Тема 6. ФІЗІОЛОГІЯ КРОВООБІГУ. 1. Загальна характеристика системи кровообігу. Фактори, які забезпечують рух крові по судинах, його спрямованість та безперервність
 Скачать 2.43 Mb. Скачать 2.43 Mb.
|

|
фізіологія кровообігу. 1. Загальна характеристика системи кровообігу. Фактори, які забезпечують рух крові по судинах, його спрямованість та безперервність. СИСТЕМА КРОВООБІГУ   ВИКОНАВЧІ ОРГАНИ МЕХАНІЗМИ РЕГУЛЯЦІЇ Нервові Гуморальні   Серце Судини  Хвилинний об’єм крові (ХОК), який є адекватним потребам організму В залежності від потреби організму ХОК може змінюватися у дорослої людини від 5 л/хв (спокій) до 30 л/хв (стан фізичного навантаження у добре тренованого спортсмена). Причиною руху крові по судинам (та через камери серця) є різниця (градієнт) тисків, що створюється завдяки: - нагнітальній (насосній) функції серця; - тонусу судин. Односторонність току крові забезпечується, завдяки певному “напрямку” градієнта тиску в системі (від аорти до порожнистих вен градієнт біля 100 мм рт. ст., від легеневого стовбура до легеневих вен – близько 20 мм рт. ст. в такому напрямку рухається кров судинами). Якщо в системі виникає ситуація, коли кров за градієнтом тиску має рухатися в протилежному напрямку (наприклад, вени, що розташовані нижче рівня серця при вертикальній позі людини), зворотньому руху крові перешкоджають клапани. Аналогічно функціонують клапани серця: перешкоджають руху крові в передсердя із шлуночків під час систоли (скорочення) останніх; з аорти та легеневого стовбура в шлуночки під час їх діастоли (розслаблення). Причиною безперервного руху крові в системі є судини компресійної камери (камери стиснення). Це аорта та крупні артеріальні судини, в стінках яких переважають еластичні волокна. Внаслідок цього вони досить пружні та здатні до розтягу. Під час періоду вигнання (частина систоли шлуночків) вони розширюються (при цьому частина енергії скорочення серця переходить в енергію напруження еластичних волокон цих судин). По закінченню вигнання судини компресійної камери стискуються (енергія напруження еластичних волокон судин переходить в енергію руху крові) та проштовхуюють кров в переферійні судини. Серцевий цикл триває в стані спокою 0,8 с, період вигнання – 0,25 с. Тобто, протягом 0,55 с кров в судини серцем не виштовхується, але рухається по ним безперервно, завдяки судинам компресійної камери. 2. Автоматія серця. Градієнт автоматії. Дослід Станіуса. Автоматія – здатність збуджуватися (генерувати ПД) без дії зовнішнього подразника (інакше – здатність до самозбудження). Ця здатність є у структурах серця, побудованих з атипічних кардіоміоцитів, а саме, в стимульному комплексі (провідній системі) серця:
Ці елементи провідної системи серця носять назву центрів автоматії й мають певний порядок. Наприклад, пазухово-передсердний вузол – центр першого порядку, передсердно-шлуночковий – другого і т.д. Градієнт автоматії – зменшення ступеня автоматії елементів провідної системи серця в напрямку від пазухово-передсердного вузла до волокон Пуркіньє. Ступінь автоматії характеризує частота, з якою центр автоматії генерує імпульси збудження (ПД). Найчастіше імпульси генерує пазухово-передсердний вузол – від 50-60 імп/хв і більше. Передсердно-шлуночковий вузол генерує ПД з меншою частотою – 30-40 імп/хв, пучок Гіса – 20-30 імп/хв і т.д. У здорової людини серце збуджується і скорочується в ритмі, що відповідає частоті генерування ПД пазухово-передсердним вузлом. Тобто, він є водієм ритму серця, або пейсмекером (центрів автоматії багато; водієм ритму в певний момент часу є лише один із центрів – той, який визначає частоту збудження і скорочення шлуночків серця). Наявність центрів автоматії робить роботу серця надійною – якщо з роботи виключається пазухово-передсердний вузол як водій ритму серця, його функції бере на себе центр автоматії другого порядку, тобто передсердно-шлуночковий вузол. В нормі більш високу частоту генерації ПД має пазухово-передсердний вузол, його імпульси досягають інших відділів (водіїв ритму нижчого порядку) раніше, ніж там відбудеться спонтанна деполяризація. Мініатюра: “Жабяче серце або дослід Станіуса”. Дійові особи: Жаба – хтива царівна з потворною пикою та маленькими зеленими грудьми; Дослідник: черговий збочений студент; Станіус – гарячий латиш брат маркіза де Сада; Рінгер – енуретичний коханець барона Захер-Мазоха. Тишина, пара. В кабінет вривається Дослідник, дико розмахуючи скальпелем та Жабою, волає: “О зрадницька потворо, о як могла розбити серце ти моє, о де взялась та злая банабацька сила, що заставляє битись серденько твоє, а може глянуть!?”. Бідкаючись, він вправно вводить ножиці між щелепами Жабі. Жаба: “Ква-а-а-а-о-о-о-так”. Дослідник: “Ну-ну”. Жаба: “Ще!!!”. Дослідник: “Ну ось і все”, – децеребрує та розтинає їй пузо. Тиша, в кабінет вбігає Рінгер та поливає своїм розчином жабяче нутро. Серце Жаби знову б’ється, вона хтиво всміхається. Дослідник накладає першу лігатуру між венозною пазухою та передсердям: “Здохни”. Знову влітає Рінгер та з криком “Замочу” робить свою мокру справу. Серце відроджується, але ритм вже не той, сили покидають принцесу... Після накладання третьої лігатури на нижню третину шлуночка у Рінгера настає зневоднення, Дослідник сходить з глузду та робить собі харакірі, перед очима Жаби під спів янголів проносяться найщасливіші моменти життя. Двері ногою вибиває Станіус з цигаркою в роті та кричить “ПЕ-РЕ-РВА-А”. Завіса. 3. Потенціал дії атипових кардіоміоцитів сино-атріального вузла, механізми походження, фізіологічна роль. Природа автоматії полягає в наявності в А-КМЦ специфічних потенціал-чутливих каналів. Ці канали змінюють свій стан, коли в ході реполяризації мембрани КМЦ (кінець попереднього ПД) мембранний потенціал досягає 60 mV. При цьому в мембрані А-КМЦ: - відкриваються повільні кальцієві канали – йони кальцію за градієнтом концентрації починають повільно входити в клітини; - відкривається повільні натрієві канали – йони натрію за градієнтом концентрації починають повільно входити в клітини; - закриваються калієві канали – зменшується вихід калію з клітини за градієнтом концентрації. Така зміна стану каналів мембран А-КМЦ веде до повільного зменшення мембранного потенціалу (деполяризація мембрани). Ця деполяризація виникає без дії зовнішнього подразника (автоматично), і коли вона досягає критичного рівня (– 45 mV), виникає пік ПД. Ця частина змін в часі мембранного потенціалу клітини, що володіє автоматією, є специфічною для неї і носить назву фази повільної діастолічної деполяризації, або спонтанної деполяризації. 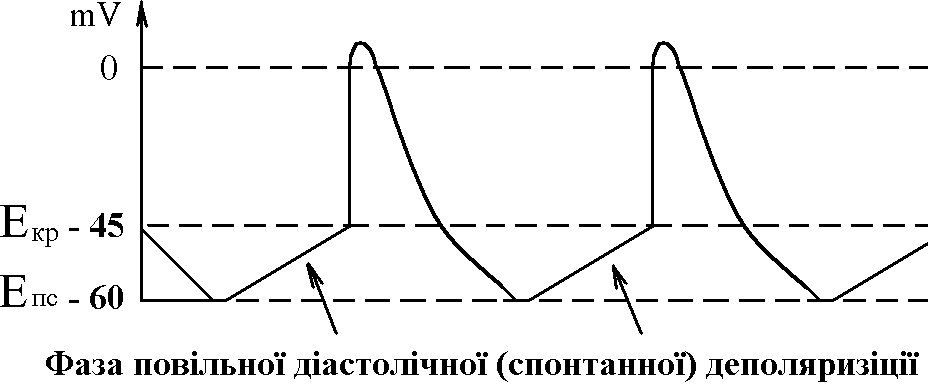 Частота, з якою центр автоматії генерує ПД, залежить від двох факторів: 1) величина порогового потенціалу; чим вона більша, тим частота менша; в звичайних умовах під впливом механізмів регуляції частіше змінюється рівень мембранного потенціалу спокою зміна порогового потенціалу зміна частоти генерації імпульсів збудження водієм ритму зміна частоти серцевих скорочень; 2) швидкість повільної діастолічної деполяризації (ПДД); механізми регуляції змінюють проникність відповідних каналів зміна швидкості ПДД зміна частоти генерації ПД водієм ритма серця зміна ЧСС. Центри автоматії другого та наступних порядків генерують імпульси збудження рідше, ніж водій ритма (пазухово-передсердний вузол), перш за все тому, що у них менша швидкість ПДД. Дуже рідко (в умовах патології) здатність до автоматії проявляється в Т-КМЦ (з яких побудований робочий міокард шлуночків та передсердь). Це відбувається при різкому зменшенні рівня мембранного потенціалу спокою цих клітин (до –60 mV, як в А-КМЦ; в звичайних умовах рівень їх потенціалу спокою дорівнює –90 mV). Такі центри автоматії носять назву ектопічних. 4. Провідна система серця. Послідовність і швидкість проведення збудження по серцю. В 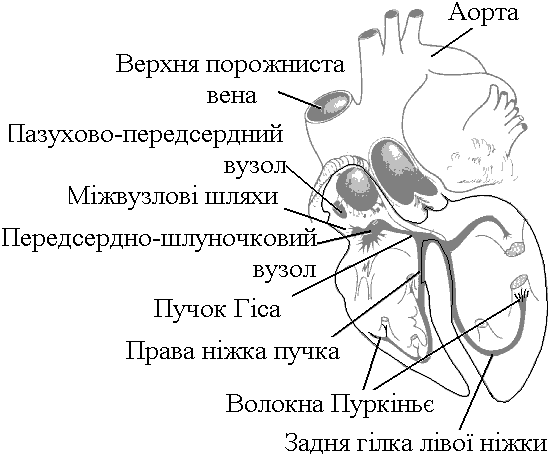 звичайних умовах послідовність руху збудження по структурах серця така: пазухово-передсердний вузол робочий міокард передсердь передсердно-шлуночковий вузол пучок Гіса ніжки пучка Гіса звичайних умовах послідовність руху збудження по структурах серця така: пазухово-передсердний вузол робочий міокард передсердь передсердно-шлуночковий вузол пучок Гіса ніжки пучка Гіса волокна Пуркіньє робочий міокард шлуночків. Швидкість проведення збудження по структурах серця різна. Вона складає: - робочий міокард передсердь та шлуночків – біля 1 м/с; - пучок Гіса, його ніжки, волокна Пуркіньє – 2-5 м/с. Тобто, швидкість велика і це має велике значення, так як забезпечує синхронність скорочень міокарду передсердь; міокарду шлуночків (для досягнення останнього результату існує провідна ситема шлуночків серця – пучок Гіса, його ніжки, волокна Пуркіньє; наявність спеціальної провідної системи шлуночків зумовлено його великою масою). Чинниками, що впливають на швидкість проведення збудження по м’язовим волокнам є: діаметр волокон, амплітуда ПД, величина порогу деполяризації, швидкість розвитку піку ПД, наявність нексусів між міокардіоцитами – вони мають низький опір, що сприяє швидкій передачі ПД з одного КМЦ на другий і збільшенню швидкості проведення збудження. Причинами великої швидкості проведення збудження по провідній системі серця є: - великий діаметр волокон; - наявність нексусів; - достатня амплітуда ПД (близько 100 мВ). Дещо менший діаметр Т-КМЦ та гірше розвинені нексуси є причиною меншої швидкості проведення збудження по ним. В ділянці передсердно-шлуночкового вузла має місце затримка проведення збудження, котра виникає внаслідок значного зменшення швидкості проведення (2-5 м/с). Наявність атріовентрикулярної затримки забезпечує послідовне скорочення передсердь та шлуночків (спочатку передсердя, а через 0,1 с – шлуночки). Причини малої швидкості проведення збудження по волокнам передсердно-шлуночкового вузла є: - малий діаметр волокон; - відсутність нексусів; - невелика амплітуда ПД (60-70 мВ); - мала швидкість розвитку ПД (в цих клітинах розвиток ПД має не натрієву, а кальцієву природу, тому розвивається повільно). 5. Потенціал дії типових кардіоміоцитів шлуночків, механізми походження, фізіологічна роль. Співвідношення у часі ПД одиночного скороченння міокарда. Типові кардіоміоцити (Т-КМЦ) не мають властивості автоматії і генерують ПД під впливом подразника (ПД, що йде від водія ритму серця). ПД в Т-КМЦ має особливості, а саме, він дуже тривалий – в шлуночках до 300 мс (в нервових волокнах – 1 мс, в скелетних м'язах – 2-5 мс). Фази ПД Т-КМЦ: 1 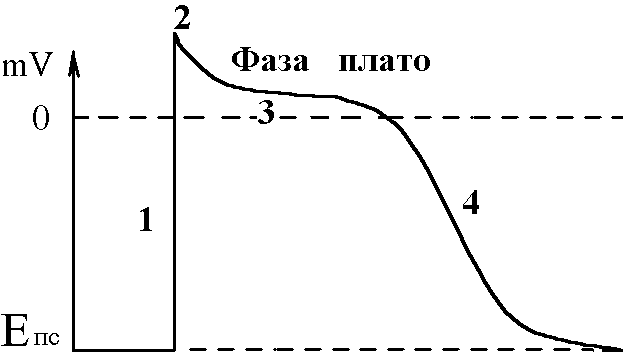 . Фаза швидкої деполяризації; початкова фаза її пов’язана із швидким входом йонів натрію, потім додається вхід йонів кальцію. . Фаза швидкої деполяризації; початкова фаза її пов’язана із швидким входом йонів натрію, потім додається вхід йонів кальцію.2. Фаза швидкої початкової реполяризації – дуже короткочасна. Пов’язана з виходом із Т-КМЦ йонів калію та вхід хлору 3. Фаза повільної реполяризації (плато) під час цієї фази мембранний потенціал Т-КМЦ мало змінюється, оскільки вихід йонів калію зрівноважується входом йонів кальцію. 4. Фаза швидкої реполяризації – пов’язана із швидким виходом із клітин калію – відновлення вихідного рівня мембранного потенціалу. Таким чином, велика тривалість ПД пов’язана з наявністю фази плато. Вона в свою чергу виникає внаслідок наявності в Т-КМЦ специфічних потенціал-чутливих кальцій-натрієвих каналів. Ці канали відкриваються під час швидкої деполяризації, коли мембранний потенціал зменшується до рівня (30-40 мВ). Ці канали повільно відкриваються, зате довго лишаються відкритими. Через них довго здійснюється вхід в Т-КМЦ йонів кальцію (значно менше – натрію) за градієнтом концентрації. 6. Періоди рефрактерності під час розвитку ПД типових кардіоміоцитів, їх значення. Значення великої тривалості ПД Т-КМЦ стає зрозумілим, якщо співставити його в часі з графіком зміни збудливості Т-КМЦ при збудженні з графіком поодинокого скорочення міокарда: 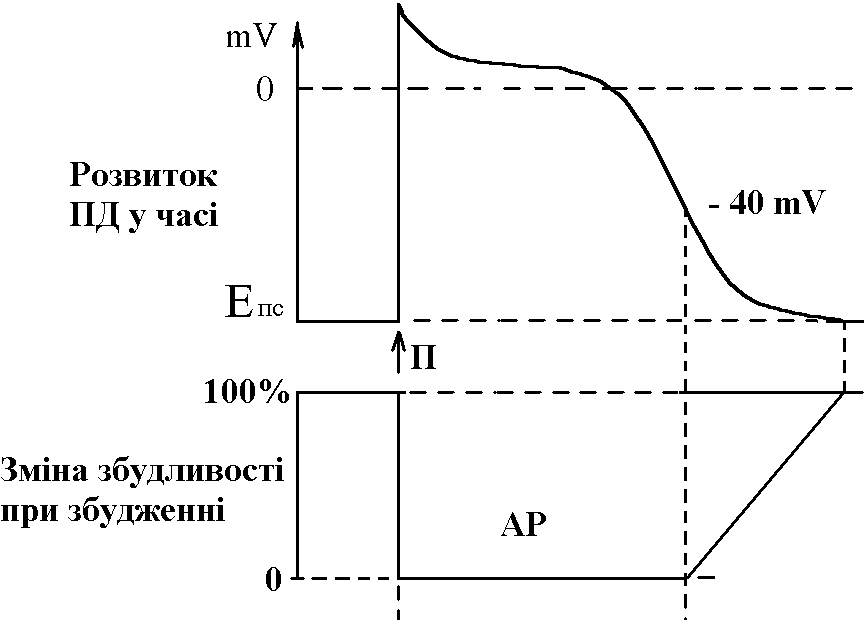 ПД Т-КМЦ тривалий через наявність фази плато. Довготривалий ПД є причиною тривалої абсолютної рефрактерної фази (АР) – час протягом якого Т-КМЦ повністю незбудливі. 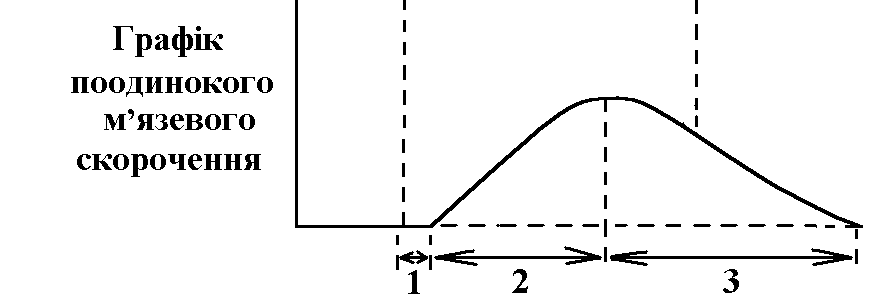 АР відповідає розвитку латентного періоду поодинокого м’язевого скорочення, періоду укорочення та значної частини періоду розслаблення. 1. Латентний період. 2. Період укорочення. 3. Період розслаблення. Завдяки такому співвідношенню у часі фаз збудливості та періодів поодинокого скорочення міокарда досягається: - неможливість виникнення в міокарді тетанічних скорочень; наступний цикл збудження (і скорочення) стає можливим тільки в фазі відносної рефрактерності, коли міокард закінчив своє скорочення і в значній мірі розслабився. Це дуже важливо, так як для виконання насосної функції серцем необхідно, щоб воно наповнилось кров’ю під час розслаблення. Тетанічне скорочення унеможливлювало б нагнітальну функцію серця; - неможливість патологічної рециркуляції збудження по структурах серця (тривала фаза абсолютної рефрактерності не дає можливості збудженню повернутись туди, де воно було деякий час тому назад). 7. Спряження збудження і скорочення в міокарді. Механізми скорочення і розслаблення міокарду. Спряження (зв’язок) збудження і скорочення в міокарді принципово проходить так само, як і в скелетних м’язах. Тобто, ПД викликає скорочення таким чином: ПД поширюється по мембрані Т-КМЦ, в тому числі і по мембрані Т-трубочок відкриття кальцієвих каналів саркоплазматичного ретикулума (СПР) вихід йонів кальцію із СПР підвищення концентрації йонів кальцію в міоплазмі з 10-8 до 10-5 моль/л дифузія йонів кальцію до скоротливих білків (протофібрил) взаємодія з регуляторними білками (з тропоніном) зміна третинної структури тропоніну та тропоміозину відкриття активних центрів актину взаємодія активних головок міозину з активними центрами актину скорочення міокарду. Необхідно підкреслити, що сила серцевих скорочень (ССС) залежить від кількості актоміозинових містків, які утворюються при скороченні. Особливостями процесу спряження збудження та скорочення в міокарді є: - необхідність для виходу йонів кальцію із СПР (кальцієвого залпу) входу йонів кальцію із міжклітинної рідини. Цей вхід проходить під час фази плато ПД; - наявність кількісного взаємозв’язку між входом кальцію в клітину під час фази плато ПД і його виходу із СПР, а отже, і ССС (регуляторні механізми, наприклад, можуть підвищувати кількість повільних кальцієвих каналів, через які проходить вхід йонів кальцію під час фази плато ПД підвищення входу кальцію під час фази плато ПД посилення виходу йонів Сa+ із СПР підвищення кількості відкритих активних центрів актину підвищення кількості акто-міозинових містків посилення ССС). Стан кальцієвих каналів можуть змінювати лікарі, призначаючи хворим їх блокатори (варапаміл) зменшення входу йонів кальцію в Т-КМЦ при їх збудженні зменшення ССС. Механізми розслаблення міокарду полягає у видаленні із міоплазми йонів кальцію, які надійшли туди під час “кальцієвого залпу”. Цей результат досягається завдяки: - активації кальцієвих насосів повздовжніх трубочок СПР (із затратами енергії АТФ); - активації кальцієвих насосів зовнішньої мембрани Т-КМЦ (із затратами енергії АТФ); - роботі натрій-кальцієвого йонообмінного механізму; цей механізм забезпечує транспорт в протилежному напрямку через зовнішню клітинну мембрану йонів Na+ (в клітину) і йонів Сa+ (з клітини). Йони Na+ входять в клітину за градієнтом концентрації, який створюється натрій-калієвим насосом (працює з затратами енергії АТФ). Спряжений з входом йонів Na+ вихід йонів Сa+ знижує його концентрацію в клітині та сприяє розслабленню міокарда. 8. Векторна теорія формування ЕКГ. ЕКГ, відведення. Походження зубців, сегментів, інтервалів ЕКГ. При збудженні та реполяризації серця виникає електричне поле, яке можна зареєструвати на поверхні тіла. При цьому між різними точками тіла створюється різниця потенціалів, яка змінюється у відповідності з коливаннями величини та напрямку цього електричного поля. Крива змін цієї різниці потенціалів в часі називається електрокардіограмою (ЕКГ). Таким чином, ЕКГ відображає збудження серця, а не його скорочення. Для розуміння генезу ЕКГ необхідно знати наступні факти: 1. Загальне електричне поле серця утворюється в результаті сумації полів чисельних окремих волокон серця; 2. Кожне збуджене волокно є диполем, що містить в собі елементарний дипольний вектор певної величини та напрямку; 3. Інтегральний вектор в кожен момент процесу збудження є результуючою окремих векторів; 4. Величина потенціалу, що вимірюється в точці, яка віддалена від джерела, залежить головним чином від величини інтегрального вектора і від кута між напрямком цього вектора та віссю відведення. |