Биологические основы селекции зерновых колосовых культур 06. 01. 05 Селекция и семеноводство
 Скачать 0.88 Mb. Скачать 0.88 Mb.
|

|
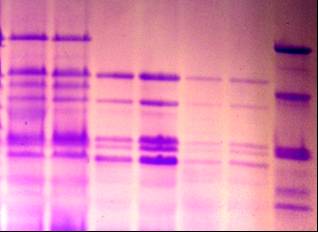
УДК 631.527:633:1 На правах рукописи БУЛАТОВА КУЛЬПАШ МАНСУРОВНА Биологические основы селекции зерновых колосовых культур 06.01.05 – Селекция и семеноводство Автореферат диссертации на соискание ученой степени доктора биологических наук Республика Казахстан Алматы, 2009 Работа выполнена в ТОО «Казахский научно-исследовательский институт земледелия и растениеводства» АО «КазАгроИнновация» Научные консультанты: доктор биологических наук, профессор, академик НАН РК, УААН и РАСХН Уразалиев Р.А., доктор биологических наук, профессор Перуанский Ю.В. Официальные оппоненты: доктор биологических наук, профессор, академик НАН РК, Шаханов Е.Ш., доктор биологических наук, профессор Сеилова Л.М., доктор биологических наук, профессор, академик РАЕН, Седловский А.И. Ведущая организация: Казахский национальный аграрный университет Защита состоится 5 июня 2009 года в 14.00 часов на заседании диссертационного совета ОД 55.05.01 при Казахском НИИ земледелия и растениеводства по адресу: 040909, Республика Казахстан, Алматинская область, Карасайский район, п. Алмалыбак, ул. Ерлепесова 1. С диссертацией можно ознакомиться в библиотеке Казахского НИИ земледелия и растениеводства. Автореферат разослан ____ ____________________ 2009 г. Ученый секретарь диссертационного совета, доктор с-х. наук Альдеков Н.А. ВВЕДЕНИЕ Актуальность проблемы Пшеница и ячмень являются основными зерновыми колосовыми культурами, возделываемыми в Казахстане. Посевные площади этих культур составляют в Республике более 15 млн. га. Одним из главных факторов увеличения валового сбора зерна является селекционный путь - создание высокоурожайных сортов с комплексом ценных хозяйственно-биологических признаков и свойств. Успех в решении этой важнейшей для сельскохозяйственной отрасли Республики задачи зависит от уровня генетического разнообразия исходных коллекций, гибридных популяций, методологической базы отбора желаемых генотипов в ходе селекционного процесса. Значимость биологических методов анализа для решения проблем генофонда, селекции и семеноводства сельскохозяйственных культур бесспорна, однако их практическое применение все еще не на должном уровне. Разработка, совершенствование и внедрение надежных и эффективных методов биологического анализа в изучение генетических ресурсов, процесс отбора ценных форм, сохранение биологических свойств сорта в ходе семеноводства являются актуальными задачами для отечественной селекции и необходимыми условиями повышения продовольственной безопасности страны. В национальном генофонде Республики сосредоточено более 30 тыс. образцов пшеницы, 3 тысяч коллекционных номеров ячменя, представленных такими категориями, как: коммерческие и местные сорта, дикие сородичи, признаковые коллекции. Генетическое разнообразие коллекционного материала не изучено, следовательно, не использовано в достаточной мере. В то же время, образцы с редкими и новыми аллелями признаков могут дать начало селекции высокоурожайных, устойчивых к неблагоприятным условиям среды, ценных по другим хозяйственно-ценным показателям сортов. Выявление таких форм и их использование в селекции сопряжено с изучением большого числа коллекционных образцов, целенаправленного подбора компонентов для скрещиваний, поэтапной оценки селекционных линий. Успешная реализация этих задач невозможна без использования достижений генетики, молекулярной биологии, физиологии, и других наук в биологическом анализе растений на этапах селекционного процесса зерновых колосовых культур. Особую значимость в их числе приобретают методы, разработанные на основе специфичности состава запасных белков, ферментов, накоплении определенных метаболитов на разных стадиях, условиях развития генотипа и сопряженности их характеристик с проявлением хозяйственно-ценных признаков, что актуально для селекции сельскохозяйственных культур. Работа была выполнена в рамках государственных программ: 1991-1995гг. «Разработать новые и усовершенствовать существующие схемы, методы селекции и на их основе создать новые высокопродуктивные, устойчивые к болезням, вредителям, неблагоприятным факторам среды и с высоким качеством продукции сорта и гибриды пшеницы, ячменя, овса, кукурузы, сорго, сои, гороха с разработкой экологических основ промышленного семеноводства». № гос.регистрациии 0194РК00028; 1996-2000гг. «Создать новые с использованием методов генетики, биотехнологии и других биологических дисциплин устойчивые к стрессовым факторам среды, высокопродуктивные сорта и гибриды зерновых, зернобобовых, сорговых культур». № гос.регистрации 0196РК00292; 2001-2005гг. «Сбор, интродукция, комплексное изучение и выявление доноров с желательными хозяйственно-ценными признаками, организация, поддержание существующих коллекций зерновых, зернобобовых, крупяных, технических и лекарственных культур на Юге и Юго-Востоке Казахстана», № гос.регистрации 0101РК00226; 2006-2008гг. «Разработка биохимических подходов для дифференциации генотипов пшеницы по устойчивости к желтой ржавчине», № гос.регистрации 0106РК00730. Цель работы – Разработать и усовершенствовать методические основы биологического анализа растений в селекции пшеницы и ячменя. Для достижения цели необходимо было решить следующие задачи: -определить вариабельность состава запасных белков пшеницы, установить генетический контроль новых субъединиц белков, усовершенствовать систему регистрации коллекционных и селекционных образцов, мутантных линий, сортов по генетико-биохимическим параметрам, провести регистрацию генетических ресурсов и выявить ценные для селекции генотипы; -разработать систему регистрации и идентификации линий, биотипов и сортов, оценить разнообразие генетических ресурсов ячменя по полиморфности белок-кодирующих локусов; -выявить биологические особенности жаро-засухоустойчивых и скороспелых форм ячменя, разработать на этой основе методы отбора желаемых генотипов в коллекционных и селекционных питомниках; - выделить коллекционные образцы – источники и доноры хозяйственно-ценных признаков для селекции пшеницы и ячменя; на основе разработанных методов и приемов биологического анализа, отобрать перспективные селекционные линии. Научная новизна Впервые для идентификации коллекционного и селекционного материала пшеницы и ячменя предлагается комплекс биологических тестов, включающих: - уточненную номенклатуру высокомолекулярных субъединиц глютенина; - сорта – дифференциаторы для идентификации, отбора и паспортизации коллекционных и селекционных линий пшеницы; - совершенствованную систему регистрации образцов ячменя по гордеинам с генетико-биохимической характеристикой их компонентов. Проведена генетико-биохимическая идентификация по белковым формулам 1831 образца генофонда зерновых колосовых культур, установлено генетическое разнообразие коллекций, выделены источники редких и ценных для селекции генов. Предложен комплекс биологических тестов для отбора скороспелых и засухоустойчивых форм ячменя в поэтапной оценке в селекционных питомниках. Предложена схема биологических тестов в селекции пшеницы и ячменя по основным хозяйственно-ценным признакам. Практическая ценность работы Проведена идентификация и регистрация генофонда пшеницы и ячменя по блокам компонентов запасных белков зерна, результаты опубликованы в 2-ух каталогах . На основе спектров запасных белков ведется паспортизация сортов пшеницы и ячменя, передаваемых на Государственное сортоиспытание; С участием соискателя переданы на Государственное сортоиспытание 2 сорта озимой мягкой и 1 сорт яровой твердой пшеницы (от 5 до 10% участия в авторстве); Созданы константные линии озимой пшеницы с эффективными генами устойчивости к желтой ржавчине, 3 из которых находятся в питомнике конкурсного сортоиспытания; Для селекции на засухоустойчивость и скороспелость создано 233 гибридных и 34 перспективных по комплексу ценных признаков и свойств линий гордеиновых биотипов ячменя, часть из которых испытывается в питомниках конкурсного сортоиспытания. Линия 61/7 показала значительное превышение по урожайности над стандартным сортом. Положения, выносимые на защиту: -усовершенствованная система регистрации коллекционных и селекционных образцов пшеницы и ячменя по белковым формулам; -идентифицированные коллекции генофонда ячменя и пшеницы, их генетическое разнообразие , ценные источники хозяйственно-биологических признаков; -комплекс биологических критериев в дифференциации и отборе скороспелых и засухоустойчивых форм ячменя; -перспективные линии и сорта пшеницы и ячменя, как практический результат биологического анализа генофонда и использования ценных источников в селекции по основным хозяйственно-ценным признакам. Апробация работы Результаты исследований были представлены и доложены на 27 научно-практических конференциях, совещаниях и симпозиумах, в т.ч. на межд.сов. «Генбанк растений и его использование в селекции», (Алматы, 1995), межд. науч. пр.конф. «Аграрная наука на рубеже веков» (Акмола, 1997), 2-ой межд. конф. «Проблемы экологии АПК и охрана окружающей среды» (Алматы,1998), межд. науч. конф. «Природные соединения – регуляторы метаболизма и адаптации растений» (Алматы, 1999), межд. науч. конф. «Физиолого-биохимические и генетические основы устойчивости и продук-тивности раст.» ( Алматы, 1999 ), мат. межд. научно-теоретической конф. «Стратегия земледелия и растениеводства на пороге ХYI века.» ( Алма-ты,1999), на 8-м Международном симпозиуме по ячменю (Adelaide, South Australia, 2000), на 4-ой межд. науч. конф. «Проблемы экологии АПК и охрана окружающей среды» (Щучинск, 2002), на международном симпозиуме “Biotechnology Approaches for Exploitation and Preservation of Plant Resources” (Ялта, 2002), межд. науч. конф. «Современное состояние проблем и достижений в области генетики и селекции» (Алматы, 2003), первой центрально-азиатской конференции по пшенице, (Алматы, 2003), 6-ой межд. науч. практ. конф. . «Научное обеспечение устойчивого развития АПК Казахстана, Кыргызстана, Монголии, России, Таджикистана и Узбекистана», (Павлодар, 2003), межд. научн. конф. «Стратегия научного обеспечения АПК РК в отраслях земледелия, растениеводства и садоводства. Реальность и перспективы», (Алматы, 2004), межд. научн. конф. «Достижения аграрной науки в области земледелия, селекции и растениеводства», (Алмалыбак, 2004), межд. конф. «Развитие ключевых направлений сельскохозяйственной науки в Казахстане: селекция, биотехнология, генетические ресурсы», (Астана, 2004), 3-ем Московском Международном конгрессе «Биотехнология: состоя-ние и перспективы развития» (Москва, 2005), Республиканской научно-теоретической конференции «Сейфуллинские чтения – 3», посвященной 50-летию основания КазГАТУ, Астана, 2007, 10-ой международной конференции по научному обеспечению азиатских территорий (Улан-Батор, 2007), на XI межд. научно-практ. конф. «Развитие АПК азиатских территорий»- (Новосибирск.-2008), международной конференции «Современные методы использования генетических ресурсов в селекции ячменя и овса» (С.Петербург, 2009). Публикации: по материалам диссертации опубликовано 76 научных работ (в списке приведено 46), в том числе 2 каталога, подана 1 заявка на получение патента РК (№ гос. регистрации 2007/0593.1). Структура диссертации диссертационная работа изложена на 270 страницах печатного текста, состоит из введения, основной части, включающей обзор литературы, объекты, методы и условия проведения исследований, результаты исследований, а также заключения, списка использованных источников, включающего 533 наименования, содержит 95 таблиц и 39 рисунков, 4 приложения. Автор выражает особую признательность научным консультантам: академику Р.А.Уразалиеву и д-ру биол. наук Перуанскому Ю.В. за ценные советы и предложения при подготовке диссертации. ОСНОВНАЯ ЧАСТЬ 1 Обзор литературы В обзоре дана оценка современного состояния по использованию методов биологического анализа растений в селекции и семеноводстве сельскохозяйственных культур по проблемам: генетические марке-ры в изучении и регистрации генофонда; молекулярные маркеры в решении проблем генофонда, селекции и семеноводства; ферменты как системы белковых марке-ров; запасные белки семян как молекулярные маркеры; методы и приемы диагнос-тики засухоустойчивости растений; биохимические особенности засухоус-тойчивых форм растений; скороспелость как механизм адаптации растений к засухе. 2 Объекты, методы и условия проведения исследований 2.1 Условия проведения исследований Аналитические исследования проводились в лаборатории КазНИИЗиР, полевые исследования велись в агроэкологических зонах Алматинской (полевые стационары КазНИИЗиР), Западно-Казахстанской (полевые стационары Актюбинская СОС), Акмолинской (полевые стационары КазНИИЗХ им.А.И.Бараева), Павлодарской (полевые стационары Павлодарского НИИСХ) областей. Агротехника на селекционных посевах была общепринятой для соответствующих зон. 2.2 Объекты исследований: 811 образцов генофонда Triticum aestivum L., включающих сорта озимого и ярового типа развития, коллекционные номера зарубежной селекции, линии международных питомников; номера селекционных питомников яровой мягкой пшеницы: СП-2 и КП – 307 и 299 номеров, соответственно; регенерантные линии дигаплоида АДГ 147 – производные сорта Саратовская 52, изогенные линии с генами устойчивости к желтой ржавчине; 264 коллекционных образца Triticum durum; 752 образца генофонда ячменя, гибридные линии ячменя F3 – F 6 поколений. 2.3 Методы исследований: Полевые оценки и структурный анализ растений проводили по стандартным методикам, применяемым для характеристики селекционного материала. Гибридизацию проводили «твелл» методом (Borlaug, в модификации Уразалиева, Шегебаева, 1981). Глютенины в составе суммарных белков эндосперма экстрагировали 0,0625 М трис- НСl буфером, рН 6,8, содержащим 5% меркаптоэтанола, 3% додецилсульфата натрия, 10% глицерина, 0,1% красителя бромфенолового синего. Система разделения соответствовала модифицированному методу Лэммли,1970 (Булатова К.М., 1985). Компонентный состав проламинов определяли путем электрофореза экстрагированной 70% этанолом белковой фракции в полиакриламидном геле в кислой системе по прописи Попереля Ф.А. и др. (Попереля Ф.А.. Асыка Ю.А. 1989). Количественное соотно-шение компонентов в белковом спектре определяли путем сканирования электро-фореграмм с последующим расчетом площадей пиков. Засухоустойчивость стан-дартных сортов ячменя определяли по числу проросших семян после прогревания и проращивания в растворе сахарозы (Методические указания ВИР под. ред. Удовенко Т.В. 1982), по уровню свободного пролина в проростках материал оценивался методом Бейтса с сотр. (Bates et.al., 1973). Общая активность пероксидазы замерялась на спектрофотометре СФ-26 и СПЕКОЛ-11 по скорости реакции окисления бензидина (по Бояркину). Перевод активности в нанокаталы проводили по Лебедевой О.В. и др.,1977. Электрофоретическое разделение пероксидаз осуществляли в трис-глициновом буфере при рН 8,3. Изозимы пероксидазы выявляли бензидиновым реагентом в ацентатном буфере при рН 4,7 с последующим переносом в 0,1 % водный раствор перекиси водорода (Кабжанова С.Б., Перуанский Ю.В.1975). Относительную активность индивидуальных изоформ определяли хронометрированием (Liu E.M. 1973). Оценка проростков озимой пшеницы на устойчивость к расам желтой ржавчины в условиях теплицы по методике Gassner, Straib (1929), полевая устойчивость растений к видам ржавчины по шкале Peterson и др. (1948) определялась в лаборатории «иммунитета растений» НИИПББ НЦБ МОН РК. Математическая обработка данных, в т.ч. и кластеризация методом UPGMA осуществлялись при помощи пакета программ Statistica V 5.0. Оценку генетического разнообразия коллекций определяли согласно способа Nei в модификации Hintum & Elings, 1991, цитировано по Morgunov A.I. et.al. 1993. В автореферате имеются следующие сокращения и обозначения: Glu A1, -B1, -D1 локусы – глютенинкодирующие локусы; ВМСГ –высокомолекулярные субъединицы глютенина. НМГ–низкомолекулярный глютенин; Hrd А,-B,–F-гордеинкодирующие локусы; ДДСNaПААГ–полиакриламидный гель с додецил-сульфатом натрия; ОЭП – относительная электрофоретическая подвижность; СИММИТ- Международный центр улучшения пшеницы и кукурузы; ИКАРДА – Международный Центр по сельскохозяйственным исследованиям в засушливых ре-гионах; КАСИП–Казахстанско-Сибирский питомник; WWERYT – Восточно – Европейский питомник сортоиспытания озимой мягкой пшеницы; КСИ- питомник конкурсного сортоиспытания; ГСИ – государственное сортоиспытание. 3 Результаты исследований 3.1 Совершенствование способов идентификации и отбора генотипов пшеницы и ячменя по биохимическим тестам Возросший в последнее время интерес к субъединичному составу глютенинов пшеницы связан с функциональной значимостью конкретных белков в детерминации хлебопекарных свойств. На этой основе рекомендованы градации отдельных высокомолекулярных субъединиц глютенина в соответствии с их вкладом в так называемую оценку по глютенину -Glutenin quality score (Payne et.al., 1987). В то же время, в ходе исследований выявляются редкие и новые варианты субъединиц глютенина пшеницы, изучение и оценка селекционной ценности которых может в значительной степени повысить эффективность маркерной селекции. Кроме того, отмечается некоторое несоот-ветствие прогнозной оценки качества по глютенинам с результатами традиционных методов оценки технологических показателей. Причиной этого может являться различие в аллельном состоянии глютенинкодирующих локусов, не выявляемое при фракционировании продуктов их биосинтеза при определенных условиях. Уточнение генетического контроля и идентификация новых и редких белковых субъединиц, выявляемых в ходе изучения коллекционного и селекционного материала необходимы как для достоверной оценки образцов, так и для расширения генетической основы создаваемых сортов путем изучения ценности генотипов со специфическими вариантами аллелей и включения их в селекционный процесс. 3.1.1 Новые и редкие аллели локусов, контролирующих биосинтез высокомолекулярных субъединиц глютенина пшеницы В электрофоретическом спектре запасных белков мягкой пшеницы (геном АА ВВ DD), при фрак-ционировании белковых проб стандартным методом Laemmli, как правило, в зоне ВМСГ, контролируемых локусом Glu B1, проявляются 2 компонента. Разделение белков в модифицированных нами условиях выявляет еще один компонент (рисунок 1, дорожка №2, отмечен белой точкой), которого нет в спектре ВМСГ твердой пшеницы (дорожка №1). Аналогичный вариант субъединицы (а) характерен для многих сортов озимой пшеницы Казахстанской селекции, в том числе и Богарной 56 (дорожка №8), для известного сорта Безостая 1. Дополнительный компонент (b) проявляется и в спектре яровой мягкой пшеницы Саратовская 52, Саратовская 29 и многих других сортов мягкой пшеницы селекции НИИСХ Юго-Востока РФ (дорожка 4, черная точка), с той лишь разницей, что он имеет меньшую электро-форетическую подвижность. Отсутствие сопутствующей субъединицы в спектре ВМСГ твёрдой пшеницы, не имеющей D геном, ее наличие у дигаплоидной линии с мутацией, приведшей к исчезновению Вх7 субъединицы, обнаружение в спектре ВМСГ дикого сородича Aegilops cylindrica (дорожка №7, показана стрелкой) аналогичной субъединицы свидетельствуют о том, что ее контроль осущест-вляется D геномом. Выявлено 4 варианта сопутствующих субъединиц, контролируемых 1D хромосомой: a, b, c, d. (носители Безостая 1, Саратовская 29, Бабило, Сурхак Ишкашимский, соответственно). Для более точной оценки генетического контроля сопутствующих субъединиц были использованы нуллисомные линии сорта Чайниз Спринг (nullisome 1D tetrasome 1B – дорожки № 3,4 на рисунке 2, nullisome 1D tetrasome 1A –дорожки №5,6, nullisome 1B tetrasome 1D – дорожка №7). Электрофорез запасных белков в модифицированных условиях показал, что биосинтез высокомолекулярной субъединицы, сопутствующей субъединице глютенина 7 в центральной части зоны ВМСГ (показано стрелкой) контролируется 1D хромосомой. Анализ частоты встречаемости субъединиц 7 в сочетании с обнаруженной «а» субъединицей в составе глютенинов мягкой пшеницы из разных селекционных центров показал, что доля образцов, имеющих в составе высокомолекулярных глютенинов субъединицы 7 и «а» наиболее значительна в наборе районированных и проходящих испытание сортов Казахстана (90,5%), среди сортов, созданных в селекционно-генетическом институте – национальном центре семеноведения и сортоизучения УААН (95,7%) и Мироновском институте пшеницы им В.М.Ремесло (100%), что является косвенным подтверждением ее значимости в детерминации хлебопекарных свойств, поскольку сорта Казахстанской селекции и этих селекционных учреждений, как правило, высококачественны. В сорте озимой мягкой пшеницы Богарная 56 выявлены биотипы, идентичные по спектру проламинов, но различающиеся по составу ВМСГ: у одного из них (типичного) субъединица 7 сопряжена с сопутствующим вариантом а, а у другого - с вариантом субъединицы b. По данным таблицы 1 , видно, что типичный биотип, имеющий в своем составе дополнительную субъединицу а, характеризуется более высокими показателями качества в сравнении с другим биотипом. Полученные данные свидетельствуют о том, что при полном следовании условиям фракционирования высокомолекулярных глютенинов в ДДС-Na ПААГе по Laemmli (1970) микрогетерогенность субъединиц нивелируется и в спектре насчитывается ограниченное число компонентов. Модификация метода, предложенная нами (Булатова, 1985), позволяет дифференцировать варианты субъединицы 7, контролируемой локусом Glu B1 и выявить в зоне электрофоретического спектра ВМСГ, традиционно считающейся месторасположением продуктов экспрессии локуса Glu B1, 4 варианта специфичных субъединиц, кодируемых D геномом. Присутствие определенных вариантов этих субъединиц сопряжено с показателями качества мягкой пшеницы, в связи с чем, их изучение и дискриминация с помощью модифицированного метода необходимы в оценке материала генофонда и в селекции высококачественных образцов. Генотипы пшеницы с сочетанием субъединиц 7 - «а» и 7 – «b» встречаются с достаточной частотой, тогда как образцов с вариантами субъединицы 7, сопряженной с «c» и «d» в генофонде практически нет, за исключением староместных сортов Западного Памира: Бабило и Сурхак Ишкашимский. С целью выявления селекционной значимости специфичных аллелей с участием этих сортов были получены, отобраны по урожайности и адаптивности к условиям Юго Востока Казахстана и переданы селекционерам 45 линий F3-F4 поколений. У значительной части сортов яровой пшеницы селекции НИИСХ Юго-Востока по результатам фракциони-рования в 10 %-ном полиакриламидном геле по прописи Laemmli (1970) субъединичный состав глютенинов был определен как 2*; 7+9; 2+12; (Morgunov A.I. et.al., 1990, Неттевич Э.Д. и др. 1991). Оценка качества по глю-тенину для таких сортов в соответствии с ранжированием Payne и др. (1987)      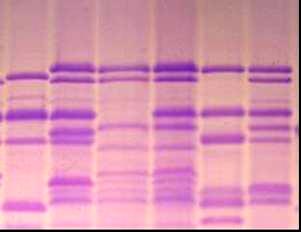 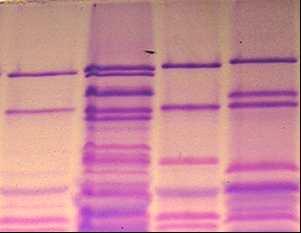 1 2 3 4 5 6 7 8 9 10 1- Наурыз 2; 2- Стекловидная 24; 3- дигаплоид Саратовской 52 с null мутацией в локусе Glu B1; 4-дигаплоид Саратовская 52 (контроль); 5 - Бабило; 6 - Ишкашимский Сурхак , 7,9,10 - образцы Aegilops cylindrica, Алматинская обл.; 8 - Богарная 56 (обозначения даны в тексте). Рисунок 1 – Спектры высокомолекулярных глютенинов Tr. Aestivum L., T.durum Desf. и Aegilops cylindrica.
1,2 – Чайниз Спринг; 3,4 – нуллисомик по 1D и тетрасомик по 1 B хромосомам; 5,6 - нуллисомик по 1D и тетрасомик по 1 А хромосомам; 7 - нуллисомик по 1B и тетрасомик по 1D хромосомам. Рисунок 2 – Электрофоретические спектры глютенинов у нулли-тетрасомных линий сорта Чайниз Спринг. равна 7, в то же время эти сорта известны как наиболее высококачественные среди яровых пшениц. В связи с этим, ряд авторов придерживается мнения, что показатель качества по оценке ВМСГ не всегда совпадает с характеристикой хлебопекарных показателей сортов яровой пшеницы, полученных классическими методами (Неттевич Э.Д и др. 1991, Абугалиева А.И. 2001). Отдельные случаи несоответствия хлебопекарных показателей качества муки бальной оценке по субъединицам глютенина лежат за пределами состава ВМСГ (Obukhova L. et.al., 2003). Причиной несоответствия реальных качественных показателей с бальной оценкой по составу ВМСГ может являться и различие в аллельном состоянии глютенинкодирующих локусов, не выявляемое по продуктам их биосинтеза при определенных условиях электрофореза. Таблица 1 – Показатели качества глютениновых биотипов сорта озимой мягкой пшеницы Богарная 56 с сочетанием 7 - «а» и 7 – «b» субъединиц глютенина.
Анализ состава ВМСГ сорта Саратовская 29 и ряда других сортов (таблица 2), проведенный в модифицированных условиях, показал, что локусом Glu D 1 детерминируется вариант ВМСГ, в котором “х” субъединица не совпадает по подвижности с положением субъединицы 2: она располагается на уровне ВМСГ 5, однако отличается от нее по интенсивности проявления. Следовательно, она не является продуктом аллели «d» и контролируется новой, ранее не описанной аллелью локуса Glu 1D. Состав ВМСГ этих сортов в связи с полученными данными регистрируется как 2*; 7+9; 5** +12, а не 2*; 7+9; 2 +12, как опубликовано указанными в таблице 2 авторами. Ряд исследователей предполагает, что линии с субъединицей у-типа 12 характеризуются лучшим качеством, нежели таковые с субъединицей 10 (Rodriguez-Quijano M.,et.al., 2000). Преимущество субъединицы х-типа 5 по вкладу в качество зерна доказано давно. По-видимому, сочетание таких ценных показателей в одном генотипе и обеспечивает высокое качество саратовских сортов и созданных с их участием форм. Еще один редкий аллельный вариант глютенинкодирующего локуса Glu B1, был выявлен в ходе изучения генетических изменений у регенерантных линий мягкой пшеницы по локусам, контролирующим биосинтез запасных белков зерна – глиадинов и глютенинов. В спектре высокомолекулярных глютенинов линии АД54, в сравнении с исходной линией выявлено отсутствие одной из высокомолекулярных субъединиц, классифицируемой по каталогу Payne P., Lawrence G. (1983) как х 7 ( рисунок 3), контролируемой локусом Glu B1. Мутантная линия с нулевой аллелью и ее исходная форма были изучены по ряду признаков урожайности и качества. Сравнение элементов урожайности показало превышение линии АД-54 над контрольным растением по ряду признаков урожайности, таких как: масса зерен с главного колоса и масса 1000 зерен (на 18,7% и 10,3%, соответственно). По ряду технологических показателей регенератная и исходная линия не отличались, однако по уровню седиментации АД-54 уступала исходной форме (29,5 мл. у регенеранта и 35,0 мл.у исходной формы). Результаты свидетельствуют, что отсутствие одной из ВМСГ, контролируемых локусом Glu B1 негативно отражается на показателях качества зерна и муки. Таблица 2- Состав ВМСГ сортов яровой мягкой пшеницы по прописи Laemmli (1970) и в модифицированных условиях.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||