Гилберт С. Биология развития. Т.1.doc , БИР. Библиография Гилберт С. Биология развития в 3х т. Т. I Пер с англ. М. Мир, 1993. 228 с
 Скачать 10.9 Mb. Скачать 10.9 Mb.
|

Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.86 ГЛАВА 3
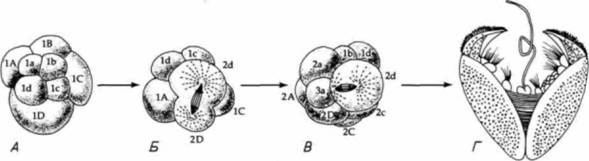
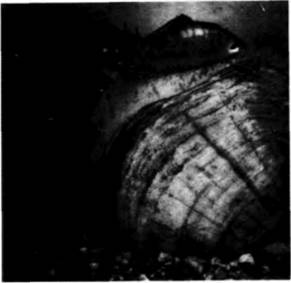
фикация морфогенеза позволила особи выжить в условиях, которые, не произойди такое изменение, были бы для нее крайне неблагоприятными. Одна из модификаций такого рода, открытая Фрэнком Лилли (Lillie, 1898), заключается в изменении типичного хода спирального дробления в семействе двустворчатыx моллюсков Unionidae. В отличие от большинства двустворчатых моллюсков Unio и родственные формы живут в реках с быстрым течением. Такие условия обитания обычно создают проблему расселения личинок: поскольку взрослые особи ведут оседлый образ жизни, свободноплавающие личинки должны были бы всегда сноситься течением вниз. Эти моллюски, однако, решили проблему расселения, осуществив два изменения в своем развитии. Первое из них касается дробления зародыша. При типичном для моллюсков дроблении либо все макромеры имеют одинаковые размеры, либо бластомер 2D является самой крупной клеткой на стадии 16 бластомеров. Однако в процессе дробления яиц Unioпри переходе к этой стадии наибольшее количество цитоплазмы получает бластомер 2d (рис. 3.17). Эта клетка делится, и ее потомки образуюn большую часть структур личинки, в том числе железу, способную образовать массивную раковину. Такая личинка (называемая глохидием) напоминает медвежий капкан в миниатюре. Глохидии имеют чувствительные волоски, которые вызывают захлопывание створок раковины, как только они прикоснутся к жабрам или плавникам проплывающей мимо рыбы. Глохидии занимаются «хич-хайкингом» (от англ. hitchhike путешествовать бесплатно на попутных машинах) с помощью рыбы, пока не достигнут стадии, на которой они готовы отделиться и метаморфизировать во взрослых моллюсков. Таким способом они могут расселяться вверх по течению. У некоторых видов глохидии высвобождаются из выводковой камеры самки и просто ожидают, чтобы рыба подплыла близко к ним. Некоторые другие виды, как, например, Lampsilisventricola, повысили вероятность встречи личинок с рыбой, осуществив еще одну модификацию развития (Welsh, 1969). У многих моллюсков развивается тонкая мантия, которая колышется около раковины и окружает выводковую камеру (marsupium). У некоторых унионид форма выводковой камеры и волнообразные движения мантии создают впечатление плывущей рыбы. Чтобы сделать иллюзию более полной, у них развивается черное «пятно-глаз» на одном конце и выступающий «хвост» на другом. «Рыба», которую мы видим на рис. 3.18. на самом деле не рыба, а выводковая
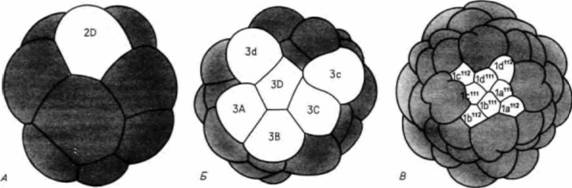
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с._______________ ДРОБЛЕНИЕ: СОЗДАНИЕ МНОГОКЛЕТОЧНОСТИ 87 камера и мантия моллюска, находящегося внизу. Когда соблазненная видом «добычи» хищная рыба подплывает достаточно близко, моллюск выбрасывает глохидии из выводковой камеры. Изменив таким образом ход развития, моллюски из семейства Unionidae сумели выжить в условиях, которые без таких модификаций были бы для них неблагоприятными. При изучении дробления у моллюсков было сделано еще одно интереснейшее открытие, а именно что некоторые бластомеры сообщаются между собой. У моллюсков, имеющих на 4-клеточной стадии бластомеры одинаковых размеров, определение того, какой из них даст начало клетке-предшественнице мезодермы, происходит между пятым и шестым делениями. В это время микромер 3D образует направленное внутрь выпячивание, которое приходит в контакт с микромерами анимального полюса. Без такого контакта бластомер 4d, отделяемый макромером 3D, не образует мезодермы (Biggelaar. Guerrier. 1979) 1. Де Лаат и др. (de Laat et al.. 1980) показали, что в то время, когда осуществляется контакт (но не раньше), между макромером 3D и центральными микромерами может происходить обмен молекулами небольших размеров. Эти исследователи инъецировали в один из макромеров краситель люцифер желтый (молекулярная масса 457). При введении красителя до 32-клеточной стадии он остается в этом единственном макромере и его потомках. Однако при инъекции красителя в бластомер 3D в начале 32-клеточной стадии в желтый цвет окрашивается первый квартет микромеров (рис. 3.19). Низкомолекулярное вещество передается от одной клетки к другой именно в то время, когда микромеры изменяют проспективные потенции бластомера 3D. Исследование с помощью просвечивающего электронного микроскопа показывает, что в это время на поверхности клеток появляются щелевые контакты. Такие изменения плазматической мембраны облегчают межклеточный обмен небольшими молекулами и ионами, и они встречаются во многих тканях как зародышей, так и взрослых организмов. (Ультраструктура этих контактов будет рассмотрена в гл. 15.) Билатеральное голобластическое дроблениеБилатеральное голобластическое дробление встречается главным образом у асцидий (оболочники). На рис. 3.20 представлены стадии дробления у асцидий Slyelapartita. Наиболее замечательная особенность этого типа дробления заключается в том, что плоскость первого деления устанавливает единственную плоскость симметрии зародыша. Каждое последующее деление ориентируется по отношению к этой плоскости симметрии, так что половина зародыша по одну сторону от первой борозды представляет собой зеркальное отражение половины зародыша по другую ее сторону. Второе деление так же, как и первое, меридиональное, но в отличие от
1 Не думайте, что мы забыли о зародышах моллюсков, у которых на 4-клеточной стадии бластомеры различаются по размерам. Им будет уделено достаточно внимания в гл. 7. |