биологи 5. А Она, прежде всего, отличается целостностью
 Скачать 1.49 Mb. Скачать 1.49 Mb.
|

|
 Полиморфизм человеческих популяций. Генетический груз.
Полиморфизм человеческих популяций. Генетический груз.| страна | АДН1 | АДГ2 |
| Англия | 0,97 | 0,03 |
| Швеция | 0,89 | 0,11 |
| Германия | 0,97 | 0,03 |
| Япония | 0,35 | 0,65 |
Однако есть другой фермент, который осуществляет расщепление альдегида и от его активности также зависит переносимость человеком алкоголя.
Генетический полиморфизм широко распространен и лежит в основе наследственной предрасположенности к заболеваниям. Однако болезни наследственных предрасположений проявляются лишь при взаимодействии генов и среды. Условия среды – недостаток или избыток питательных веществ, наличие психогенных факторов, токсических веществ и др. Клиническое течение болезней может быть разнообразно. Чем больше воздействие факторов среды, тем больше лиц больных с предрасположенностью к данному заболеванию. Болезни протекают тяжелее (гипертония, ревматизм, сахарный диабет и другие),
Есть моногенные и полигенные болезни.
Моногенные болезни наследственного предрасположения – наследственные заболевания, проявляющиеся из-за мутации одного гена или проявляющиеся при действии определенного фактора среды (аутосомно-рецессивные или сцепленные с Х-хромосомой).
Проявляются при воздействии факторов:
- физических;
- химических;
- пищевых;
- загрязнения среды.
Парамиотомия – в сырую погоду происходят тонические спазмы мышц при холоде, под влиянием тепла – проходят. Болезнь связана с термочувствительным белком. Реакция проявляется в младенчестве и не изменяется на протяжении жизни человека.
Пигментная ксеродерма - веснушчатая кожа особого типа. Проявляется в 4-6 лет. Дети не переносят УФ-свет возникают злокачественные опухоли, такие дети умирают от метастаз еще до 15 лет. Не переносят также и гамма-лучей.
Синдром Блюма. Пигментная «бабочка» на лице, маленький рост, удлиненная голова. Евреи, поляки, беларусы, австрийцы. Погибают до 18 лет. Не переносят УФ-облучения, гамма-лучей.
Альфа-1 антитрипсин при загрязнении воздуха, табачном дыме проявляется острой закупоркой бронхов или циррозом печени.
У европеоидов люди, не переносящие молоко, составляют 10-20%, в Африке – 70-80%.
Влияние лекарственных средств: сульфаниламидные препараты провоцируют заболевания крови.
Есть полигенные болезни наследственного происхождения – такие болезни, которые возникают при действии многих факторов (мультифакториальные) и в результате взаимодействия многих генов. Установить диагноз в таком случае очень сложно, т.к. действует много факторов, и появляется новое качество при взаимодействии факторов.
Широкий полиморфизм помогает популяции приспосабливаться к условиям среды. У здоровых людей нет противоречия между средой и генотипом, если возникает это противоречие - проявляются болезни наследственного предрасположения. Любые классификации болезней включают группу подобных заболеваний.
Процесс видообразования с участием такого фактора, как естественный отбор, создает разнообразие живых форм, приспособленных к условиям обитания. Среди разных генотипов, возникающих в каждом поколении благодаря резерву наследственной изменчивости и перекомбинации аллелей, лишь ограниченное число обусловливает максимальную приспособленность к конкретной среде. Можно предположить, что дифференциальное воспроизведение этих генотипов в конце приведет к тому, что генофонды популяций будут представлены лишь «удачными» аллелями и их комбинациями. В итоге произойдет затухание наследственной изменчивости и повышение уровня гомозиготности генотипов.
В природных популяциях, однако, наблюдается противоположное состояние. Большинство организмов являются высокогетерозиготными. Отдельные особи гетерозиготны частично по разным локусам, что повышает суммарную гетерозиготность популяции. Так, методом электрофореза на 126 особях рачка Euphausia superba, представляющего главную пищу китов в антарктических водах, изучали 36 локусов, кодирующих первичную структуру ряда ферментов. По 15 локусам изменчивость отсутствовала. По 21 локусу имелось по 3—4 аллеля. В целом в этой популяции рачков 58% локусов были гетерозиготными и имели по 2 аллеля и более. В среднем у каждой особи по 5,8% гетерозиготных локусов. Средний уровень гетерозиготности у растений составляет 17%, беспозвоночных — 13,4, позвоночных — 6,6%. У человека этот показатель равен 6,7%. Столь высокий уровень гетерозиготности нельзя объяснить только мутациями в силу относительной их редкости.
Наличие в популяции нескольких равновесно сосуществующих генотипов в концентрации, превышающей по наиболее редкой форме 1%1, называют полиморфизмом. Наследственный полиморфизм создается мутациями и комбинативной изменчивостью. Он поддерживается естественным отбором и бывает адаптационным (переходным) и гетерозиготным (балансированным).
Адаптационный полиморфизм возникает, если в различных, но закономерно изменяющихся условиях жизни отбор благоприятствует разным генотипам. Так, в популяциях двухточечных божьих коровок Adalia bipunctata при уходе на зимовку преобладают черные жуки, а весной—красные (рис. 11.7). Это происходит потому, что красные формы лучше переносят холод, а черные интенсивнее размножаются в летний период.
Балансированный полиморфизм возникает, если отбор благоприятствует гетерозиготам в сравнении с рецессивными и доминантными гомозиготами. Так, в опытной численно равновесной популяции плодовых мух Drosophila melanogaster, содержащей поначалу много мутантов с более темными телами (рецессивная мутация ebony), концентрация последних быстро падала, пока не стабилизировалась на уровне 10% (рис. 11.8). Анализ показал, что в созданных условиях гомозиготы по мутации ebony и гомозиготы по аллелю дикого типа менее жизнеспособны, чем гетерозиготные мухи. Это и создает состояние устойчивого полиморфизма по соответствующему локусу. Явление селективного преимущества гетерозигот называют сверхдоминантностью. Механизм положительного отбора гетерозигот различен. Правилом является зависимость интенсивности отбора от частоты, с которой встречается соответствующий фенотип (генотип). Так, рыбы, птицы, млекопитающие предпочитают обычные фенотипические формы добычи, «не замечая» редких.
В качестве примера рассмотрим результаты наблюдений, выполненных на обыкновенной наземной улитке Cepaea nemoralis, раковина у которой бывает желтая, различных оттенков коричневого цвета, розовая, оранжевая или красная. На раковине может быть до пяти темных полос. При этом коричневая окраска доминирует над розовой, а они обе — над желтой. Полосатость является рецессивным признаком. Улитки поедаются дроздами, использующими камень как наковальню, чтобы разбить раковину и добраться до тела моллюска. Подсчет числа раковин разной окраски вокруг таких «наковален» показал, что на траве или лесной подстилке, фон которых достаточно однороден, добычей птиц чаще оказывались улитки с розовой и полосатой раковиной. На пастбищах с грубыми травами или в живых изгородях с более пестрым фоном чаще поедались улитки, раковины которых окрашены в светлые тона и не имели полос.
Самцы относительно редких генотипов могут иметь повышенную конкурентоспособность за самок. Селективное преимущество гетерозигот обусловливается также явлением гетерозиса. Повышенная жизнеспособность межлинейных гибридов отражает, по-видимому, результат взаимодействия аллельных и неаллельных генов в системе генотипов в условиях гетерозиготности по многим локусам. Гетерозис наблюдается в отсутствие фенотипического проявления рецессивных аллелей. Это сохраняет скрытыми от естественного отбора неблагоприятные и даже летальные рецессивные мутации.
В силу разнообразия факторов среды обитания естественный отбор действует одновременно по многим направлениям. При этом конечный результат зависит от соотношения интенсивности разных векторов отбора. Конечный результат естественного отбора в популяции зависит от наложения многих векторов отборов и контротборов. Благодаря этому достигается одновременно и стабилизация генофонда, и поддержание наследственного разноообразия.
Балансированный полиморфизм придает популяции ряд ценных свойств, что определяет его биологическое значение. Генетически разнородная популяция осваивает более широкий спектр условий жизни, используя среду обитания более полно. В ее генофонде накапливается больший объем резервной наследственной изменчивости. В результате она приобретает эволюционную гибкость и может, изменяясь в том или ином направлении, компенсировать колебания среды в ходе исторического развития.
В генетически полиморфной популяции из поколения в поколение рождаются организмы генотипов, приспособленность которых неодинакова. В каждый момент времени жизнеспособность такой популяции ниже уровня, который был бы достигнут при наличии в ней лишь наиболее «удачных» генотипов. Величину, на которую приспособленность реальной популяции отличается от приспособленности идеальной популяции из «лучших» генотипов, возможных при данном генофонде, называют генетическим грузом. Он является своеобразной платой за экологическую и эволюционную гибкость. Генетический груз — неизбежное следствие генетического полиморфизма.
Эволюция адаптации – основной результат действия естественного отбора. Классификация адаптации: морфологические, физиолого-биохимические, этологические, видовые адаптации: конгруэнции и кооперации. Относительность органической целесообразности.
Ответ: Адаптацией считается любая особенность особи, популяции, вида или сообщества организмов, которая способствует успеху в конкуренции и обеспечивает устойчивость к абиотическим факторам. Это позволяет организмам существовать в данных условиях среды и оставлять потомство. Критериями адаптации являются: жизнеспособность, конкурентоспособность и фертильность.
Виды адаптации
Все адаптации делят на аккомодации и эволюционные адаптации. Аккомодации представляют собой обратимый процесс. Они возникают при резком изменении условий среды. Например, при переселении животные попадают в новую для них обстановку, но постепенно привыкают к ней. Например, человек, переселившийся из средней полосы в тропики или на Крайний Север, некоторое время испытывает дискомфорт, но со временем привыкает к новым условиям. Эволюционная адаптация необратима и возникшие изменения генетически закрепляются. Сюда относят все приспособления, на которые действует естественный отбор. Например, покровительственная окраска или быстрый бег.
Морфологические адаптации проявляются в преимуществах строения, покровительственной окраске, предостерегающей окраске, мимикрии, маскировке, приспособительном поведении.
Преимущества строения – это оптимальные пропорции тела, расположение и густота волосяного или перьевого покрова и т.п. Хорошо известен облик водного млекопитающего – дельфина.
Мимикрия – это результат гомологичных (одинаковых) мутаций у разных видов, которые помогают выжить незащищённым животным.
Маскировка – приспособления, при которых форма тела и окраска животных сливаются с окружающими предметами
Физиологические адаптации - приобретение специфических особенностей обмена веществ в разных условиях среды. Они обеспечивают функциональные преимущества организма. Их условно разделяют на статические (постоянные физиологические параметры — температура, водно-солевой баланс, концентрация сахара и т. п.) и динамические (адаптации к колебаниям действия фактора — изменение температуры, влажности, освещенности, магнитного поля и т. п.). Без такой адаптации невозможно поддержание устойчивого обмена веществ в организме в постоянно колеблющихся условиях внешней среды. Приведем некоторые примеры. У наземных амфибий большое количество воды теряется через кожу. Однако многие их виды проникают даже в пустыни и полупустыни. Очень интересны приспособления, развивающиеся у ныряющих животных. Многие из них могут сравнительно долго обходиться без доступа кислорода. Например, тюлени ныряют на глубину 100—200 и даже 600 метров и находятся под водой 40—60 минут. Поразительно чувствительны органы химического чувства насекомых.
Биохимические адаптации обеспечивают оптимальное течение биохимических реакций в клетке, например, упорядочение ферментативного катализа, специфическое связывание газов дыхательными пигментами, синтез нужных веществ в определенных условиях и т. п.
Этологические адаптации представляют собой все поведенческие реакции, направленные на выживание отдельных особей и, следовательно, вида в целом. Такими реакциями являются:
- поведение при поиске пищи и полового партнера,
- спаривание,
- выкармливание потомства,
- избегание опасности и защита жизни в случае угрозы,
- агрессия и угрожающие позы,
- незлобивость и многие другие.
Некоторые поведенческие реакции наследуются (инстинкты), другие приобретаются в течение жизни (условные рефлексы).
Видовые адаптации обнаруживаются при анализе группы особей одного вида, по своему проявлению они весьма разнообразны. Основными из них являются различные конгруэнции, уровень мутабильности, внутривидовой полиморфизм, уровень численности и оптимальная плотность населения.
Конгруэнции представляют собой все морфофизиологические и поведенческие особенности, которые способствуют существованию вида как целостной системы. Репродуктивные конгруэнции обеспечивают размножение. Некоторые из них непосредственно связаны с репродукцией (соответствие половых органов, приспособления к вскармливанию и др.), тогда как другие лишь опосредованно (различные сигнальные признаки: зрительные - брачный наряд, ритуальное поведение; звуковые - пение птиц, рев самца оленя во время гона и др.; химическими - различные аттрактанты, например, феромоны насекомых, выделения у парнокопытных, кошачьих, собачьих и др.).
К конгруэнциям относят все формы внутривидовой кооперации,— конституциональной, трофической и репродуктивной. Конституциональная кооперация выражается в согласованных действиях организмов в неблагоприятных условиях, которые повышают шансы на выживание. Зимой пчелы собираются в шар, и выделяемое ими тепло расходуется на совместное согревание. При этом самая высокая температура будет в центре шара и особи с периферии (где холоднее) будут постоянно стремиться туда. Таким образом происходит постоянное перемещение насекомых и они совместными усилиями благополучно перезимуют. Также сбиваются в тесную группу пингвины во время насиживания, овцы в холодное время и др.
Трофическая кооперация состоит в объединении организмов с целью добывания пищи. Совместная деятельность в этом направлении делает процесс более продуктивным. Например, стая волков гораздо эффективнее охотится, нежели отдельная особь. При этом у многих видов имеет место разделение обязанностей - одни особи отделяют выбранную жертву от основного стада и гонят ее в засаду, где затаились их сородичи и т. д. У растений подобная кооперация выражается в совместном затенении почвы, что способствует удержанию в ней влаги.
Репродуктивная кооперация повышает успешность размножения и способствует выживанию потомства. У многих птиц особи собираются на токовищах, и в таких условиях облегчается поиск потенциального партнера. То же самое происходит на нерестилищах, лежбищах ластоногих и др. Вероятность опыления у растений повышается, когда они растут группами и расстояние между отдельными особями невелико.
Закон органической целесообразности, или закон Аристотеля
1. Чем глубже и разностороннее изучает наука живые формы, тем полнее раскрывается их целесообразность, т. е. целенаправленный, гармоничный, как бы разумный характер их организации, индивидуального развития и отношения с окружающей средой. Органическая целесообразность раскрывается в процессе познания биологической роли конкретных особенностей живых форм.
2. Целесообразность присуща всем видам. Она выражается в тонком взаимном соответствии структур и назначения биологических объектов, в приспособленности живых форм к условиям жизни, в естественной целенаправленности особенностей индивидуального развития, в приспособительном характере форм существования и поведения биологических видов.
3. Органическая целесообразность, ставшая предметом анализа античной науки и служившая основанием для телеологических и религиозных истолкований живой природы, получила материалистическое объяснение в учении Дарвина о творческой роли естественного отбора, проявляющейся в адаптивном характере биологической эволюции.
Такова современная формулировка тех обобщений, истоки которых восходят к Аристотелю, выдвинувшему представления о целевых причинах.
Изучение конкретных проявлений органической целесообразности одна из важнейших задач биологии. Выяснив, для чего служит та или иная особенность исследуемого биологического объекта, в чем биологическое значение этой особенности, мы благодаря эволюционной теории Дарвина приближаемся к ответу на вопрос, почему и каким образом она возникла. Рассмотрим проявления органической целесообразности на примерах, относящихся к различным областям биологии.
В области цитологии яркий, наглядный пример органической целесообразности — деление клеток у растений и животных. Механизмы эквационного (митоз) и редукционного (мейоз) деления обусловливают постоянство числа хромосом в клетках данного вида растений или животных. Удвоение диплоидного набора в митозе обеспечивает сохранение постоянства числа хромосом в делящихся соматических клетках. Гаплоидизация хромосомного набора при образовании половых клеток и восстановление его при образовании зиготы в результате слияния половых клеток обеспечивают сохранение числа хромосом при половом размножении. Отклонения от нормы, приводящие к полиплоидизации клеток, т. е. к умножению числа хромосом против нормального, отсекаются стабилизирующим действием естественного отбора или служат условием генетического обособления, изоляции полиплоидной формы с возможным превращением ее в новый вид. При этом в действие вновь вступают цитогенетические механизмы, обусловливающие сохранение хромосомного набора, но уже на новом, полиплоидном, уровне.
В процессе индивидуального развития многоклеточного организма происходит образование клеток, тканей и органов различного функционального назначения. Соответствие этих структур их назначению, их взаимодействие в процессе развития и функционирования организма — характерные проявления органической целесообразности.
Обширную область примеров органической целесообразности представляют приспособления для размножения и распространения живых форм. Назовем некоторые из них. Например, споры бактерий обладают высокой устойчивостью к неблагоприятным условиям среды. Цветковые растения приспособлены к перекрестному опылению, в частности с помощью насекомых. Плоды и семена ряда растений приспособлены к распространению с помощью животных. Половые инстинкты и инстинкты заботы о потомстве характерны для животных самого различного уровня организации. Строение икры и яиц обеспечивает развитие животных в соответствующей среде. Молочные железы обеспечивают полноценное питание потомства у млекопитающих.
Адаптация - это приспособление живых систем к тем или иным условиям среды обитания. Все виды адаптации - это результат действия эволюционного процесса на основе естественного отбора. Адаптации могут возникать в абиотических и биотических факторов и быть направлены на повышение устойчивости организмов.
Пути адаптации могут быть разными, а именно:
1. Морфологические (защитная окраска, колючки, толстая кутикула, волосяной покров, жировой слой и т.д..).
2. Физиологические адаптации (устойчивость физиологических параметров: постоянная температура тела, содержание кислорода, углекислого газа, содержание сахара в крови и т.д..).
3. Биохимические адаптации (постоянство биохимических процессов).
4. Этологические адаптации (поведенческие реакции как адаптации организма).
С позиций учения Ч. Дарвина были объяснены разнообразные адаптации живых существ к условиям среды обитания. В частности, изучены различные виды защитных расцветок, формы тела и поведения организмов, которые делают их менее уязвимыми для врагов.
Животные с защитными окраской и формой тела в случае опасности, маскируясь, принимают определенную позу. Например, замерев, гусеницы бабочек-пядениц или тропические насекомые палочники становятся похожими на сухие сучки и совершенно незаметны на растениях. В умеренных широтах благодаря сезонным линькой млекопитающие и птицы приобретают темный летнего или светлого зимнего окраску, что соответствует фону окружающей среды. Некоторые животные способны изменять окраску в зависимости от фона среды - камбала, осьминоги, хамелеоны и другие. По явления демонстрации, наоборот, окраску и поведение животных делают их заметными на фоне окружающей среды. Предупредительная и угрожающе окраску и поведение сигнализируют врагам о неис-тивнисть таких животных (колорадский жук, солнышко) или их хорошую защищенность (жалоносных осы, пчелы, ядовитые змеи)
Хорошим примером адаптации к условиям среды является мимикрия. Мимикрия (от греч. Мимикос - подражательный) - это способность к уподоблению по окраске, форме или поведением организмов одного вида (моделей) особями другого (имитаторами). Две формы мимикрии открыли английский ученый Г. Бейтс и немецкий - Ф. Мюллер.
По бейтсивськои мимикрии хуже защищен вид уподобляется хорошо защищенном, а за мюллеривськои - несколько защищенных видов подражают друг друга, образуя своеобразное кольцо: их враги, выработав рефлекс отвращения к одному из видов «кольца», не трогают также и других.
Примером бейтсивськои мимикрии служат некоторые тропические бабочки-белянки, подобные несъедобных для птиц бабочек других семейств. Различные бабочки, мухи, жуки подражают ядовитых ос и пчел, неядовитые змеи - ядовитых т.д.. Мюллеривську мимикрию иллюстрируют ядовитые членистоногие с предупредительным красной окраской с черными пятнами (солнышка, клоп-солдатик и др.). Или желто-черным (различные виды ос, некоторые пауки).
Мимикрия у растений - это отдельные приспособления, напоминающие другие виды. Так, у некоторых растений цветки не имеют нектарники, однако они напоминают цветки красивых нектароносиив и этим привлекают насекомых-опылителей. Цветки некоторых тропических орхидей по форме и окраске напоминают самок некоторых видов бабочек. Самцы этих насекомых опыляют их в случае спаривания.
Следствием адаптационных процессов является существование аналогичных и гомологичных органов, а также рудиментов и атавизмов.
Гомология (от греч. Гомология - соразмерность) - это соответствие общего плана строения органов разных видов, обусловлена их общим происхождением.
Вследствие адаптации к различным условиям жизни гомологичные органы у разных видов могут значительно различаться между собой, и единство их происхождения устанавливают только на основании исследований внутреннего строения, индивидуального развития, данных палеонтологии и т.д..
Примерами гомологичных органов являются передние конечности (нога, крыло, рука, ласты т.д.) различных позвоночных животных или видоизменения корня высших растений (корнеплод, корневая клубень т.п.). У особей разных таксономических групп может наблюдаться конвергенция - появление различных признаков в результате приспособления различных организмов к тем же условиях проживания (бабочки и птицы, киты и рыбы). Так возникают аналогичные органы.
Аналогии (от греч. Аналогия - сходство) - это сходство строения органов различного происхождения, которые выполняют одинаковые функции.
Дивергенция — процесс расхождения признаков в процессе адаптации к разным условиям существования.
При дивергенции сходство между организмами объясняется общностью их происхождения, а различия — приспособлением к разным условиям среды. Примером дивергенции форм является возникновение разнообразных по морфофизиологическим особенностям вьюрков от одного или немногих предковых видов на Галапагосских островах. Расхождение внутривидовых форм и видов по разным местообитаниям определяется конкуренцией в борьбе за одинаковые условия, выход из которых и заключается в расселении по разным экологическим нишам.
Дивергенция может быть одним из путей видообразования, когда в результате действия элементарных эволюционных факторов популяции и группы популяций приобретают и сохраняют признаки, все более заметно отличающие их от родительского вида, что может приводить к распаду исходного вида на два и более дочерних. В конечном счете дивергенция приводит к формированию более крупных таксонов — родов, семейств и т.д. — которые продолжают расходиться.
Результат дивергенции: гомологичные органы у родственных форм приобретают разные функции и разное строение
Конвергенция — процесс эволюционного развития неродственных групп в сходном направлении и приобретение ими сходных признаков в процессе адаптации к одинаковым условиям среды.
Классическим примером конвергентного развития считается возникновение сходных форм тела у акуловых (первичноводные формы), ихтиозавров и китообразных (вторичноводные формы). При конвергентном развитии сходство между неродственными организмами бывает всегда только внешним (эволюционным изменениям в одном направлении подвергаются внешние признаки как результат приспособления к одинаковым условиям среды). По форме тела акула и дельфин сходны, но по таким существенным чертам, как строение кожных покровов, черепа, мускулатуры, кровеносной системы, дыхательной и других систем, эти группы позвоночных различны.
При конвергентном способе эволюции возникают аналогичные органы.
Параллелизм (параллельное развитие) — такое явление, когда сходство признаков возникает в результате сходного (но независимого) изменения гомологичных структур.
Причиной параллелизма, по-видимому, является высокая вероятность похожих мутаций одних и тех же генов у разных видов. То же явление в рамках генетики описывает закон гомологических рядов наследственной изменчивости Н.И. Вавилова. Согласно этому закону, родственные виды характеризуются сходными рядами наследственной изменчивости. Поэтому сходные признаки могут возникать у родственны видов независимо — параллельно. Так, некоторые виды бабочек, принадлежащие к разным родам, имеют глазки на крыльях, а их ближайшие родственники глазков не имеют. Этот признак независимо возникал у разных видов, а не был унаследован от общих предков. От общих предков все бабочки унаследовали гены, мутации которых приводят к образованию глазков.
Другой пример того же рода — независимое возникновение саблезубых форм у млекопитающих. Широко известны вымершие саблезубые кошки, в частности, саблезубый тигр. Саблезубые кошки — это подсемейство семейства кошачьих, но саблезубость встречалась и в других отрядах млекопитающих, ныне вымерших (например, псевдо-саблезубые кошки). А в палеонтологической летописи Австралии есть сумчатый саблезубый волк. Таким образом, один и тот же признак в разных группах млекопитающих возник независимо.
Взависимости от того, изменяется ли уровень организации в эволюционирующих группах, выделяют два основных типа эволюции: аллогенез и арогенез.
При аллогенезе у всех представителей данной группы сохраняются без изменения основные черты строения и функционирования систем органов, благодаря чему уровень организации их остается прежним. Аллогенная эволюция
53

происходит в пределах одной адаптивной зоны — совокупности экологических ниш, различающихся в деталях, но сходных по общему направлению действия основных средовых факторов на организм данного типа. Интенсивное заселение конкретной адаптивной зоны достигается благодаря возникновению у организмов идиоадаптаций — локальных морфофизиологических приспособлений к определенным условиям существования. Пример аллогенеза с приобретением идиоадаптаций к разнообразным условиям обитания в отряде насекомоядных млекопитающих см. на рис. 13.2.
Рис. 13.2. Аллогенез в отряде насекомоядных млекопитающих
Наземные формы: а—прыгунчик, б—землеройка, в—еж; земноводные формы: г— кутора, д—выдровая землеройка, е—выхухоль; роющие формы: ж—крот, з— златокрот
54
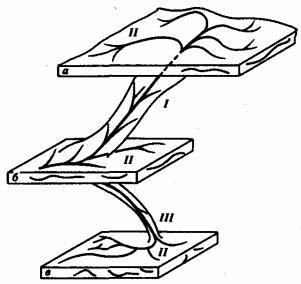
Арогенез — такое направление эволюции, при котором у некоторых групп внутри более крупного таксона появляются новые морфофизиологические особенности, приводящие к повышению уровня их организации. Эти новые прогрессивные черты организации называют ароморфозами. Ароморфозы позволяют организмам заселять принципиально новые, более сложные адаптивные зоны. Так, арогенез ранних земноводных был обеспечен появлением у них таких основных ароморфозов, как пятипалые конечности наземного типа, легкие и два круга кровообращения с трехкамерным сердцем. Завоевание адаптивной зоны с более сложными для жизни условиями (наземной по сравнению с водной, воздушной по сравнению с наземной) сопровождается активным расселением в ней организмов с появлением у них локальных идиоадаптаций к различным экологическим нишам.
Таким образом, периоды арогенной эволюции группы могут сменяться периодами аллогенеза, когда в результате возникающих идиоадаптаций новая адаптивная зона заселяется и используется наиболее эффективно. Если в ходе филогенеза организмы осваивают более 49
простую по сравнению с исходной адаптивную зону, например, переходя к прикрепленному неподвижному образу жизни или становись паразитами, то они претерпевают морфофизиологический регресс, или дегенерацию, утрачивая часть прогрессивных черт, которыми обладали их предки, и оказываясь на более низком уровне организации. Нередко и новая, более простая, адаптивная зона дает возможность широких идиоадаптаций к не менее разнообразным условиям обитания в ней, т.е. обеспечивает аллогенез (рис. 13.3).
Рис. 13.3. Направления эволюции групп:
I — арогенез, II — аллогенез, III — дегенерация; а, б, в — адаптивные зоны
55
Современный органический мир характеризуется огромным многообразием форм, отличающихся как по уровню организации, так и по разнообразию локальных адаптации именно благодаря смене типов эволюции, протекающей своеобразно в каждой из эволюционирующих групп. Наряду с высокоорганизованными формами существуют примитивные, свободноживущие и паразитические организмы, возникшие эволюционно относительно недавно, и реликтовые группы, время бурного адаптационного процесса которых давно прошло.
13.1.3. Формы эволюции групп
Существуют две элементарные формы филогенеза: филетическая и дивергентная эволюция.
Филетическая эволюция — это изменения, происходящие в одном филогенетическом стволе, эволюционирующем во времени как единое целое. Реконструированы филогенетические ряды слона, лошади и других организмов, в которых можно наблюдать постепенное нарастание количества и степени выраженности признаков, характерных для современных форм (рис. 13.4).
Дивергентная эволюция заключается в образовании на основе одной предковой группы двух или нескольких производных (рис. 13.5). Она приводит к дифференциации более крупных таксонов на более мелкие, например классов на отряды, родов на виды.
Как филетическая, так и дивергентная эволюция протекают на общей генетической базе, поэтому между организмами сохраняется более или менее выраженное генотипическое и морфофункциональное сходство.
Сопоставление филогенеза в разных группах позволяет выделить и некоторые общие закономерности соотносительной эволюции. Так, при попадании в одну и ту же среду обитания двух или более филогенетических групп неродственных организмов у них обычно проявляется конвергенция признаков. При этом сходные
экологические задачи они решают сходным образом. Конвергентные адаптации возникают в этом случае на разной генетической основе, затрагивают в первую очередь поверхностные признаки, не распространяясь на общий план строения и наиболее существенные черты организации соответствующих групп. Примером конвергентной эволюции являются форма тела и особенности локомоции в воде у акуловых рыб, водных пресмыкающихся — ихтиозавров, костистых рыб, пингвинов, ластоногих и китообразных млекопитающих, внутреннее строение которых полностью соответствует особенностям, характерным для классов, к которым они относятся (рис. 13.6).
Другая форма соотносительной эволюции — параллелизм — реализуется в двух или нескольких группах, связанных более или менее отдаленным родством, которое основано на дивергенции от общего предка. В связи с общностью части генофондов, унаследованных от предков, у них возникают сходные адаптации в условиях действия факторов отбора в одинаковом направлении. Параллельное филетическое развитие двух родственных групп обеспечивается реализацией закона гомологических рядов (см. разд. 13.3.5). Примером параллелизма является эволюция одногорбого и двугорбого верблюдов соответственно в Африке и Центральной Азии, имеющих сходные адаптации к жизни в пустынях
Параллелизм можно наблюдать и в родственных группах организмов, разобщенных временем. Так, во второй половине кайнозойской эры в семействе кошачьих такой признак, как саблезубость (гипертрофия клыков верхней челюсти), возникал по крайней мере четырежды у представителей