7. Спинной мозг (гистоморфофункциональные характеристики). Серое вещество спинного мозга. Сегментарнорефлекторный аппарат спинного мозга (строение, функции, синдромы поражения на различных анатомических и морфофункциональных уровнях).
 Скачать 0.82 Mb. Скачать 0.82 Mb.
|

|
7. Спинной мозг (гисто-морфофункциональные характеристики). Серое вещество спинного мозга. Сегментарно-рефлекторный аппарат спинного мозга (строение, функции, синдромы поражения на различных анатомических и морфофункциональных уровнях). Реципрокная иннервация мышц. Механизм возвратного торможения. Спинной мозг - цилиндрический тяж, покрытый оболочками, свободно располагающийся в полости позвоночного канала. Вверху он переходит в medulla oblongata; внизу спинной мозг достигает области 1-го или верхнего края 2-го поясничного позвонка. Диаметр спинного мозга не везде одинаков, в двух местах обнаруживаются два веретенообразных утолщения: в шейном отделе – шейное утолщение – intumescentia cervicalis (от 4-го шейного до 2-го грудного позвонка); в самой нижней части грудного отдела – поясничное утолщение – intumescentia lumbalis – (от 12-го грудного до 2-го крестцового позвонка). Оба утолщения соответствуют областям замыкания рефлекторных дуг от верхних и нижних конечностей. Образование этих утолщений тесно связано с сегментарным принципом строения спинного мозга. В спинном мозге насчитывается в общей сложности 31−32 сегмента: 8 шейных (СI−СVIII), 12 грудных (ThI−ThXII), 5 поясничных (LI−LV), 5 крестцовых (SI−SV) и 1−2 копчиковых (СоI−СII). Поясничное утолщение переходит в короткий конусовидный отдел, в мозговой конус – conus medullaris s. terminalis, от которого отходит длинная тонкая конечная нить – filum terminale. Длина спинного мозга в среднем у мужчин достигает 45 см, у женщин – 41-42 см. Соответственно формирующимся в последствии периферическим нервам в спинном мозге различают pars cervicalis, из которой формируются шейные нервы, pars thoracalis– грудные, и pars lumbalis, из которой выходят поясничные и крестцовые нервы. Внешняя форма спинного мозга Средний диаметр спинного мозга равен 1 см, в местах, где отходят спинномозговые нервы для конечностей образуются утолщения: шейное (intumescentia cervicalis) − на уровне С5−Th2 сегментов и пояснично-крестцовое (intumescentia lumbosacralis) на уровне L2−S2 сегментов. Всего на протяжении спинного мозга отходят 124 корешка (по 62 задних и передних). Из них формируется 31 пара спинномозговых нервов. По средней линии передней, или вентральной, поверхности спинного мозга идет передняя срединная щель – fissura mediana anterior; по средине задней поверхности тянется поверхностная продольная борозда – sulcus medianus posterior. Этими двумя образованьями спинной мозг разделяется на две симметричные половины. По сторонам от sulcus medianus posterior, в каждой половине спинного мозга идет sulcus lateralis posterior, в которую вступают задняя корешковая нить. Кнаружи от fissura mediana anterior проходит sulcus lateralis anterior, которая не представляет истинной борозды. В верхней грудной и шейной частях между sulcus medianus posterior и sulcus lateralis posterior замкнута тонкая продольная бороздка – sulcus intermedius posterior. Выходящие из sulcus lateralis anterior передние корешковые нити образуют отдельные, разделенные промежутками пучочки, передние корешки – radices anteriores. Задние корешковые нити, расположенные в один ряд вдоль sulcus lateralis posterior, образуют, сходясь кнаружи, такие же пучочки, задние корешки – radices posteriores. Затем передний и задний корешковые пучки следуют вместе к определенному foramen intervertebrale. Здесь задний корешковый пучок образует небольшое утолщение – ganglion spinale. В дальнейшем оба корешка соединяются друг с другом, образуя смешанный спинномозговой нерв, который затем делится на переднюю и заднюю ветви. Отходящие от спинного мозга корешковые пучки направляются в сторону соответственного спинномозгового отверстия в теле позвонка (рис. 1). А так как скорость роста спинного мозга, отстает от скорости роста позвоночного 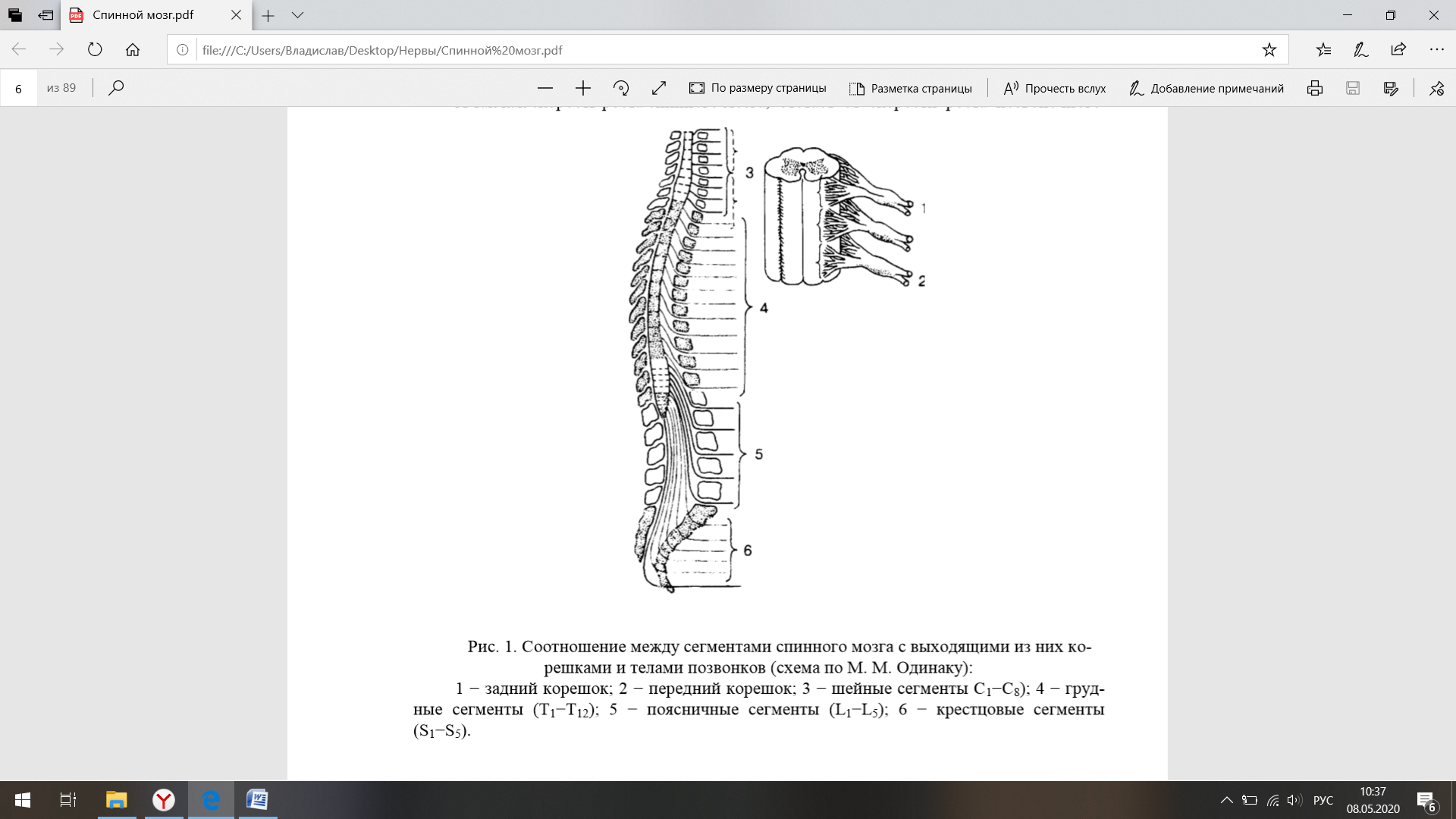 столба, то корешковые пучки направляются еще и вниз, и тем больше вниз, чем ближе к хвостовому концу спинного мозга. Направление нервных корешков в поясничной части спинного мозга внутри позвоночного канала становится почти параллельным продольной оси спинного мозга, так, что conus medullaris и filum terminale оказываются лежащими среди густого пучка нервных корешков, и вследствие сходства с конским хвостом все образование получило название cauda equina. Продольными бороздами разделяются следующие канатики спинного мозга: передний канатик – funiculus anterior – между fissura mediana anterior и sulcus lateralis anterior; боковой канатик – funiculus lateralis – между sulcus lateralis anterior и sulcus lateralis posterior; задний канатик – funiculus posterior – между sulcus medianus posterior и sulcus lateralis posterior, который разделяется посредством sulcus intermedius posterior на медиальный и латеральный канатики; медиальный канатик называется нежным пучком – fasciculus gracilis – , или пучок Голля, латеральный называется клиновидным пучком – fasciculus cuneatus –, или пучок Бурдаха. 1.1.2. Внутреннее расположение частей спинного мозга Спинной мозг состоит из симметричных половин. На поперечном срезе в спинном мозге можно легко различить серое и белое вещества (рис. 2). Серое вещество на поперечном срезе имеет очертание буквы "Н" и лежит в центре. Серое вещество состоит из нервных клеток с их отростками, нейроглии и сосудов. Часть нейронов, связанных между собой с помощью синаптических структур, располагается группами, что обычно обозначается как ядра. Основными являются: 1) двигательные, или моторные, клетки передних рогов, дающие волокна передних корешков (периферические двигательные нейроны); 2) чувствительные клетки − вторые нейроны болевой и температурной чувствительности, расположенные в заднем роге; 3) клетки проприоцепторов мозжечка − вторые нейроны, расположенные в основании заднего рога; 4) клетки вегетативных центров − преимущественно в боковых рогах; 5) ассоциативные клетки. В центре "Н"-образного серого вещества расположен центральный канал – canalis centralis – , окруженный студенистым веществом – substantia gelatinosa centralis, и выстланный эпендимой. Центральный канал расширяется вверху при переходе спинного мозга в продолговатый и переходит в четвертый желудочек. Книзу, в нижнем конце conus terminalis он расширяется в ventriculus terminalis (Krause), при переходе спинного мозга в filum terminale канал вновь суживается и оканчивается слепо. В раннем постнатальном периоде у доношенных детей спинномозговой канал запустевает. Кпереди от центрального канала расположена передняя серая спайка – commisura grisea anterior, перед которой лежит commisura alba anterior, которая доходит вентрально до fissura mediana anterior. Позади центрального канала расположена задняя серая спайка – commisura posterior (рис. 2). В каждой половине спинного мозга лежит передняя, более толстая часть серого вещества, передний рог – cornu anterius, и более тонкая задняя часть, задний рог – cornu posterius. Так как серое вещество тянется через весь спинной мозг непрерывно, то передние и задние рога кажутся колонками серого вещества – columnae griseae. 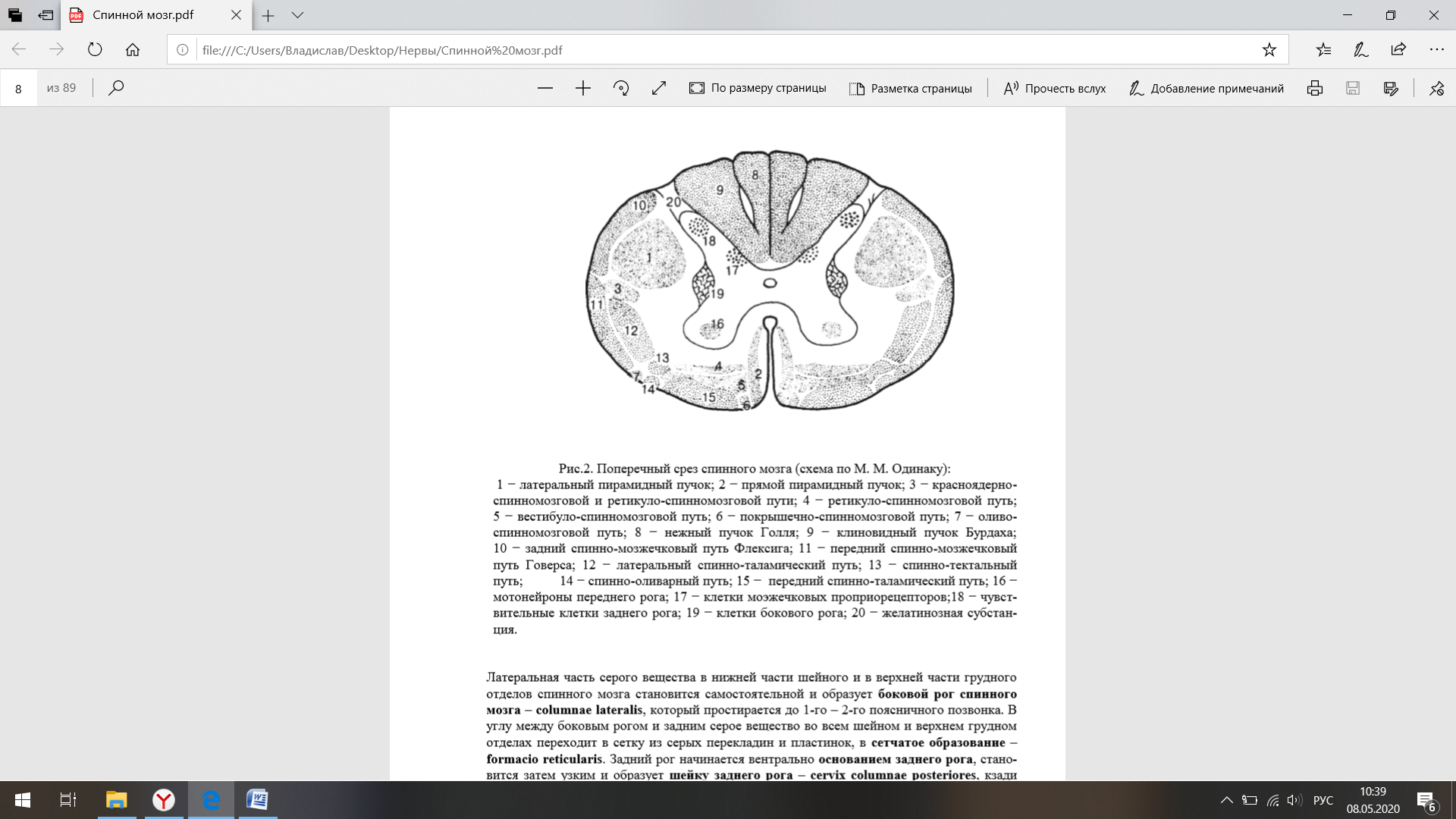 Латеральная часть серого вещества в нижней части шейного и в верхней части грудного отделов спинного мозга становится самостоятельной и образует боковой рог спинного мозга – columnae lateralis, который простирается до 1-го – 2-го поясничного позвонка. В углу между боковым рогом и задним серое вещество во всем шейном и верхнем грудном отделах переходит в сетку из серых перекладин и пластинок, в сетчатое образование – formacio reticularis. Задний рог начинается вентрально основанием заднего рога, становится затем узким и образует шейку заднего рога – cervix columnae posteriores, кзади переходящую в головку заднего рога – caput columnae и в верхушку заднего рога – apex columnae posteriores. Верхушка заднего рога состоит из полулунного поля – substantia gelatinosa (Rolandi) и лежит далее кзади от краевой зоны, или зонального слоя. Медиально от шейки заднего рога, по соседству с comissura posterior от нижнего отдела спинного мозга до верхнего поясничного отдела располагается внутри серого вещества маленькое ядро, nucleus dorsalis (Stillingi, Clarkii), столб Кларка. Состав клеток, находящихся в задних и передних рогах спинного мозга, неоднороден. В задних рогах располагаются чувствительные клетки, отростки которых переходят через среднюю линию спинного мозга в боковой столб противоположной стороны и составляют путь поверхностной чувствительности. В основании заднего рога выделяется обособленная группа клеток, относящихся к системе мозжечковой проприоцепции. Отростки этих клеток направляются в боковые столбы спинного мозга (передний перекрещивается на уровне собственного сегмента, задний − идет в боковой канатик своей стороны) и в составе спинноцеребеллярных путей доходят до ядра шатра червя мозжечка. В передних рогах спинного мозга различают три типа двигательных клеток: α-большие, α -малые и γ-нейроны. В боковых рогах спинного мозга располагаются сегментарные вегетативные эфферентные клетки. На уровне сегментов CVIII−LIII располагаются эфферентные клетки симпатической нервной системы. Кроме того, в спинном мозге находятся центры серого вещества. На уровне CVIII−THI, образован симпатический цилиоспинальный центр − centrum ciliospinale, симпатические волокна которого выходят с передними корешками, проходят через систему шейных симпатических ганглиев, симпатический нерв и ресничный узел; в глазу иннервируют три гладкие мышцы: m. dilatator pupillae − расширяющий зрачок; m. tarsalis superior − расширяющий глазную щель; m. orbitalis − обусловливающий своим напряжением соответствующую степень выстояния глазного яблока из глазницы. На уровне крестцовых сегментов SIII−SV находится спинальный парасимпатический центр регуляции функции тазовых органов (centrum vesicospinale et anospinale). Корковая иннервация их двусторонняя. Отсюда выходят волокна. направляющиеся к мочевому пузырю (его детрузору) и прямой кишке в составе тазового нерва (n. pelvicus) и снабжающие их гладкую, непроизвольную мускулатуру. От этих же сегментов начинается и срамной нерв (n. pudendus), иннервирующий поперечно-полосатые наружные, произвольные сфинктеры мочевого пузыря и прямой кишки. На этом же уровне берут начало симпатические волокна, обеспечивающие вазомоторные функции и влияющие на тонус гладких мышц тазовых органов, в частности, на тонус внутреннего непроизвольного сфинктера мочевого пузыря. На уровне уровне LI−LIII − симпатический центр регуляции органов малого таза: I−III поясничных сегментов находится центр эякуляции, а эрекции − на уровне II−V. Кроме того, в передних и задних рогах спинного мозга расположено большое количество вставочных нейронов, обеспечивающие замыкание рефлекторных дуг, связь между выше и ниже расположенными сегментами спинного мозга, связь между половинами спинного мозга, обеспечивающие десинхронизацию работы α-больших мотонейронов передних рогов спинного мозга и реципроктное торможение (клетки Реншоу).Между клетками серого вещества расположены клетки глии. Спинной мозг обладает сегментарным строением. Участок серого вещества спинного мозга с парой входящих и выходящих корешков называется сегментом спинного мозга. Сегмент спинного мозга входит в состав метамера тела, к которому определенный участок кожи (дерматом), группа мышц (миотом), кости (склеротом) и внутренние органы (спланхнотом), иннервируемые одним сегментом. Реципрокная иннервация мышц. Механизм возвратного торможения. Нервная регуляция работы скелетных мышц (регуляция запуска и выполнения всех движений), осуществляется двигательными центрами ЦНС. Они должны гарантировать строго необходимую степень возбуждения и торможения иннервирующих эти мышцы мотонейронов, чтобы возникающие мышечные сокращения обеспечивали только нужное движение. Нервная регуляция соответствия позы и движения, их правильного сопряжения − одна из важнейших функций двигательных центров. В спинном мозге сенсорные афферентные волокна образуют множество связей с мотонейронами, главным образом − через интернейроны. От того, какие связи задействованы, зависит активация или торможение определенных движений. Нейронные цепи, составляющие рефлекторные дуги, на которых основаны спинальные рефлексы, представляют собой конкретные анатомические образования. Под спинальным рефлексом (reflexus (лат.) − отражение, загибание назад) правильнее понимать изменение нейронной активности, вызываемое спинальными афферентами и приводящее к запуску или торможению движения. Сегментарный аппарат спинного мозга − это совокупность функционально взаимосвязанных нервных структур, обеспечивающих сложную и разнообразную рефлекторную деятельность спинного мозга, морфологической основой которой являются простые рефлекторные дуги. В состав его входят: заднекорешковые волокна, вставочные нейроны и их интерсегментарные проводники, крупные нейроны собственных ядер передних рогов и начальная часть их аксонов, составляющих передние корешковые волокна. Рефлекторная дуга состоит из афферентов, центральных нейронов и мотонейронов, следовательно активируемых во время рефлекса. Промежуток времени между поступлением стимула и реакцией эффектора называется временем рефлекса. В большинстве случаев оно определяется в основном временем проведения в афферентных и эфферентных путях, а также в центральных участках рефлекторной дуги. Сюда добавляется время, необходимое для (1) преобразования рецептором стимула в распространяющийся импульс; (2) передачи возбуждения в синапсах центральных нейронов (синаптическая задержка); (3) передачи его от эфферентных путей к эффектору (например, для генерирования потенциала концевой пластинки) и (4) активации эффектора в результате возбуждения мембраны (например, для электромеханического сопряжения). * Рефлекторная дуга Рефлекс растяжения, вызываемый растяжением мышцы. Ia-волокна мышечных веретен образуют возбуждающие синапсы на гомонимных α-мотонейронах (т.е. мотонейронах их собственных мышц). Активация первичных окончаний мышечных веретен при растяжении мышцы ведет, таким образом, к возбуждению гомонимных мотонейронов. В этом рефлексе участвует только один синапс − между волокном Ia и гомонимным мотонейроном − поэтому он называется моносинаптическим рефлексом растяжения мышцы (используется также термин миотатический рефлекс). Это самый простой пример полной рефлекторной дуги (пример: коленный рефлекс). Клинически важные примеры таких рефлекторных тестов включают: растяжение мышцы, закрывающей рот, путем постукивания по подбородку (рефлекс закрывания рта); растяжение двуглавой мышцы плеча путем постукивания по ее сухожилию около локтя; растяжение трехглавой мышцы голени путем постукивания по пяточному сухожилию (рефлекс ахиллова сухожилия). Такого рода рефлексы, вызываемые постукиванием по сухожилию, клиницисты называют Т−рефлексами (от tendon − сухожилие). Последовательная проверка рефлексов растяжения представляет интерес в связи с тем, что их дуги проходят через разные сегменты спинного мозга, а значит, нарушения определенных рефлексов могут показать, на каком его уровне локализован патологический процесс. Клиническим показателем служит не столько ее абсолютная величина, сколько разница между рефлексами с правой и с левой стороны тела или аномалии некоторых из них, выявляемые при сравнении с общим рефлекторным поведением. Облегчение Т-рефлексов. Н−рефлексы. Моносинаптический рефлекс растяжения у человека можно также вызвать электрической стимуляцией афферентов Ia в составе мышечного нерва; в этом случае его называют Н-рефлексом (по имени Р. Hoffmann). Период молчания. После Т- или Н-рефлекса тонус мышц резко падает на короткий период времени (100−500 мс). Этот пострефлекторный период молчания обусловлен сочетанием, по крайней мере, четырех факторов: 1. Синхронное рефлекторное сокращение снимает напряжение мышечных веретен и, следовательно, ослабляет или прекращает тоническую возбуждающую афферентную стимуляцию, обеспечиваемую их первичными окончаниями. 2. Рефлекторное сокращение активирует сухожильные органы Гольджи, а это оказывает тормозное действие на гомонимные мотонейроны. 3. Синхронное возбуждение мотонейронов на короткое время усиливает торможение, производимое клетками Реншоу. 4. Гиперполяризующие следовые потенциалы после потенциалов действия в мотонейронах, участвующих в рефлексе, временно снижают их возбудимость. Реципрокное торможение агонистов афферентами Ia. Волокна Ia образуют не только моносинаптические возбуждающие связи с гомонимными мотонейронами в составе дуг рефлекса растяжения, но и тормозные связи с мотонейронами-антагонистами. Тормозная рефлекторная дуга включает центральный интернейрон, следовательно относится к дисинаптическому типу − в ней два центральных синапса: возбуждающий между волокнами Ia и интернейронами и тормозной между аксонами интернейронов и мотонейронами. Поскольку это самая короткая тормозная рефлекторная дуга среди всех известных, в ее случае говорят о прямом торможении. Однако предпочтительнее термин реципрокное антагонистическое торможение, подчеркивающий, что за счет такой рефлекторной дуги мотонейроны каждой мышцы-антагониста в паре (например, сгибателя и разгибателя в данном суставе) тормозятся во время активации другого компонента пары. В функциональном плане реципрокное антагонистическое торможение усиливает результат активации волокон Ia, вызывающей или усиливающей сокращение одной из мышц сустава, путем одновременного ослабления возбуждающей стимуляции мышцы, которая противодействовала бы этому сокращению. Поскольку и в последней мышце есть волокна Ia, связанные с ее антагонистом, вся система состоит из четырех рефлекторных дуг. При их активации пассивным (вызванным внешним воздействием) изменением положения сустава рефлексы этому изменению противодействуют, способствуя поддержанию исходной длины мышцы. Так, если под действием силы тяжести коленный сустав начинает сгибаться, растяжение мышечных веретен разгибателя (первый рефлекс) усиливает возбуждение гомонимных мотонейронов и (второй рефлекс) торможение мотонейронов сгибателя. Кроме того, уменьшение растяжения мышечных веретен сгибателя ослабляет (третий рефлекс) возбуждение гомонимных мотонейронов и (четвертый рефлекс) реципрокное торможение мотонейронов разгибателя (это называют растормаживанием). В конечном счете мотонейроны разгибателей возбуждаются, а сгибателей − тормозятся. Все четыре рефлекторные дуги в совокупности образуют систему регуляции длины мышцы. Функции γ-петли. Сокращения мышцы, вызываемые активацией интрафузальных волокон. При возбуждении первичных окончаний мышечных веретен в результате сокращения интрафузальных волокон, вызванного γ-мотонейронами, активность волокон Ia непосредственно возбуждает гомонимные мотонейроны, как и при растяжении мышцы. Таким образом, мышечные веретена могут вызывать сокращения экстрафузальных волокон в двух случаях: (1) при растяжении мышцы; (2) при активации аксонов γ-мотонейронов, приводящей к сокращению интрафузальных волокон. Второй механизм получил название γ -петли . Преимущество прямой активации α-мотонейронов супраспинальными центрами − короткий латентный период, но ее недостаток состоит в нарушении тонкого равновесия системы регуляции мышечной длины, включающей рефлекс растяжения. В результате растяжение веретен соответствующей мышцы может стать недостаточным (подпороговым) или избыточным (насыщающим). В отличие от этого, активация γ-петли обеспечивает укорочение мышцы с минимальными (если они вообще происходят) изменениями частоты импульсации афферентов мышечных веретен. |