Гилберт С. Биология развития. Т.1.doc , БИР. Библиография Гилберт С. Биология развития в 3х т. Т. I Пер с англ. М. Мир, 1993. 228 с
 Скачать 10.9 Mb. Скачать 10.9 Mb.
|

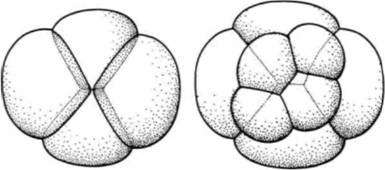
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.______ ДРОБЛЕНИЕ: СОЗДАНИЕ МНОГОКЛЕТОЧНОСТИ 83 этих анимальных клеток развивались не производные эктодермы, а мезодермальная ткань. Поскольку в норме мезодерму образуют анимальные клетки, которые прилежат к предшественникам энтодермы, представляется вероятным, что вегетативные бластомеры оказывают влияние на прилежащие клетки, побуждая их дифференцироваться в мезодермальные ткани (см. гл. 4). Таким образом, наличие бластоцеля обеспечивает сохранение клетками, предназначенными для образования покровного эпителия и нервной системы, своих исходных свойств. Спиральное голобластическое дроблениеСпиральное дробление характерно для нескольких групп червей (кольчатых, турбеллярий, немертин) и всех моллюсков, за исключением головоногих. Оно во многих отношениях отличается от радиального дробления. Во-первых, яйца не делятся параллельно или перпендикулярно анимально-вегетативной оси. Плоскости делений дробления ориентированы наклонно, что приводит к «спиральному» расположению дочерних бластомеров. Во-вторых, число контактов между клетками больше, чем при радиальном дроблении. Фактически они принимают расположение, наиболее стабильное в термодинамическом отношении, подобное расположению контактирующих мыльных пузырьков (рис. 3.13). В-третьих, зародыши со спиральным типом дробления проходят меньше делений до начала гаструляции. Это позволило проследить судьбу каждой отдельной клетки бластулы. Когда сопоставили судьбы отдельных клеток зародышей кольчатых червей, плоских червей и моллюсков, то оказалось. что у клеток, занимающих одно и то же положение, судьба в принципе одинакова (Wilson. 1898). Возникающие таким образом бластулы не имеют бластоцеля и носят название стерробластул. На рис. 3.14 и 3.15 показано дробление зародыша моллюска. Первые два деления почти меридиональны и приводят к образованию четырех крупных макромеров (обозначенных А, В, С и D). У многих видов эти бластомеры различаются по своим размерам (причем D бывает самым крупным) это позволяет идентифицировать отдельные бластомеры. При последующих делениях каждый макромер на своем анимальном полюсе отделяет маленький микромер. Каждый последовательно появляющийся квартет микромеров смещается вправо или влево относительно сестринских макромеров, что приводит к характерному для данного типа дробления расположению бластомеров по спирали. Если смотреть на зародыш со стороны анимального полюса, то можно видеть, что верхние концы митотических веретен бывают наклонены поочередно то по часовой стрелке, то против нее. В результате микромеры оказываются смещенными по отношению к отделяющим их макромерам то влево, то вправо. При третьем делении макромер А дает две дочерние клетки макромер 1Α и микромер 1a. Бластомеры В, С и D ведут себя сходным образом, формируя первый квартет микромеров. Если на этой стадии смотреть на дробящиеся яйца со стороны анимального полюса, то у большинства видов микромеры оказываются смещенными вправо по отношению к отделившим их макромерам (дексиотропная спираль) в противоположность леотропной спирали при их смещении влево. При четвертом делении макромер 1А делится на макромер 2А и микромер 2а, а микромер 1a на два микромера, 1a1 и 1a2. При последующем делении макромер 2А дает бластомеры ЗА и За, a такие микромеры, как 1a2, дает клетки 1a21 и 1a22. Смещение плоскости дробления влево или вправо контролируется цитоплазматическими факторами. Это было установлено путем изучения мутаций закручивания раковин у улиток. У некоторых улиток устье, которым открывается спираль раковины, находится с правой стороны от ее завитка, тогда как у других устье располагается слева. Обычно направление закручивания бывает одинаковым у всех особей данного вида. Однако иногда встречаются мутанты. Например, среди представителей видов с устьем, расположенным справа, встречаются особи, у которых оно расположено слева. Результаты
|