Б1 Понятие биомембрана
 Скачать 2.06 Mb. Скачать 2.06 Mb.
|

47. Классификация скелетных мышцКритерии классификации
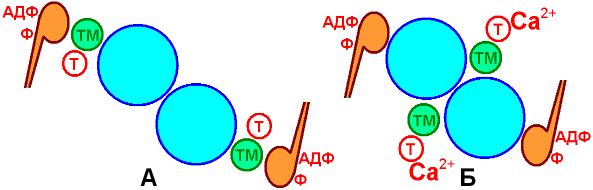
Экстра- и интрафузальныеИнтрафузальные мышечные волокна + чувствительные нервные окончания = мышечные веретёна.– это рецептор, формирует и передает в ЦНС информацию о состоянии скелетной мышцы. Экстрафузальные мышечные волокна образуют основную массу мышцы и выполняют всю работу, необходимую для движения и поддержания позы. Фазные и тонические мышечные волокнаЭкстрафузальные фазные, осуществляющие энергичные и быстрые сокращения. тонические, специализирующиеся на поддержании статического напряжения, или тонуса. Быстрые и медленные мышечные волокнаСкорость сокращения определяется типом миозина. Различают миозин с высокой АТФазной активностью (быстрый) 2й тип волокон и низкой (медленный) 1 тип. Оксидативные и гликолитические мышечные волокнаИсточник энергии для мышечного сокращения АТФ. Два основных пути образования АТФ - окислительный и гликолитический. Окислительные небольшого диаметра, много капилляров, содержа много митохондрий Гликолитические большего диаметра, мало капилляров, мало митохондрий Оксидативные много миоглобина и мало гликогена, красный цвет Гликолитические много гликогена и мало миоглобина. Сводная классификация мышечных волоконРазличают 2 типа скелетных мышечных волокон: I типа (медленные) и II типа (быстрые). Волокна I типа – медленныеоксидативные (красные). Волокна II типа делятся на 2 подтипа – быстрые оксидативные (тип IIа) и быстрые гликолитические (тип IIb). Волокна I типа относят к неутомляемым, IIа – малоутомляемым, IIб – быстроутомляемым. 48. Структурно-функциональная организация скелетной мышцы. Механизм мышечного сокращения и расслабленияДвигательная единицаМышечные волокна не являются функциональной единицей скелетной мускулатуры. Эту роль выполняет нейромоторная, или двигательная единица которая включает мотонейрон и группу мышечных волокон, иннервируемых разветвлениями аксона этого мотонейрона, расположенного в ЦНС В мышцах, обеспечивающих наиболее точные и быстрые движения, двигательная единица состоит из нескольких мышечных волокон, в то время как в мышцах, участвующих в поддержании позы, двигательные единицы включают несколько сотен и даже тысяч мышечных волокон. Все мышечные волокна каждой двигательные единицы относятся к одному типу. Поэтому ДЕ можно относить к типам I, IIa, IIb В мышце волокна разных ДЕ расположены вперемежку. Композиция скелетных мышцКаждая мышца уникальна по спектру входящих в её состав типов мышечных волокон. В зависимости от преобладания в мышцах конкретного типа мышечных волокон скелетные мышцы относят к «красным» и «белым» либо «быстрым» и «медленным». Этот спектр генетически детерминирован Механизм мышечного сокращения и расслабленияМодель скользящих нитейХ.Хаксли и Дж.Хансона (1954 г.). при сокращении длина А-диска не меняется, а I-диск и H‑полоска уменьшаются и даже исчезают Отсюда был сделан вывод, что тонкие и толстые нити при сокращении не изменяют свою длину, а скользят относительно друг друга
49. Электромеханическое сопряжение
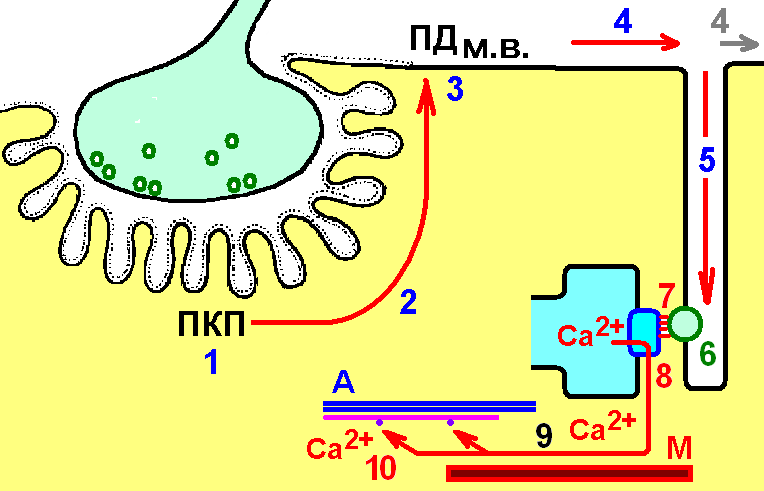
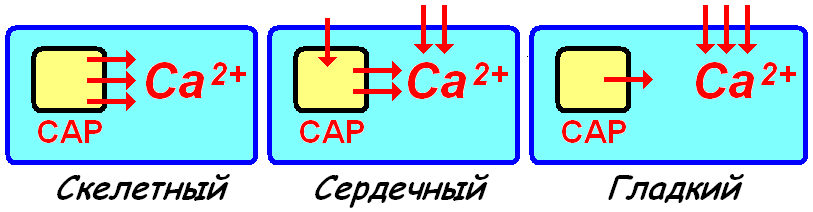
при сокращении скелетного миоцита-процесс преобразования электрического сигнала в химический, а затем в мышечное сокращение. Расположение структур, обеспечивающих передачу электрического сигнала сокращение в мышечном волокне, строго упорядочены К регулярно расположенным Т‑трубочкам с двух сторон подходят терминальные цистерны саркоплазматического ретикулума (САР). Т‑трубочка и расположенные с двух сторон от неё цистерны образуют триаду. Z‑мембраны и зоны перекрытия актиновых и миозиновых нитей расположены рядом с триадами.
Особенности электромеханического сопряжение при сокращении сердечного миоцита
Рианодиновые рецепторы, расположенные на мембране саркоплазматического ретикулума, активируются входящим из межклеточного вещества Ca2+. Кофеин также оказывает на эти рецепторы активирующее действие Особенности электромеханического сопряжение при сокращении гладкого миоцитаУ гладких миоцитов основное колличество Ca2+ в цитозроль поступает из интерстиция Каналы Ca2+ на мембране плохо выраженного саркоплазматического ретикулума управляются рецепторами инозитолтрифосфата (ИТФ). При возбуждении плазматической мембраны активируется фосфолипаза С и образуется ИТФ. Этот механизм функционирует в лимфоцитах, овоцитах. |