Гилберт С. Биология развития. Т.1.doc , БИР. Библиография Гилберт С. Биология развития в 3х т. Т. I Пер с англ. М. Мир, 1993. 228 с
 Скачать 10.9 Mb. Скачать 10.9 Mb.
|

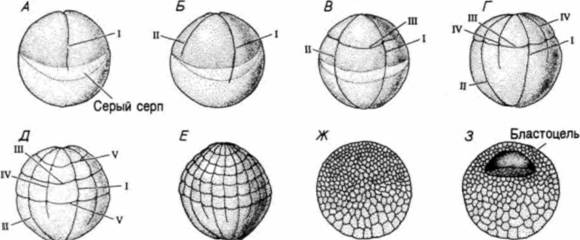
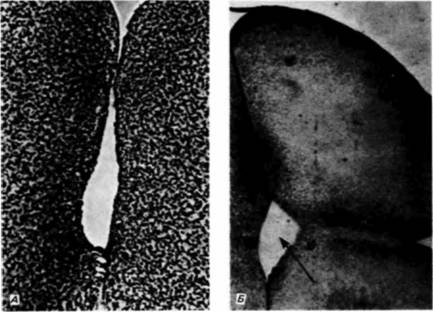
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.ДРОБЛЕНИЕ: СОЗДАНИЕ МНОГОКЛЕТОЧНОСТИ 81 этих клеток никогда не бывает направлено в сторону бластоцеля: такая ориентация клеточной популяции в одной плоскости способствует ее дальнейшему распространению вширь Согласно другому предположению (Wolpert, Gustafson, 1961; Wolpert, Mercer, 1963), давление жидкости бластоцеля не является необходимым условием получения наблюдаемого эффекта. Авторы этой гипотезы придают особое значение разной адгезивности клеток по отношению друг к другу и к гиалиновому слою. Предполагается, что, пока клетки остаются прочно связанными с гиалиновым слоем, единственной возможностью для них остается расширение поверхности клеточного пласта. Формирование бластулы является скорее следствием этой экспансии, чем какого-либо другого процесса. На наружной поверхности клеток бластулы образуются реснички (рис. 3.9). что позволяет бластуле вращаться внутри оболочки оплодотворения. Вскоре клетки секретируют фермент вылупления, который переваривает оболочку. Зародыш вылупляется и становится свободноплавающей бластулой. АмфибииДробление у зародышей большинства бесхвостых и хвостатых амфибий радиально-симметричное и голобластическое, как у иглокожих. Однако яйцо амфибий содержит гораздо больше желтка. Этот желток, сосредоточенный в вегетативном полушарии, затрудняет дробление. Так, первое деление начинается на анимальном полюсе и медленно распространяется вниз на вегетативную область (рис. 3.10). У хвостатой амфибии аксолотля борозда дробления распространяется в анимальной области со скоростью, близкой к 1 мм/мин. Ниже области серого серпа скорость ее распространения снижается и при приближении к вегетативному полюсу достигает 0.02— 0.03 мм/мин (Нага. 1977). На рис. 3.11, А представлена микрофотография яйца лягушки на стадии первого деления дробления, полученная с помощью сканирующего электронного микроскопа. Можно видеть складки в борозде дробления и различия между бороздами в анимальном и вегетативном полушариях. В то время, когда борозда первого деления дробления еще не завершила разделения богатой желтком цитоплазмы вегетативного полушария, борозды второго деления уже закладываются вблизи от анимального полюса. Борозды этого деления располагаются под прямыми углами к первой борозде и ложатся, так же, как и она, меридионально (рис. 3.11. Б). Третье деление, как это можно было ожидать, бывает экваториальным. Однако из-за большой концентрации желтка в вегетативной области борозды третьего деления дробления располагаются значительно ближе к анимальному полюсу. Они разделяют зародыш лягушки на четыре анимальных бластомера меньшего размера и четыре более крупных бластомера в вегетативной области (рис. 3.11, В). В результате этого неравного голобластического деления возникают область быстро делящихся бластомеров вблизи анимального полюса и область более медленно делящихся бластомеров. На последующих стадиях
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.82 ГЛАВА 3
дробления анимальная область содержит многочисленные мелкие клетки, тогда как в вегетативной области имеется лишь относительно небольшое число крупных богатых желтком бластомеров (рис. 3.10). Зародыш амфибий, содержащий от 16 до 64 клеток, обычно называют морулой (от латинского названия тутовой ягоды, на которую он отдаленно похож). На 128-клеточной стадии появляется хорошо различимый бластоцель, и принято считать, что в это время зародыш достиг стадии бластулы. На самом деле образование бластоцеля прослеживается с самого первого деления дробления. Показано (Kalt. 1971), что у Xenopuslaevisпервая борозда дробления в анимальном полушарии образует расширение, причем между бластомерами возникает маленькая полость, изолированная от наружной среды плотными межклеточными контактами (рис. 3.12). Эта полость при последующих делениях расширяется и преобразуется в бластоцель. Бластоцель, вероятно, выполняет две основные функции. Во-первых, он представляет собой полость, которая даст возможность клеткам при гаструляции мигрировать внутрь зародыша. Во-вторых, существование бластоцеля предотвращает преждевременное взаимодействие между клетками, которые находятся выше и ниже его. Когда Ньюкуп (Nieuwkoop, 1973) брал у зародыша тритона клетки из крыши бластоцеля и помещал их вблизи богатых желтком вегетативных клеток дна бластоцеля, то из
|