случае посев выполнен на кровяном агаре; подобная постановка исследования используется при оценке чувствительности гемофильных и анаэробных бактерий, не растущих на обычных питательных средах 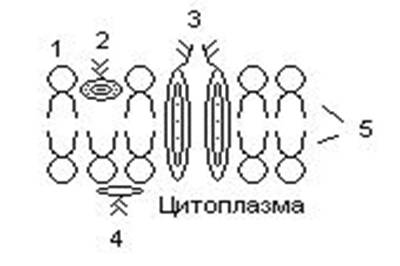 1.36.Схема «Строение цитоплазматической мембраны прокариотов». 1 - фосфолипиды, определяют непроницаемость за счёт гидрофобных липидных «головою); 2 - поверхностные белки с углеводными цепочками (адгезины и т.п., определяют рецепторную функцию); 3 - интегральные белки (транслоказы, порины и т.п. - определяют избирательную проницаемость клетки, транспортную функцию, участие в процессах питания, биосинтезе, делении клетки); 4 - дыхательные и др. Внутриклеточные ферменты (определяют высокую метаболическую активность прокариотической клетки, участвуют в энергетическом обмене (АТФ), процессах репликации ДНК, делении клетки); 5 - билипидный слой из молекул фосфолипидов с вкраплениями поверхностных и интегральных белковых молекул. 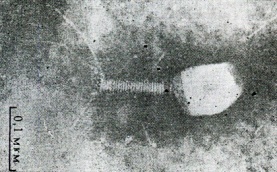 1.37. Фотография «Бактериофаг под электронным микроскопом» 1 - бактериофаг (Т2) 2 - состоит из головки, отростка, базальной пластины, шипов и нитей-филаментов; головка фага покрыта белковым чехлом, в котором полипептидные субъединицы расположены по кубическому типу симметрии, внутри головки находится гигантская непрерывная двунитевая молекула ДНК, суперсперилизованая за счёт внутренних белков фага: спермина и путресцина; в дистальной части отростка расположен фермент лизоцим, необходимый для разрушения глихозндных связей пептидогли кана, входящего в состав клеточных стенок бактерий 3 - репродукция в бактериальной клетке; бактериофаги могут быть вирулентными и умеренными; вирулентные фаги характеризуются тем, что цикл их развития в бактериальной клетке обязательно завершается выходом фагов из бактерии и гибелью бактериальной клетки, т.е. Наблюдается продуктивная форма инфекции (примером является бактериофаг кишечных бактерий Т2) 4 - адгезия фагов на специфических рецепторах клеточной стенки бактерий с помощью нитей и шипов отростка; сокращение белкового чехла отростка и инъекция нуклеиновой кислоты (ДНК) внутрь бактериальной клетки; синтез ранних фаговых белков, которые запускают синтез фаговой ДНК 5 - приготовление препаратов для диагностики (фаготипирование стафилококка) или лечения бактериальных инфекций (стафилококковый, брюшнотифозный, холерный, чумной фаги) 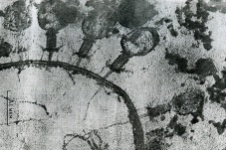 1.38 Фотография «Атака клетки бактериофагами под электронным микроскопом» 1 - адсорбция бактериофагов на поверхности бактериальной клетки 2 - клетка и бактериофаги, прикрепившиеся к клеточной стенке, а также нити деспирализованной ДНК фагов в цитоплазме клетки 3 - внутриклеточная репродукция вирулентных фагов 4 - адгезия фагов на специфических рецепторах клеточной стенки бактерий с помощью нитей и шипов отростка; сокращение белкового чехла отростка и инъекция нуклеиновой кислоты (ДНК) внутрь бактериальной клетки; синтез ранних фаговых белков, которые запускают синтез фаговой ДНК и далее - поздних фаговых белков 5 - цикл литической инфекции бактериальных клеток вирулентным фагом (в отличие от лизогенного цикла умеренного фага); в медицинской практике используется для диагностики (фаготипирование стафилококка) или лечения бактериальных инфекций (стафилококковый, брюшнотифозный, холерный, чумной фаги) 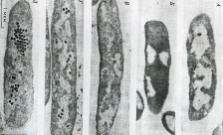 139. Фотография «Репродукция бактериофага в клетке под электронным микроскопом» 1 - разъединенный синтез отдельных структурных элементов вируса с последующей их сборкой (репродукция вирулентного бактериофага Т2 в кишечной палочке) 2 - бактериальная клетка, содержащая фаговые частицы 3 - внутриклеточная репродукция вирулентных фагов (морфогенез фага вдет по трем основным направлениям, независимо приводящим к формированию головок, отростков и нитей, которые затем объединяются, образуя зрелые фаговые частицы; процесс контролируют более 60 фаговых генов) 4 - А-Б-В - скрытый период сборки фаговых компонентов (14 мин); Г- появление первых собранных фагов (15 мин); Д - начало «выхода» фагов из клетки (40 мин); наряду с образовавшимися зрелыми частицами фага в цитоплазме бактерий накапливается свободный фаговый лизоцим, являющийся поздним фаговым белком, который изнутри истончает клеточную стенку бактерий и она разрывается под действием внутреннего осмотического давления 5 - Образуются новые фаги (до 300 фагов), а бактериальная клетка погибает 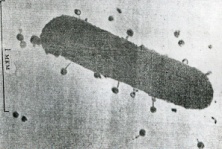 1.40. Фотография «Адсорбция бактериофагов на бактериальной клетке под электронным микроскопом» 1 - адсорбция бактериофагов на поверхности бактериальной клетки 2 - клетка и множество бактериофагов, находящихся вокруг и прикрепившихся к клеточной стенке бактерии 3 - внутриклеточная репродукция вирулентных фагов — адгезия фагов на специфических рецепторах клеточной стенки бактерий; инъекция нуклеиновой кислоты (ДНК) внутрь бактериальной клетки; синтез ранних фаговых белков, которые запускают синтез фаговой ДНК и далее - поздних фаговых белков 5 - используется для диагностики (фаготипирование стафилококка) или лечения бактериальных инфекций (стафилококковый, брюшнотифозный, холерный, чумной фаги) 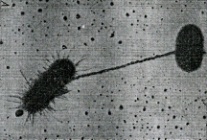 1.41. Фотография «Формирование коньюгативного мостика между бактериями» 1- процесс конъюгации между jf+ и F- бактериями 2 - на электронной фотографии - клетки донора и реципиента (F-), хорошо видна половая пиля; половые пили представляют нитевидные образования полые внутри, образуемые белком пилином (количество пилей колеблется в пределах 1-10) 3 - конъюгация является вариантом рекомбинативной изменчивости у бактерий; в процессе конъюгации от клетки донора к клетке реципиенту осуществляется перенос фактора фертильности (F-плазмида), а также и других плазмид и генов хромосомы (плазмиды резистентности к антибиотикам, tox-гены и т.п.). Бактерии доноры, чтобы иметь возможность осуществлять конъюгацию, должны обладать дополнительным генетическим материалом - Р плазмидой (от слова фертильность - плодовитость). F плазмида имеет синонимы - половой фактор, фактор фертильности, фактор плодовитости, F фактор; она представляет собой двунитевую ковалентно замкнутую молекулу ДНК, которая по молекулярной массе значительно меньше хромосомы бактериальной клетки; в составе F фактора имеются гены, отвечающие не только за репликацию, но и кодирующие синтез половых пилей (ворсинок) на поверхности бактериальной клетки. 4 - после того как произошла коагтрегация двух бактерий и половые пили соединили обе конъюгирующие клетки, происходит сокращение пилей в результате чего в течение нескольких минут бактерия реципиент подтягивается вплотную к донору и клеточные стенки обеих бактерий входят в тесный контакт, формируя цитоплазматический мостик; далее осуществляется репликация плазмиды и перенос генетической информации 5 - передача генов резистентности к антибиотикам, кодирующих синтез ферментов, факторов вирулентности, ускорение процесса репродукции у бактерий в случае формирования Hfr-пслеток 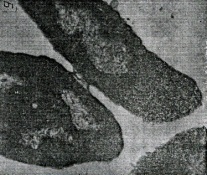 1.42. Фотография «Конъюгация у бактерий под электронным микроскопом» 1 - процесс конъюгации между F+ и F- бактериями 2 - на электронной фотографии - клетки донора (F+) и реципиента (F-), хорошо виден цитоплазматический мостик; 3 - передача генетического материала в редких случаях может происходить по половым пилям, но все-таки в подавляющем большинстве случаев она происходит по цитоплазматичеекому мостику 4 - существуют два состояния полового фактора: свободное нахождение в цитоплазме, и тогда бактерии обозначаются F* и интегрированное в хромосому. В последнем случае бактерии обозначаются Hfr, т.е.обладающис высокой частотой рекомбинации (от англ.: hight frequence of recombinantion). Если половой фактор находится в свободном состоянии в цитоплазме, то в клетку реципиент передается только его генетический материал. 5 - передача генов резистентности к антибиотикам, кодирующих синтез ферментов, факторов вирулентности, ускорение процесса репродукции у бактерий в случае формирования Hfr-гклеток  1.43. Схема «Формирование репликативной вилки хромосомы» 1 - схема репликации ДНК (формирование репликативной вилки) 2 - процесс расплетания нитей ДНК и участвующие в нём ферменты 3 - синтез ДНК на обеих матричных цепях идет в направлении от 5' к 3' концам. На ведущей нити ДНК он происходит непрерывно, а на отстающей прерывисто в виде фрагментов Оказаки, каждый из которых включает 1000-2000 нуклеотидов 4 - образование репликативной вилки ДНК начинается в определённом участке ДНК (ориджин) при участии фермента ДНК-гиразы; дальнейшее формирование репликативной вилки обеспечивают ферменты хелнказы. Одна хеликаза передвигается по ведущей нити ДНК, а другая по отстающей. Кроме хеликаз в формировании репликативной вилки участвуют SSB - белки, специфически связывающиеся с каждой из расплетенных нитей ДНК. Они предотвращают преждевременное их соединение. Вновь синтезированные нити ДНК всегда содержат на 51 концах несколько рибонуклеотидов, т.е. Синтез ДНК начинается с синтеза короткой цепи РНК, связанной с матрицей ДНК. Эту РНК-затравку образует специальный фермент, который называется ДНК-праймаза (праймер - затравка). Затем синтез ДНК продолжается благодаря присоединению к РНК-затравке дезокеирибонуклеотидов с помощью ДНК-полимиразы III. Удаление РНК-затравки и застройка брешей осуществляется ДНК-полимеразой I. На отстающей нити ДНК, остающиеся между двумя соседние фрагментами Оказаки, однонитевые разрывы заделывает ДНК-лигаза. Раскручивание узлов, образующихся перед репликативной вилкой сверхсперилизованными нитями ДНК, осуществляет фермент топоизомераза. Энергия, необходимая для расплетания нитей ДНК поставляется в результате гидролиза АТФ 5 - делению бактериальной клетки обязательно должна предшествовать репликация ДНК 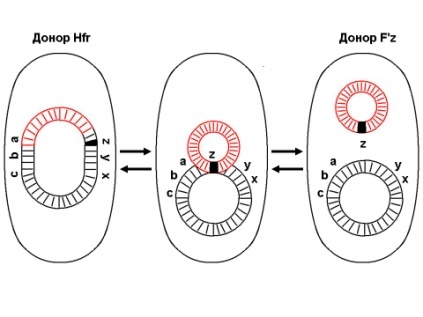 1.44. Схема «Конъюгация у бактерий. Формирование F рекомбинантов» 1 - конъюгация у бактерий: формирование Ррекомбинантов 2 - донор Hfr, вычленение плазмиды с геном z, донор f'z 3 - существуют два состояния полового фактора: свободное нахождение в цитоплазме, и тогда бактерии обозначаются F*" и интегрированное в хромосому - обозначаются Hfr, т.е.обладающие высокой частотой рекомбинации (от англ.: hight frequence of recombinantion). Бели половой фактор находится в свободном состоянии в цитоплазме, то в клетку реципиент передается только его генетический материал 4 - У Hfr бактерий может происходить выход полового фактора из состава хромосомы в цитоплазму, в результате возникает клон F* клеток. В некоторых случаях при выходе F фактора он может включить расположенный рядом ген бактериальной хромосомы (путем рекомбинации). В таких случаях рядом с обозначением полового фактора дается символ соответствующего гена (z). Следовательно, особенностью Hfr бактерий является передача генов собственной хромосомы в клетку реципиент. Например, fmac, т.е. В составе полового фактора находится лактозный оперон, отвечающий за утилизацию лактозы. В результате бактерии имеют делецию, соответствующую лактозному гену, являющегося уже составной частью полового фактора. Если в состав полового фактора войдут гены, жизненно необходимые бактерии, то потеря его клеткой может привести к ее гибели. Поскольку половые факторы могут включаться в различные участки бактериальной хромо сомы, то F плазмида может содержать различные гены. 5 - передача генов резистентности к антибиотикам, кодирующих синтез ферментов, факторов вирулентности, ускорение процесса репродукции у бактерий в случае формирования Hfr-гклеток  1.45. Схема «Этапы конъюгации у бактерий» 1 - конъюгация у бактерий 2 - конъюгирующие клетки, этапы репликации F плазмиды с последующим формированием из F- клетки 3- конъюгация является вариантом рекомбинативной изменчивости у бактерий; в процессе конъюгации от клетки донора к клетке реципиенту осуществляется перенос фактора фертильности (F-плазмида), а также и других плазмид и генов хромосомы (плазмиды резистентности к антибиотикам, tox-гены и т.п.). Бактерии доноры, чтобы иметь возможность осуществлять конъюгацию, должны обладать дополнительным генетическим материалом - F плазмидой (от слова фертильность - плодовитость). F плазмида имеет синонимы - половой фактор, фактор фертильности, фактор плодовитости, F фактор; она представляет собой двунитевую ковалентно замкнутую молекулу ДНК, которая по молекулярной массе значительно меньше хромосомы бактериальной клетки; в составе F фактора имеются гены, отвечающие не только за репликацию, но и кодирующие синтез половых пилей (ворсинок) на поверхности бактериальной клетки. 4 - расплетание ДНК плазмиды; переход нити ДНК из F+ донора с 5* конца в F Реципиент; достраивание комплементарной нити; формирование F плазмиды 5 - передача генов резистентности к антибиотикам, кодирующих синтез ферментов, факторов вирулентности и т.п. 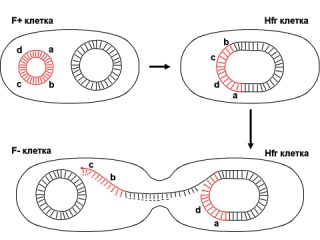 1.46. Схема «Конъюгация у бактерий. Формирование Hfr рекомбинантов» 1 - конъюгация у бактерий: формирование Hfr рекомбинантов 2 - донор F+, Hfr клетка, деление Hfr рекомбинантов 3 - существуют два состояния полового фактора: свободное нахождение в цитоплазме, и тогда бактерии обозначаются F+ и интегрированное в хромосому - обозначаются Hfr, т.е.обладающие высокой частотой рекомбинации (от англ.: hight frequence of recombinantion). Если половой фактор находится в свободном состоянии в цитоплазме, то в клетку реципиент передается только его генетический материал 4 - У Hfr штаммов (у них половой фактор включен в хромосому) конъюгация начинается с разрезания одной нити ДНК в области включенного полового фактора. Поэтому в клетку реципиент входит сначала небольшой участок ДНК F фактора наподобие "локомотива", а затем гены хромосомы донора (вагоны, которые тянет "локомотив"). На нити ДНК, входящей в реципиент, синтезируется комплементарная ей нить. Затем перешедшие в клетку реципиент гены донора вступают в рекомбинацию с генами реципиента, замещая свои аллели. У Hfr бактерий может происходить выход полового фактора из состава хромосомы в цитоплазму, в результате возникает клон F* клеток. 5 - передача генов резистентности к антибиотикам, кодирующих синтез ферментов, факторов вирулентности, ускорение процесса репродукции у бактерии в случае формирования Hfr-гклеток 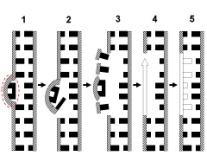 |