Б1 Понятие биомембрана
 Скачать 2.06 Mb. Скачать 2.06 Mb.
|

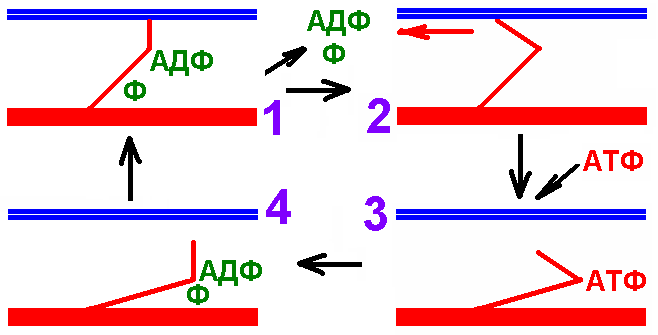
50. Рабочий цикл миозиновых (поперечных) мостиков
Головка миозина совершает около 5 циклов в сек. Когда одни головки тянут, другие в это время свободны и готовы вступить в очередной цикл. Следующие друг за другом гребковые движения стягивают тонкие нити к центру саркомера, тянут за собой Z-линии, - сокращение саркомера и укорочение. Циклы повторяться, пока в межфибриллярном пространстве есть Ca2+. ( уходит из межфибриллярного пространства в цистерны) или АТФ образуются не быстро, не обеспечивая энергией поперечные мостики. Напряжение, развиваемое мышечным волокном, зависит от числа замкнутых поперечных мостиков, а скорость развития напряжения — от числа замкнутых поперечных мостиков, образуемых в единицу времени, т.е. от скорости их прикрепления к актину. По-видимому, даже при максимальном возбуждении только часть поперечных мостиков эффективно связана с актиновыми миофиламентами. При повышении скорости укорочения мышцы число прикрепленных поперечных мостиков в каждый момент времени уменьшается. Это и объясняет падение силы сокращения мышцы с увеличением скорости ее укорочения. Если же мышца не укорачивается (при изометрическом сокращении), напряжение в мышечных волокнах создается за счет повторного прикрепления поперечных мостиков на одних и тех же фиксированных участках актиновых нитей. В этом случае никакой внешней работы не совершается, а энергия (АТФ) расходуется на поддержание напряжения мышцы. Тяга, развиваемая сократительными элементами в миофибриллах, передается сарколемме и через нее — на сухожилия мышц. Период химико-механических изменений в сократительном миофибриллярном аппарате мышечного волокна, на протяжении которого в нем возникает и поддерживается активная механическая тяга, определяющая процесс сокращения, называется периодом активного состояния. цикл миозиновых мостиков гладкого миоцита
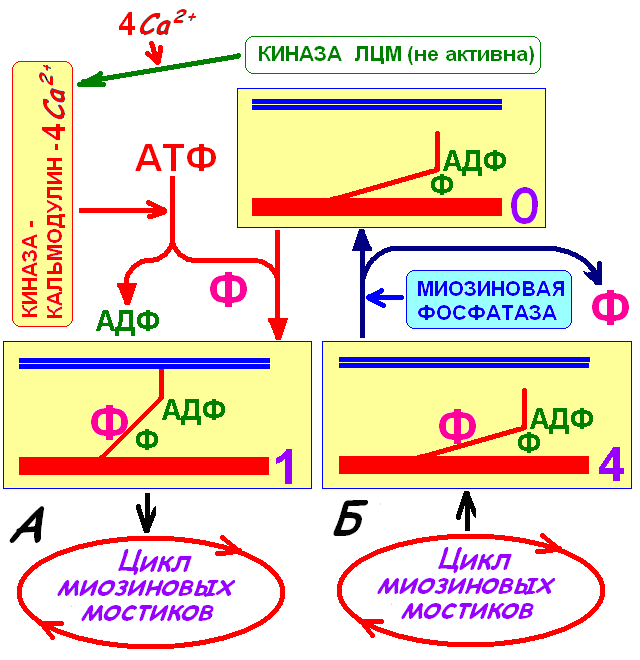
Инициация сокращения в ответ на увеличение концентрации Ca2+ происходит в результате связывания четырёх ионов Ca2+ с кальмодулином. Кальций-кальмодулиновый комплекс активирует киназу лёгких цепей миозина, которая фосфорилирует лёгкие цепи на головках миозина. Головках миозина приобретает аффинитет к актну при условии нахождения на ней АДФ и фосфата (продуктов гидролиза АТФ). Начинается цикл миозиновых мостиков. 51. Расслабление. Энергетика мышечного сокращенияРасслабление скелетного миоцитаПовышенная концентрация Ca2+ в миофибриллярном пространстве сохраняется всего несколько миллисекунд, а затем они «перекачиваются» обратно в цистерны саркоплазматического ретикулума с помощью специального так называемого «кальциевого насоса». Поскольку перекачивание идет против диффузионного градиента, нужна затрата определенной энергии. Ее источником служит расщепление АТФ: на возврат 2 Са2+ в цистерны расходуется 1 АТФ. Ca2+‑АТФаза саркоплазматического ретикулума закачивает Ca2+ из саркоплазмы в цистерны ретикулума, где Ca2+ связывается с кальсеквестрином. При низкой саркоплазматической концентрации Ca2+ тропомиозин закрывает миозинсвязывающие участки и препятствует их взаимодействию с миозином После смерти, когда содержание АТФ в мышечных волокнах снижается вследствие прекращения её синтеза, головки миозина оказываются устойчиво прикреплёнными к тонкой нити. Это состояние трупного окоченения (rigormortis) продолжается, пока не наступит аутолиз, после чего мышцы можно растянуть. У сердечного и гладкого миоцитов Ca2+‑АТФаза саркоплазматического ретикулума закачивает Ca2+ из саркоплазмы в цистерны ретикулума и выкачивает из цитозоля в межклеточное пространство. У гладкого миоцита миозиновые головки дефосфорилируются фосфатазой легких цепей миозина и головки миозина теряют аффинитет к актину. |