Генетика. 34. Ген как структурная и функциональная единица наследственности. Классификация и свойства генов. Регуляторные гены и регуляторные последовательности
 Скачать 3.46 Mb. Скачать 3.46 Mb.
|

|
34. Ген как структурная и функциональная единица наследственности. Классификация и свойства генов. Регуляторные гены и регуляторные последовательности. Элементарной функциональной единицей наследственности является ген, его химическая основа — молекула ДНК. Критерием функции гена является его экспрессия (работа) в процессе реализации генетической информации: ДНК —> РНК —> белок —> признак. По своему функциональному значению гены классифицируются на структурные, регуляторные и гены-модуляторы. Структурные гены делятся на две группы: гены I, кодирующие структуру белков (полипептидов), гены II — структуру рРНК, тРНК. Регуляторные гены координируют активность структурных генов на уровне клетки и на уровне организма в процессе онтогенеза, а также детерминируют синтез регуляторных белков. Наряду с регуляторными генами, есть регуляторные последовательности (промотор, оператор, терминатор, энхансер, элемент перед промотором), функции которых выявляются при взаимодействии со специфическими регуляторными белками. Структурные гены делят на гены «домашнего хозяйства», продукты экспрессии которых необходимы постоянно для жизнедеятельности любого типа клеток (гены рРНК, тРНК, гены гистонов, гены тубулинов и др.), и гены «роскоши» — тканеспецифические гены, обеспечивающие специализированные функции клеток, т. е. гены функционально активные только в определенных типах клеток и на определенных стадиях развития организма (гены глобина, инсулина, кристаллина, иммуноглобулинов (локус HLA) и др. 35. Организация и строение генов у эукариот. Понятие о мозаичном гене, особенности его строения. Понятие о кластере. У эукариот основным типом организации генов являются кластеры. Кластеры генов — это группа родственных генов со сходными функциями, локализованная в определенных участках хромосом. В состав кластера входят активно функционирующие гены и псевдогены. Часто кластеры являются семейством генов, произошедшим от некоего гена-предка. Классическим примером являются гены глобинов в составе А- и В-кластеров. В конце 70-х годов было выяснено, что у эукариот имеются гены, которые содержат «лишнюю» ДНК, не представленную в молекуле мРНК. Они получили название мозаичных, прерывистых генов; генов, имеющих экзон-интронное строение. 1.Мозаичные гены эукариот имеют больший размер, чем последовательность нуклеотидов, представленная в мРНК (3-5%). 2.Мозаичные гены состоят из экзонов и интронов. Интроны удаляются из первичного транскрипта и отсутствуют в зрелой мРНК, которая состоит только из экзонов. Число и размеры интронов и экзонов индивидуальны для каждого гена, но интроны по размерам значительно больше экзонов. 3.Ген начинается экзоном и заканчивается экзоном, но внутри гена может быть любой набор интронов (гены глобина имеют 3 экзона и 2 интрона). Экзоны и интроны обозначаются цифрами или буквами в порядке их расположения вдоль гена. 4. Порядок расположения экзонов в гене совпадает с их расположением в мРНК. 5.На границе экзон-интрон имеется определённая постоянная последовательность нуклеотидов (ГТ - АГ), присутствующая во всех мозаичных генах. 6. Экзон одного гена может быть интроном другого. 7. В мозаичном гене иногда нет однозначного соответствия между геном и кодируемым им белком, то есть одна и та же последовательность ДНК может принимать участие в синтезе различных вариантов белка. 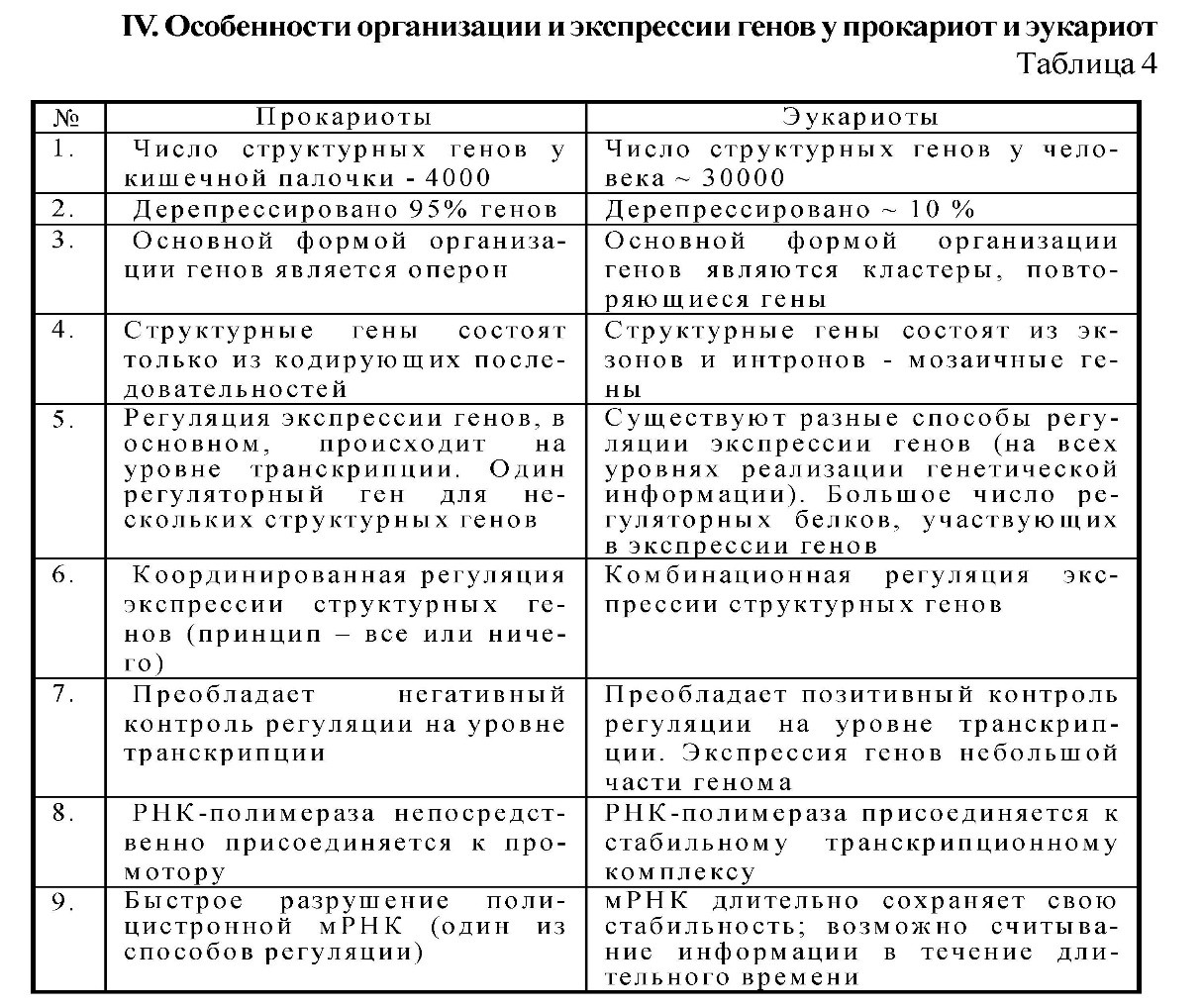 36. Организация генов и регуляция активности генов у прокариот. Строение оперона, экспрессия структурных генов оперона. См. вопр. № 35. У прокариот основным типом организации генов являются опероны (например, лактозный оперон кишечной палочки Е. coli). Оперон Е. coli — это группа структурных генов А, В, С, расположенных друг за другом, которые имеют общий промотор, оператор (нуклеотидные последовательности промотора и оператора перекрываются) и терминатор. Они участвуют в одном метаболическом цикле (в данном случае расщепление лактозы до глюкозы и галактозы) и регулируются координированно. Структурные гены в составе оперона находятся под контролем оператора. Регуляция оперона осуществляется геном-регулятором. Возможны два состояния лактозного оперона: 1 — оперон «выключен», 2 — оперон «включен». 1. Связывание белка-репрессора с оператором прекращает процесс транскрипции: РНК-полимераза не может присоединиться к промотору, экспрессии структурных генов не происходит, ферменты не синтезируются. 2. Поступающий в клетку индуктор (лактоза) соединяется с белком-репрессором, отбирая его от оператора, что обеспечивает присоединение РНК-полимеразы к промотору и экспрессию генов. Результатом этого является синтез белков-ферментов, которые расщепляют лактозу до глюкозы и галактозы. Лактозный оперон, как единица транскрипции, регулируется по типу негативной регуляции: гены экспрессируются при условии, что они не выключены регуляторным белком (белком-репрессором). 37. Регуляция экспрессии генов у эукариот. Уровни регуляции. У эукариот все клетки многоклеточного организма содержат одинаковую ДНК, но в то же время они отличаются друг от друга по морфологическим, физиологическим и биохимическим свойствам. В основе этих фенотипических различий лежит экспрессия разных генов и соответственно синтез разных мРНК и белков. Большая часть белков синтезируется всеми клетками. Для этого необходима экспрессия генов «домашнего хозяйства», они постоянно экспрессированы. Некоторые белки синтезируются только в определенных клетках (глобины и др.), синтез таких белков, а соответственно и экспрессия генов регулируется: гены «включаются» и «выключаются». Экспрессия генов регулируется на различных уровнях реализации наследственной информации: претранскрипционном, транскрипционном, посттранскрипционном, претрансляционном, трансляционном и посттрансляционном. 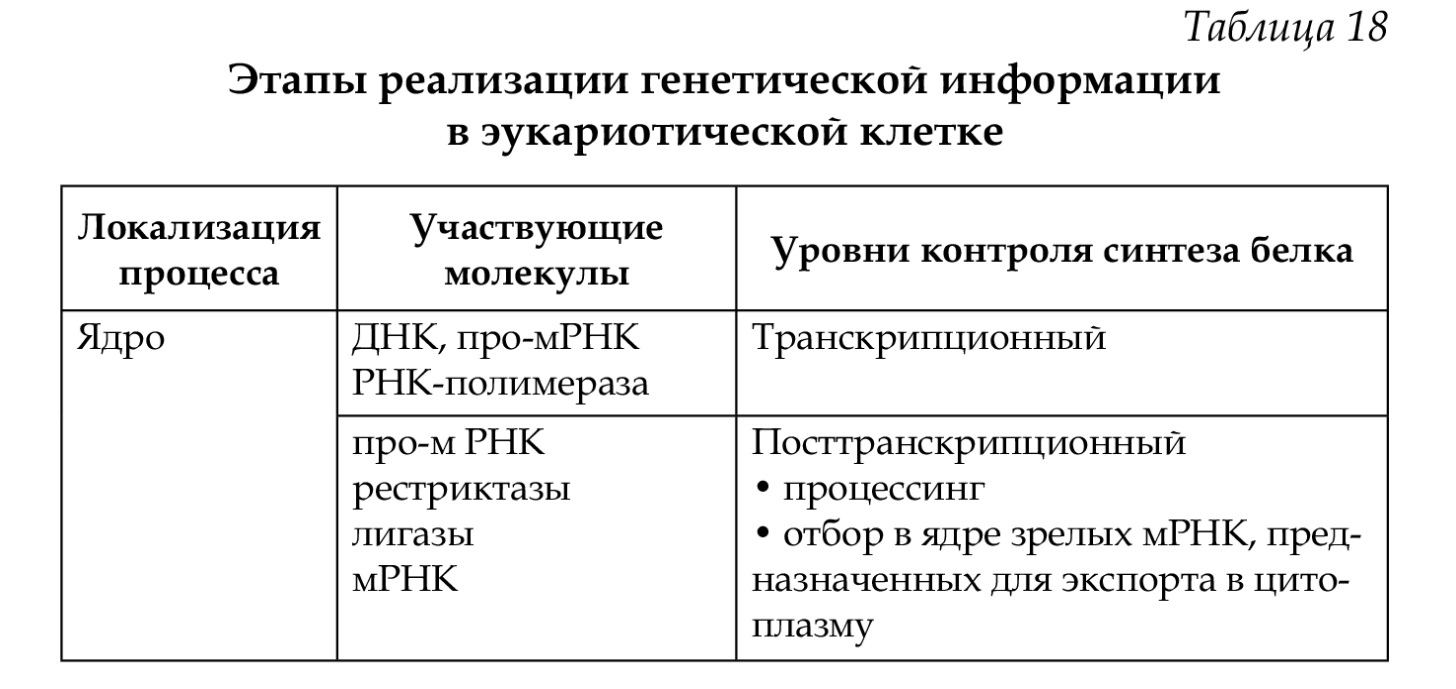 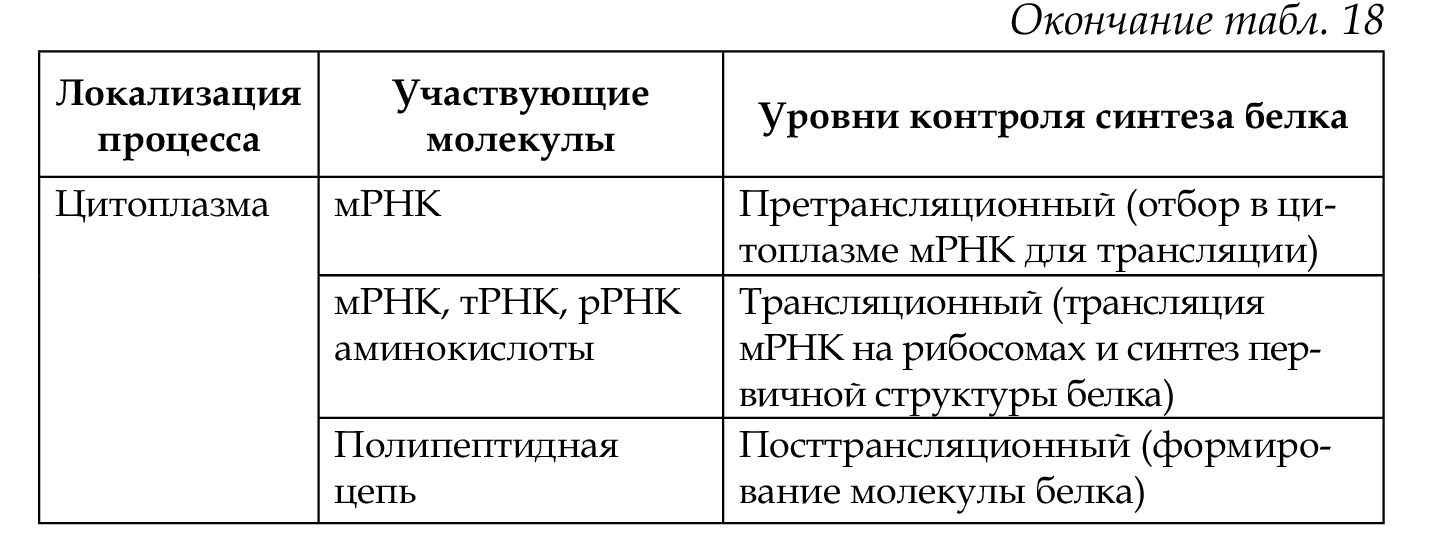 38. Глобиновые гены, их локализация, строение и избирательная экспрессия в процссе онтогенеза. Гемоглобин состоит из гема и белка тетрамера-глобина. Глобиновый тетрамер состоит из двух идентичных P-цепей и двух идентичных a-цепей. Аминокислотная последовательность каждой глобиновой цепи кодируется своими собственными генами, входящими соответственно в состав А- и В-кластера. У человека A-кластер располагается в 16 хромосоме, а В-кластер — в 11 хромосоме. На разных стадиях эмбриогенеза происходит избирательное и последовательное включение и выключение генов А- и В-кластеров. Гены глобина располагаются в А- и В-кластерах в определенной последовательности и транскрибируются слева направо в порядке их экспрессии. У человека образование «взрослого» гемоглобина включает несколько этапов: эмбриональный гемоглобин, гемоглобин плода и гемоглобин взрослого человека (после рождения). Гены глобина по внутреннему строению являются мозаичными. 39. Гибридологический метод – основной метод генетического анализа. Законы Г. Менделя. Гибридологический метод разработал Г. Мендель. Гибридологический метод — это анализ характера наследования признаков с помощью системы скрещиваний, суть которых состоит в получении гибридов и анализе их потомков в ряду поколений. Особенности гибридологического метода: 1. Подбор исходных родительских пар (гомозиготы с четкими альтернативными признаками). 2. Получение гибридов и последующее их скрещивание между собой. 3. Использование количественного учета потомков, различающихся по отдельным признакам в ряду последовательных поколений (результаты скрещивания анализируются статистически-математическим анализом). Моногибридным называется скрещивание, при котором прослеживается наследование одного признака. Первый закон Менделя — закон единообразия гибридов первого поколения: при скрещивании гомозиготных особей, отличающихся альтернативным проявлением одного признака, все потомство будет единообразным по фенотипу и генотипу. Полученные особи называются гибридами. Второй закон Менделя — закон расщепления: при скрещивании гибридов первого поколения между собой в потомстве происходит расщепление признака по фенотипу 3 :1, а по генотипу 1:2:1. Дигибридное скрещивание — это скрещивание, при котором анализируется наследование двух признаков одновременно. Третий закон Менделя — закон независимого наследования и независимого комбинирования признаков: при ди- и полигибридных скрещиваниях каждый признак наследуется независимо от другого, расщепляясь в соотношении 3:1. При дигибридном скрещивании в F2 формируются четыре фенотипа в соотношении 9:3 :3 :1, при этом два из них — рекомбинантные. Закон соблюдается, если неаллельные гены находятся в разных парах гомологичных хромосом и отсутствует взаимодействие между ними. 40. Условия менделирования признаков. Примеры нарушений закономерностей наследования. Законы Менделя выполняются при определенных условиях. Условия менделирования признаков: • моногенное наследование; • полное доминирование; • равновероятное образование всех гамет; • равновероятная встреча всех гамет при оплодотворении; • равновероятная выживаемость всех зигот; • отсутствие летальных мутаций; • отсутствие взаимодействий между неаллельными генами (при полигибридном скрещивании); • полная пенетрантность гена; • выраженная стойкая экспрессивность гена. 41. Взаимодействие аллельных генов. Примеры. Формулы скрещивания. Взаимодействие аллельных генов — это взаимодействие между аллелями одного и того же гена. Взаимодействие осуществляется по типу: полного доминирования, неполного доминирования, сверхдоминирования, кодоминирования. Полное доминирование — доминантный аллель полностью подавляет рецессивный; проявление доминантного аллеля не зависит от присутствия в генотипе другого аллеля; гетерозиготы и гомозиготы фенотипически не отличаются (желтый цвет горошин доминирует над зеленым, карие глаза — над голубыми). Неполное доминирование — доминантный аллель неполностью подавляет рецессивный, а у гетерозигот формируется промежуточный фенотип (розовые цветки ночной красавицы). Кодоминирование — два доминантных аллеля одного гена проявляются в фенотипе независимо друг от друга (организм с генотипом IA IB определяет синтез двух видов антигенов А и В). Сверхдоминирование — у гетерозигот признак выражен сильнее, чем у гомозигот; доминантный аллель в гетерозиготном состоянии имеет более сильное проявление, чем в гомозиготном (гетерозисная сила). 42. Взаимодействие неаллельных генов. Примеры. Формулы скрещивания. Различают три основных типа взаимодействия неаллельных генов: комплементарность, эпистаз (качественные признаки), полимерия (количественные признаки). Комплементарность — тип взаимодействия, при котором два неаллельных гена, обычно доминантных, находясь одновременно в генотипе, приводят к формированию нового проявления признака. У человека для образования некоторых признаков необходимо комплементарное взаимодополняющее действие генов (формирование нормального слуха, синтез гемоглобина). Эпистаз — тип взаимодействия, при котором наблюдается подавление действия одного гена другим, неаллельным ему. Подавляющий ген называется эпистатическим геном (геном супрессором или ингибитором), а подавляемый — гипостатическим. Различают эпистаз доминантный (доминантный аллель одного гена подавляет проявление доминантных аллелей другого гена) и рецессивный (рецессивный аллель подавляет проявление доминантных аллелей другого гена только в гомозиготном состоянии). Полимерия — тип взаимодействия неаллельных генов, когда при формировании признака важно количество доминантных генов, влияющих на степень его выраженности. Гены обозначаются одинаковыми буквами (цифровое обозначение с нижним индексом), чтобы подчеркнуть однонаправленность их действия А1, А2, ..., А10. У человека по типу полимерии наследуются пигментация кожи (чем больше доминантных аллелей, тем больше образуется меланина и тем интенсивнее окраска кожи), а также большинство количественных признаков, которые можно измерить или подсчитать (вес, рост, артериальное давление). 43. Явление множественного аллелизма, его закономерности, значение. Наследование групп крови системы АВО. Развитие признака определяется двумя аллелями одного гена (А и а), которые занимают идентичные локусы гомологичных хромосом. Иногда ген имеет не два, а большее число аллелей, которые возникают в результате мутации. Многократное мутирование одного и того же гена образует серию множественных аллелей, а само явление называется явлением множественного аллелизма. Оно имеет широкое распространение: окраска шерсти у кроликов, цвет глаз у дрозофилы, система групп крови АВО у человека. Имеются определенные закономерности множественного аллелизма: — каждый ген может иметь большое число аллелей; — любой аллель может возникнуть в результате прямой и обратной мутации любого члена серии множественных аллелей или от аллеля дикого типа; — в диплоидном организме могут одновременно находиться два любых аллеля из серии множественных аллелей; — аллели находятся в сложных доминантно-рецессивных отношениях между собой: один и тот же аллель может быть доминантным по отношению к одному аллелю и рецессивным по отношению к другому, а между иными аллелями доминирование может отсутствовать, и наблюдается кодоминирование и др.; — члены серии множественных аллелей наследуются так же, как и пара аллелей, т. е. наследование подчиняется менделевским закономерностям (кроме кодоминирования); — разные сочетания аллелей в генотипе обуславливают различные фенотипические проявления одного и того же признака; — серии аллелей увеличивают комбинатов ну ю изменчивость. Примером множественного аллелизма у человека является наличие трех аллелей гена, определяющего наследование групп крови системы АВО. • система определяется тремя аллелями одного гена I (IA, Iв, I°); ген I расположен в 9-й хромосоме: 9q34; • из всей серии аллелей одновременно в генотипе диплоидного организма находятся два аллеля (I°I0, IAIA, IAI°, 1в1в и др.); • аллели IA, 1в доминантны по отношению к аллелю 1° — полное доминирование, между собой аллели 1А и 1в — кодоминантны; • доминантный аллель гена может проявлять свое действие в гомо- (IAIA, IBIB) и гетерозиготном организмах (1А1°, 1в 1°), а рецессивный аллель гена — только в гомозиготном организме (1° 1°); • различные сочетания аллелей в генотипе дают разные фенотипы: 4 группы крови I (0), II (А), III (В), IV (АВ), которые различаются между собой антигенными свойствами эритроцитов. Антигены (агглютиногены) находятся на поверхности эритроцитов (гликокаликс); • особенностью системы является наличие в сыворотке крови спецефических антител (агглютининов), разноименных по отношению к собственным агглютиногенам (они одновременно находятся в крови); • разнообразие групп крови обеспечивает фенотипический полиморфизм в популяциях человека по данному признаку. Группы крови являются примером однозначной нормы реакции организма (группа крови не изменяется в течение жизни ни при каких изменениях среды). 44. Изменчивость и ее формы. Значение изменчивости в онто- и филогенезе. Изменчивость — свойство организмов изменять признаки, полученные от родителей, или приобретать новые в процессе индивидуального развития. Различают модификационную, генотипическую (комбинативную и мутационную) и онтогенетическую изменчивость. 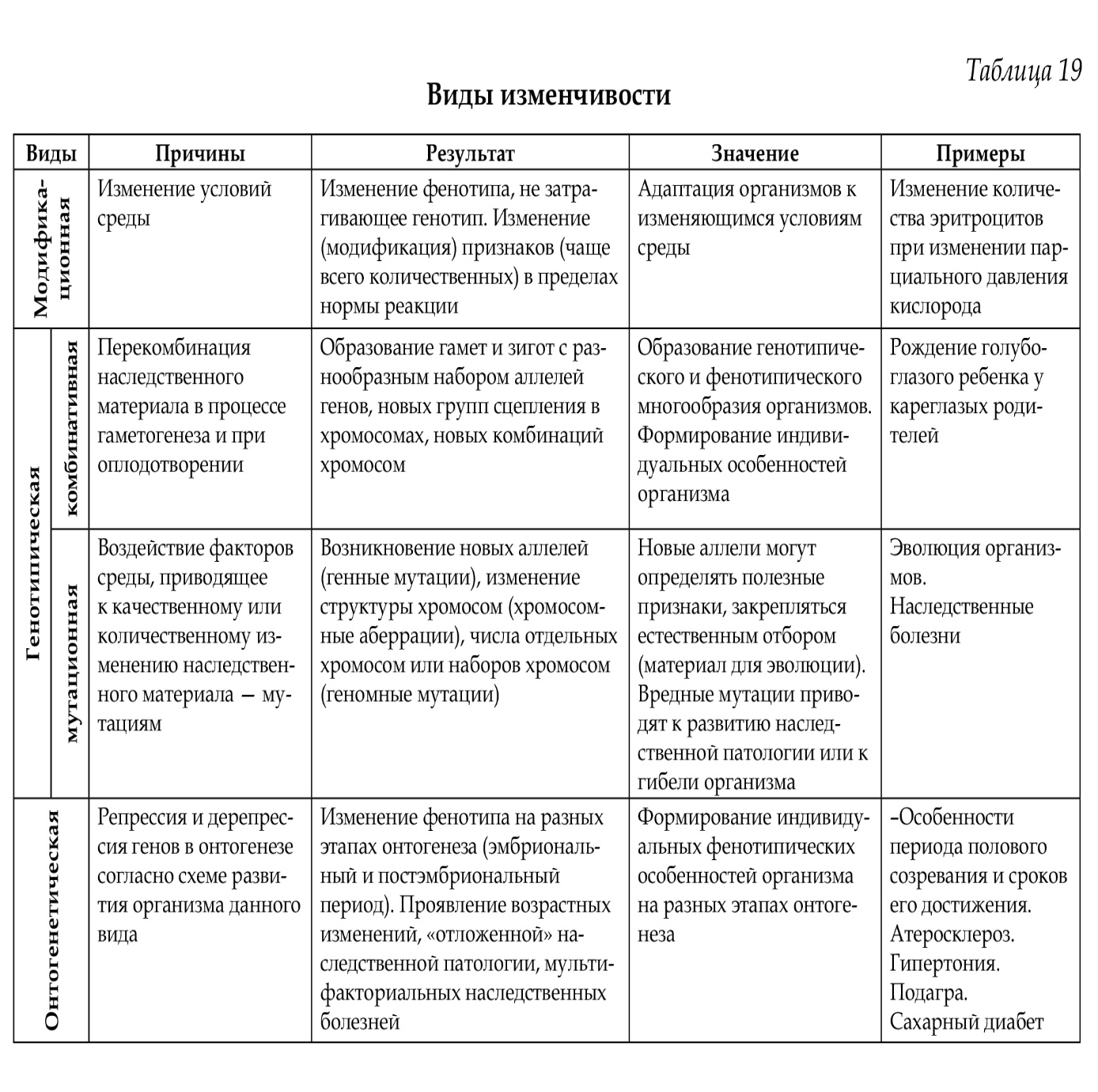 45. Модификационная изменчивость. Норма реакции. Онтогенетическая изменчивость. Реализация генотипа в фенотип. См. вопр. № 44. Норма реакции — способность генотипа формировать в онтогенезе, в зависимости от условий среды, разные фенотипы. Она характеризует долю участия среды в реализации признака и определяет модификационную изменчивость вида. Чем шире норма реакции, тем больше влияние среды и тем меньше влияние генотипа в онтогенезе. Процесс реализации генетической информации в фенотип организма в онтогенезе начинается с находящегося под прямым генетическим контролем образования простых белков (полипептиды, протеины). Функциональные свойства последних определяются аминокислотной последовательностью, которая задается последовательностью триплетов нуклеотидов в ДНК соответствующего структурного (смыслового, кодирующего, транскрибируемого и транслируемого, экспрессируемо-го) гена. Таким образом, полипептид, будучи первичным продуктом генетической активности, свойства которого напрямую определяются геном, является элементарным (простым) фенотипическим признаком. В соответствии с приведенным выше определением, элементарные фенотипические признаки организма - это в основном простые белки с каталитической, транспортной, рецепторной и другими функциями. 46. Генотипическая изменчивость, её виды (комбинативная, мутационная). Механизмы возникновения, значение. См. вопр. № 44. 47. Классификация мутаций. Мутагенез, его виды, мутагенные факторы. Мутации — это внезапные, случайные, ненаправленные, устойчивые изменения наследственного материала; одни и те же мутации могут возникать повторно. Существуют различные классификации мутаций: • по уровню организации живого (молекулярный, клеточный, организменный, популяционный); • по уровню организации наследственного материала (генные, хромосомные, геномные); • по преимущественному проявлению в фенотипе (морфологические, физиологические, биохимические); • по способу возникновения (спонтанные, индуцированные); • по влиянию на жизнь организма (летальные, полулетальные); • по типам клеток (соматические, генеративные); • по локализации в клетке (ядерные и цитоплазматические). Мутагенез - процесс возникновения, формирования и реализации наследственных нарушений. Основой мутационного процесса являются мутации. Мутации происходят как в естественной среде обитания организмов, так и в условиях направленного воздействия мутагенами. В зависимости от этого различают спонтанный и индуцированный мутагенез. Спонтанный мутагенез - это самопроизвольный процесс возникновения мутаций под влиянием естественных факторов среды. Существует несколько гипотез относительно генеза спонтанных мутаций: естественная радиация, наличие генов-мутаторов, определенное соотношение мутагенов и антимутагенов и др.По современным данным мутации возникают при нарушении процесса репликации и репарации ДНК. Спонтанный мутационный процесс характеризуется определенной интенсивностью (частотой генных, хромосомных и геномных мутаций), непрерывностью, ненаправленностью, отсутствием специфичности; он является одной из биологических характеристик вида (стабильность генотипа) и протекает постоянно. Частота спонтанных мутаций подвергается генному контролю (ферменты репарации) и параллельно влиянию естественного отбора (появление новых мутаций уравновешивается их элиминацией). Познание закономерностей спонтанного мутагенеза, причин его возникновения необходимо для создания специальных методов слежения за мутациями, чтобы контролировать их количество у человека. Индуцированный мутагенез - возникновение мутаций под влиянием направленных специальных факторов внешней среды - мутагенов. Способностью индуцировать мутации обладают различные мутагены физической, химической и биологической природы, которые вызывают соответственно радиационный, химический и биологический мутагенез. Физические мутагены: ионизирующее излучение, ультрафиолет, температура и др. Ионизирующая радиация оказывает непосредственное действие на гены (разрыв водородных связей ДНК, изменение нуклеотидов), хромосомы (хромосомные аберрации) и геномы (изменение числа и наборов хромосом). Химические мутагены (лекарственные препараты, никотин, алкоголь, гербициды, пестициды, кислоты, соли и др.) вызывают генные, реже хромосомные мутации. Биологические мутагены (вирусы, живые вакцины и др.) вызывают генные мутации и хромосомные перестройки. |