Гилберт С. Биология развития. Т.1.doc , БИР. Библиография Гилберт С. Биология развития в 3х т. Т. I Пер с англ. М. Мир, 1993. 228 с
 Скачать 10.9 Mb. Скачать 10.9 Mb.
|

Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.92 ГЛАВА 3
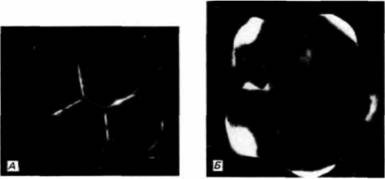
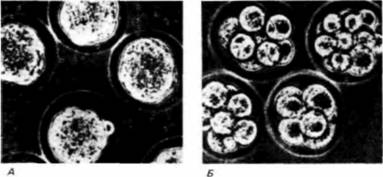
званием поляризации. Разные компоненты клеточной поверхности мигрируют в разные области клетки (Ziomck. Johnson. 1980). Это можно обнаружить, пометив определенные молекулы на клеточной поверхности флуоресцентными красителями. Мечение одним таким красителем, узнающим класс гликопротеинов, показывает, что на 4-клеточной стадии эти гликопротеины беспорядочно распределены по всей плазматической мембране (рис. 3.27. А). Однако на стадии незавершенного третьего деления меченые молекулы обнаруживаются преимущественно на полюсах бластомеров. наиболее удаленных от центра агрегата (рис. 3.27, Б). Это перемещение является, по-видимому, результатом межклеточных взаимодействий, так как оно наблюдается при выделении из зародыша двух связанных между собой бластомеров, но не бластомеров, изолированных поодиночке. Во-вторых, в процессе компактизации участвуют специфические белки клеточной поверхности. Клетки опухоли, называемой тератокарциномой, сходны с клетками внутренней клеточной массы. При инкубации 8-клеточных мышиных зародышей в антисыворотке к стволовым клеткам тератокарциномы компактизация подавляется (рис. 3.28) (Kemlerei al.. 1977; Johnson cl al.. 1979). Таким образом, в клетках тератокарциномы и в бластомерах 8-клеточного зародыша имеется молекула (или молекулы), которая необходима для того, чтобы произошла компактизация. Одной такой молекулой является увоморулин — гликопротеин с молекулярной массой 120 000 дальтон, который обнаруживается на плазматической мембране клеток этих двух типов. Антитела к молекуле увоморулина вызывают декомпактизацию морулы и подавляют прикрепление клеток тератокарциномы друг к другу (Hyafil el al., 1981; Рeуneras et al.. 1983). При осуществлении функции этого гликопротеина важную роль играет углеводная часть молекулы, поскольку туникамицин (вещество, подавляющее гликозилирование белков) также предотвращает компактизацию. В-третьих, плазматическая мембрана при компактизации может также изменяться вследствие реорганизации цитоскелета. На соседних клеточных поверхностях в результате образования актиновых микрофиламентов возникают микроворсинки, прикрепляющие клетки друг к другу. Именно на микроворсинках, по-видимому, функционирует увоморулин как медиатор межклеточной адгезии. Уплощение поверхностей соседних бластомеров в месте их контакта может вызываться укорочением микроворсинок путем деполимеризации актина (Pratt et al., 1982; Sutherland, Calarco-Gillam, 1983). Таким образом, представление, что причиной компактизации являются изменения архитектоники поверхности бластомеров, получает все больше подтверждений.
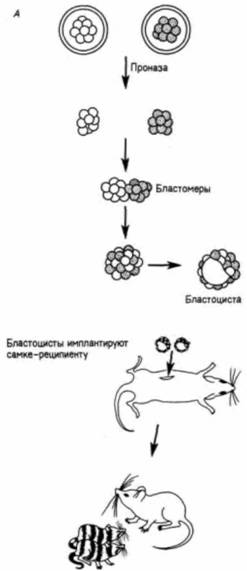
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.ДРОБЛЕНИЕ СОЗДАНИЕ МНОГОКЛЕТОЧНОСТИ 93 Образование внутренней клеточной массыВозникновение внутренней клеточной массы, отличающейся от трофобласта. является важнейшим событием в раннем развитии млекопитающих. Каким образом развитие клетки направляется по тому или другому из этих двух путей? Каким образом клетка получает информацию о том. что ома должна участвовать в образовании организма взрослого млекопитающего или вспомогательной ткани, которая отбрасывается при рождении? Наблюдения, проведенные на живых зародышах, позволяют предположить, что это важное решение является просто результатом того, что в определенное время клетка оказывается в соответствующем месте. До завершения 8-клеточной стадии разные бластомеры явно не отличаются друг от друга по биохимии, морфологии или потенциям. Тем не менее компактизация приводит к образованию внутренних и наружных клеток, глубоко различающихся по своим свойствам. Путем маркировки разных бластомеров в многочисленных исследованиях было показано, что клетки, оказавшиеся снаружи, формируют трофобласт, тогда как клетки, оказавшиеся внутри, образуют зародыш (Tarkowski. Wroblewska, 1967)1. Если любой бластомер 4-клеточного зародыша мыши поместить на наружную поверхность рыхлой массы бластомеров другого зародыша, то из пересаженной клетки разовьется только ткань трофобласта (Hillman et al., 1972). Следовательно, будет ли клетка участвовать в образовании трофобласта или зародыша, определяется, по-видимому, тем, где оказалась клетка после компактизации — на поверхности или внутри зародыша. Если большинство клеток бластоцисты дает начало трофобласту, то сколько же клеток фактически образует зародыш? Один из путей для получения ответа на этот вопрос заключается в создании аллофенных мышей. Аллофенные мыши являются результатом развития химерных зародышей, полученных путем слияния двух зародышей на ранних стадиях дробления (обычно 4- или 8-клеточных). Как показано на рис. 3.29, у двух генетически различаю- 1 Установлено, что внутренние клетки чаще всего происходят от того бластомера, который на 2-клеточной стадии делится первым. Потомки этого бластомера обычно первыми достигают 8-клеточной стадии и делятся таким образом, что оказываются внутри рыхлого скопления бластомеров (Graham, Kelly, 1977).
|