гистология ответы на экзамен Г. Ответы на экзаменационные вопросы по гистологии Часть 1. Экзаменационные вопросы по гистологии 1 Определение и задачи гистологии
 Скачать 308.57 Kb. Скачать 308.57 Kb.
|

|
№40 Теории кроветворения; роль гистологии в развитии гематологии. Первая попытка обобщения имеющихся материалов в виде теории кроветворения была предпринята в 1880 году Эрлихом - была предложена дуалистическая теория кроветворения: из отдельных 2- родоначальных клеток начинается и происходит лимфоцитопоэз и миелопоэз. В начале ХХ века Ашоф и Шиллинг предложили триалистическую теорию кроветворения - т.е. к 2-м родоначальным клеткам лимфоцитопоэза и миелопоэза был добавлен третья отдельная родоначальная клетка для моноцитопоэза. Существовала еще полифилитическая теория, предполагающая наличие отдельных родоначальных клеток для каждой разновидности форменных элементов крови. Основоположником современной унитарной теории кроветворения является отечественный гистолог Максимов (работал на кафедре гистологии ВМА в С-Петербурге). Еще в 1907 году Максимов утверждал, что все клетки крови развиваются из единой одной и той же родоначальной клетки; мало того, он назвал эту клетку - морфологически это малый лимфоцит. Однако имеющиеся в то время методы исследований не позволяли экспериментально доказать верность этой теории. Максимов в ходе гемоцитопоэза клетки крови подразделял на 4 группы: 1 группа - клетки с неограничанной возможностью превращений, т.е. родоначальная клетка, способная развиваться и превратиться в любой форменный элемент крови. 2 группа - клетки с частично ограниченный способностью развиваться в ту или иную форму клеток крови. 3 группа - клетки со строго ограничанной возможностью развития. 4 группа - клетки крови не способные изменяться. Последующие исследования показали верность унитарной теории кроветворения Максимова. Отечественные ученые Кассирский, Алексеев внесли существенный вклад в области цитохимических и электронно-микроскопических исследований клеток крови в разных стадиях гемоцитопоэза. Канадские исследователи Till и Mc-Culloch при помощи оригинальной серии экспериментов со смертельно облученными мышами доказали существование стволовых кроветворных клеток (СКК). Современная схема кроветворения в варианте, который Вы будете изучать, составлена в 1973 году Чертковым и Воробьевым. Согласно этой схеме все клетки крови в процессе гемцитопоэза подразделены на 6 классов. 1-й класс - полипотентные стволовые кроветворные клетки (ПСКК). Морфологически выглядат как малые темные лимфоциты. В норме у здорового человека у ПСКК обмен веществ на низком уровне, 80% ПСКК находится в G0 фазе, т.е. в покое - не делятся. ПСКК полипотентны - могут дифференцироваться в любую клетку крови, способны к самоподдержанию - автоматически поддерживается определенное количество ПССК в организме. При необходимости способны к ускоренной пролиферации, 1 клетка может дать до 100 митозов. Активность ПСКК регулируется микроокружением и гуморально - гемопоэтинами. 2-й класс - полустволовые клетки (ПСК) - клетки предшественники миелопоэза, клетки предшественники лимфопоэза. Взаимопереход этих клеток еще возможен при изменении специфического микроокружения. Морфологически выглядат как малые темные лимфоциты. 3-й класс - унипотентные предшественники, имеется отдельный предшественник для каждого форменного элемента крови. Взаимопереход между направлениями дифференцировки становится невозможным. Морфологически выглядат как малые темные лимфоциты. Если все клетки 1-3 класса между собой морфологически не различимы и все выглядат как малые темные лимфоциты, то начиная с 4-го класса созревающие клетки становятся морфологически идентифицируемыми. 4-й класс - бластные клетки, дифференцируются в строго определенном направлении, морфологически различимы. 5-й класс - созревающие клетки. В клетках появляются специфические для каждой клетки структуры, клетки постепенно теряют способность к делению. 6-й класс - зрелые клетки крови. №41 Эмбриональное (первичное) кроветворение. Кроветворение в стенке желточного мешка. У человека начинается в конце 2й – в начале 3й недели эмбрионального развития. В мезенхиме стенки желточного мешка обособляются зачатки сосудистой крови, или кровяные островки. Мезенхимные клетки в них округляются, теряют отростки и преобразуются в стволовые клетки (СК). Клетки, ограничивающие кровяные островки, уплощаются, соединяются между собой и образуют эндотелиальную выстилку будущего сосуда. Часть стволовых клеток дифференцируется в первичные клетки (бласты). Большинство первичных кровяных клеток митотически делится и превращается в первичные эритробласты, характеризующиеся крупным размером (мегалобласты). Это превращение совершается в связи с накоплением гемоглобина в цитоплазме бластов, при этом сначала образуются полихроматофильные эритробласты, а затем оксифильные эритробласты с большим содержанием гемоглобина. Такой тип кроветворения называется МЕГАЛОБЛАСТИЧЕСКИМ. Наряду с мегалобластическим в стенке желточного мешка начинается нормобластическое кроветворение, при котором из бластов образуются вторичные эритробласты, сначала они превращаются в полихроматофильные эритробласты, далее в нормобласты, из которых образуются вторичные эритроциты (нормоциты). Развитие эритроцитов в стенке желточного мешка происходит внутри первичные кровеносных сосудов, т.е. интраваскулярно. Одновременно экстраваскулярно из бластов, расположенных вокруг сосудистых стенок, дифференцируется небольшое количество гранулоцитов – нейтрофилов и эозинофилов. После редукции желточного мешка основным органом кроветворения становится печень. На 3-4-й неделе жизни эмбриона закладывается печень, которая уже на 5-й неделе жизни эмбриона становится центром кроветворения. Гемоцитобласты в печени возникают из окружающих капилляры клеток печеночных долек. Из этих гемоцитобластов образуются вторичные эритроциты. Одновременно из других клеток происходит образование гранулоцитов. Кроме того, в кроветворной ткани печени формируются гигантские клетки, или мегакариоциты, из которых образуются тромбоциты. К концу внутриутробного периода кроветворение в печени прекращается. Универсальный кроветворный орган в первой половине эмбриональной жизни представляет собой селезенка. В ней развиваются все клетки крови. По мере роста плода образование эритроцитов в селезенке и в печени угасает, и этот процесс перемещается в костный мозг, который впервые закладывается в конце 2-го месяца эмбриональной жизни в ключицах, а позднее — и во всех других костях. На втором месяце внутриутробного развития закладывается вилочковая железа, в которой начинается образование лимфоцитов, в дальнейшем расселяющихся в другие лимфоидные органы. У 3-месячного плода в области шейных лимфатических мешков начинают формироваться зачатки лимфатических узлов. На ранних стадиях развития в них образуются лимфоциты, гранулоциты, эритроциты и мегакариоциты. Позже образование гранулоцитов, эритроцитов, и мегакариоцитов подавляется, и продуцируются только лимфоциты — основные элементы лимфоидной ткани. К моменту рождения ребенка процессы кроветворения усиливаются. №42 Дефинитивное (вторичное) кроветворение. Постэмбриональный гемоцитопоэз представляет собой процесс физиологической регенерации крови, который совершается в гемопоэтических тканях (миелоидной). Миелопоэз происходит в миелоидной ткани, расположенной в эпифизах трубчатых и полостях многих губчатых костей. Здесь развиваются форменные элементы крови: эритроциты, гранулоциты, моноциты, кровяные пластинки, предшественники лимфоцитов. В миелоидной ткани находятся стволовые клетки крови и соединительной ткани. Лимфопоэз происходит в лимфоидной ткани (тимус, селезенка, лимфоузлы). Она выполняет 3 основные функции ( образование лимфоцитов, образование плазмоцитов и удаление клеток и продуктов их распада). Дефинитивное кроветворение, или физиологическая регенерация крови, представляет собой многоступенчатый процесс, в котором различают молодые гемопоэтические, компетентные, коммитирован-ные, дифференцирующиеся и специализированные клетки. Процесс образования гемоцитов начинается с тотипотентной стволовой клетки, которая обладает масштабными пролиферативными способностями. Клетки, возникающие из нее, (полипотентные) обладают ограниченными потенциями. Попадая в разное окружение, дают начало лимфоцитопоэзу и миелопоэзу. Размножение и дифференцировка полипотентных клеток дает начало третьему классу - унипотентным колониеобразующим клеткам или единицам КОЕ; КОЕ моноцитов, КОЕ миелоцитов (нейтрофилов, эозинофилов, базофилов) и КОЕ эритроцитов. В результате размножения клеток КОЕ возникает четвертый класс гемопоэтических клеток - компетентные бласты. Индуцибельные гены ядер бластных элементов обеспечивают программу реализации диф-ференцировки и специализации пятого класса гемоцитов - проклеток. №43 Стволовая кроветворная клетка; доказательства ее наличия. Стволовые клетки являются полипотентными предшественниками всех клеток крови и относятся к самоподдерживающейся популяции клеток. Они редко делятся. Впервые представление о родоначальных клетках крови сформулировал в начале 20 века А.А. Максимов, который считал, что по своей морфологии они сходны с лимфоцитами. В настоящее время это представление нашло подтверждение и дальнейшее развитие в новейших экспериментальных исследования, проводимых главным образом на мышах. Выявление СКК стало возможными при применении метода колониеобразования. Экспериментально показано, что при введении смертельно облученным животным (утратившим собственные стволовые клетки) взвеси клеток красного костного мозга или фракции, обогащенной СКК, в селезенке появляются колонии клеток – потомков одной СКК. Пролиферативную активность СКК модулируют колонистимулирующие факторы (КСФ), ИЛ-3. Каждая СКК в селезенке образует одну колонию и называется селезеночной колониеобразующей единицей (КОЕ-С). Подсчет колоний позволяет судить о количестве стволовых клеток, находящихся во введенной взвеси клеток. Исследования клеточного состава колоний позволило выявить 2 линии их дифференцировки. Одна линия дает начало мультипотентной клетке – родоначальнице гранулоцитарного, моноцитарного, эритроцитарного и мегакариоцитарного рядов гемопоэза (КОЕ-ГЭММ). Вторая линия дает начало мультипотентной клетке – родоначальнице лимфопоэза (КОЕ-Л). Из мультипотентных клеток дифференцируются олигопотентные (КОЕ-ГМ) и унипотентные родоначальные клетки. Методом колониеобразования определены родоначальне унипотентные клетки для моноциотов, нейтрофильных гранулоцитов, эозинофилов, базофилов, эритроцитов, мегакариоцитов, из которых образуются клетки-предшественники. Полипотентные, олигопотентные и унипотентные клетки морфологически не различаются. №44 Эритропоэз, стадии и клеточные формы. Понятие об эритроне. Родоначальницей эритроидных клеток человека, как и других клеток крови, является полипотентная стволовая клетка крови (СКК), способная формировать колонии. Дифференцирующаяся полипотентная СКК дает 2 типа мультипотентных частично коммитированных СКК: 1) коммитированные к лимфоидному типу дифференцировки, 2) КОЕ-ГЭММ, единицы, образующие смешанные колонии, состоящие из гранулоцитов, эритроцитов, моноцитов и мегакариоцитов. Из второго типа мультипотентных СКК дифференцируются унипотентные единицы: бурстообразующая (БОЕ-Э) и колониеобразующая (КОЕ-Э) эритроидные клетки, которые являются коммитированными родоначальными клетками эритропоэза. БОЕ-Э, по сравнению с КОЕ-Э, является менее дифференцированной и при интенсивном размножении быстро образует крупную колонию клеток и малочувствительна к эритропоэтину. КОЕ-Э более зрелая клетка, образует более мелкие колонии и чувствительна к эритропоэтину. Образующиеся из КОЕ-Э эритроидные клетки морфологически идентифицируются. Сначала образуется проэритробласт. Проэритробласт – клетка, имеющая круглое ядро с мелкозернистым хроматином, 1-2 ядрышка, цитоплазму со средней базофилией, в которой содержатся свободные рибосомы и полисомы , слаборазвитый аппарат Гольджи и гранулярная ЭПС. Базофильный эритробласт – клетка меньшего размера, содержит больше гетерохроматина. Цитоплазма клетки обладает выраженной базофильностью в связи с накоплением в ней рибосом, в которых начинается синтез гемоглобина. Полихроматофильный эритробласт – ядро содержит много гетерохроматина. Следующая стадия дифференцировки – образование оксифильного эритрона (нормобласта). Это клетка небольшого размера, имеющая маленькое ядро. В цитоплазме эритробласта содержится много гемоглобина, обеспечивающего его оксифилию. Ретикулоцит – безъядерная клетка с небольшим содержанием рибосом, обусловливающих наличие участков базофилии, и преобладанием гемоглобина. При выходе в кровь ретикулоцит созревает в эритроцит в течение 1-2 суток. Эритроцит – клетка, образующаяся на конечной стадии дифференцировки клеток эритроидного ряда. Имеет форму двояковогнутого диска. Период образования эритроцита занимает 7 дней, его продолжительность жизни – 120 дней. Таким образом, в процессе эритропоэза происходят уменьшение размера клетки в 2 раза, уменьшение размера и уплотнение ядра и его выход из клетки, уменьшение содержания РНК, накопление гемоглобина, потеря способности к делению. Из одной СКК в теч. 7-10 дней в результате 12 делений образуется 2048 зрелых эритроцитов. Эритрон — совокупность незрелых и зрелых, неподвижных и циркулирующих, расположенных интра- и экстравазально клеток эритроцитарного ряда, находящихся на всех стадиях развития — образования (резервный, пролиферирующий, созревающий пул в кроветворной ткани), функционирования (циркулирующий пул в крови) и гибели (в макрофагоцитах органов кроворазрушения). №45 Гранулоцитопоэз, стадии и клеточные формы. Дифференцировка и созревание клеток гранулоцитопоэза происходит в костном мозге, где из коммитированных, морфологически неидентифицируемых клеток-предшественников КОЕ-ГМ (колониеобразующая единица грануломоноцитопоэза) и КОЕ-Г (колониеобразуюшая единица гранулоцитопоэза) формируется пул пролиферирующих гранулоцитов, состоящий из миелобластов, промиелоцитов и миелоцитов. Все эти клетки характеризуются способностью к делению. Другой пул, образующийся в костном мозге – это непролиферирующие (созревающие) клетки – метамиелоциты, палочкоядерные и сегментоядерные гранулоциты. Созревание клеток сопровождается изменением их морфологии: уменьшением ядра, конденсацией хроматина, исчезновением ядрышек, сегментацией ядра, появлением специфической зернистости, утратой базофилии и увеличением объема цитоплазмы. Процесс формирования зрелого гранулонита из миелобласта осуществляется в костном мозге в течение 10 13 дней. Регуляция гранулоцитопоэза обеспечивается колониестимулирующими факторами: ГМ-КСФ (гранулоцитарно-макрофагальный фактор) и Г-КСФ (гранулоцитарный колониестимулирующий фактор), действующими до конечной стадии созревания гранулоцитов. На стадии поздних миелобластов и промиелоцитов происходит образование первичных гранул (азурофильной зернистости), специфическим маркером которых является миелопероксидаза. В цитоплазме миелоцитов начинается формирование специфической зернистости (вторичные гранулы). Маркерами вторичных гранул являются лактоферрин, катионный белок кателицидии, В12-связываюший белок и другие факторы. В состав вторичных гранул также входит лихоцим, коллагеназа, металлопротеиназы. Количество вторичных гранул увеличивается в клетке но мере ее созревания, в зрелых сегмеитоядериых гранулоцитах на их долю приходится 70-90%, остальные 10-30% составляет азурофильная зернистость. Зрелые гранулоциты костного мозга образуют гранулоцитарный костномозговой резерв, насчитывающий около 8,8 млрд/кг и мобилизуемый в ответ на специфический сигнал при бактериальных инфекциях. Покидая костный мозг, гранулоциты представляют собой полностью дифференцированные клетки, имеющие полный спектр поверхностных рецепторов и цитоплазматических гранул с набором многочисленных биологически активных веществ. Нейтрофилы составляют 60 70% общего числа лейкоцитов крови. После выхода нейтрофильных гранулоцитов из костного мозга в периферическую кровь часть их остается в свободной циркуляции в сосудистом русле (циркулирующий пул), другие занимают пристеночное положение, образуя маргинальный пул. Зрелый нейтрофил пробывает в циркуляции 8 10 часов, затем поступает в ткани, образуя по численности значительный пул клеток. Продолжительность жизни нейтрофильного гранулоцита в тканях составляет 2-3 дня. Функцией нейтрофилов является участие в борьбе с микроорганизмами путем их фагоцитоза. Содержимое гранул способно разрушить практически любые микробы. В нейтрофилах содержатся многочисленные ферменты (кислые протениазы, миелопероксидаза, лизоцим, лактоферрин, целочная фосфатаза и др.), вызывающие бактериолиз и переваривание микроорганизмов. Эозинофилы составляют 0.5-5% от всех лейкоцитов крови, циркулируют в течение 6-12 часов, после чего поступают в ткани, срок полужизни – 12 суток. В клетках содержится значительное количество гранул, основным компонентом которых является главный щелочной белок, а также перекиси, обладающие бактерицидной активностью. В гранулах выявляются кислая фосфатаза, арилсульфатаза, коллагеназа, эластаза, глюкуроиидаза, катепсин, миелонероксидаза и другие ферменты. Обладая слабой фагоцитарной активностью, эозинофилы обусловливают внеклеточный цитолиз, тем самым участвуя в противогельминтном иммунитете. Другой функцией этих клеток является участие в аллергических реакциях. Базофилы и тучные клетки имеют костномозговое происхождение. Предполагают, что предшественники тучных клеток покидают костный мозг и через периферическую кровь попадают в ткани. Дифференцировка базофилов в костном мозг длится 1,5-5 суток. Ростовым фактором базофилов и тучных клеток являются ИЛ-3, ИЛ-4. Созревшие базофилы поступают в кровоток, где период их полужизни составляет около 6 часов. На долю базофилов приходится всего 0,5% от общего числа лейкоцитов крови. Базофилы мигрируют в ткани, где через 1-2 суток после осуществления основной эффекторной функции гибнут. В гранулах этих клеток содержатся гистамин, хондроигинсульфаты А и С, гепарин, серотонин, ферменты (трипсин, химотринсии, пероксидаза, РНК-аза и др.). Базофилы имеют на клеточной мембране высокую плотность рецепторов к IgE, обеспечивающих не только связывание IgE, но и освобождение гранул, содержимое которых обусловливает развитие аллергических реакций. Базофилы также способны к фагоцитозу. Тучные клетки крупнее базофилов, имеют округлое ядро и много гранул, которые по составу аналогичны гранулам базофилов. 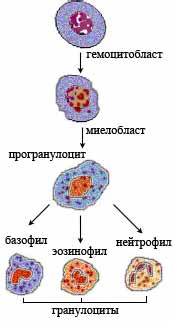 №46 Гуморальная и нервная регуляция гемопоэза. Регуляция гемопоэза - гемопоэз или кроветворение происходит под влиянием различных факторов роста, которые обеспечивают деление и дифференцировку клеток крови в красном костном мозге. Выделяют две формы регуляции: гуморальную и нервную. Нервная регуляция осуществляется при возбуждении адренэргических нейронов, при этом происходит активация гемопоэза, а при возбуждении холинэргических нейронов - торможение гемопоэза. Гуморальная регуляция происходит под действием факторов экзо- и эндогенного происхождения. К эндогенным факторам относятся: гемопоэтины (продукты разрушения форменных элементов), эритропоэтины (образуются в почках при снижении концентрации кислорода в крови), лейкопоэтины (образуются в печени), тромбоцитопоэтины: К (в плазме), С (в селезенке). К экзогенным витамины: В3 - образование стромы эритроцитов, В12 - образование глобина; микроэлементы (Fe, Cu...); внешний фактор Касла. А также такие факторы роста как: интерлейкины, колониестимулирующие факторы КСФ, факторы транскрипции — специальные белки, регулирующие эксп-рессию генов гемопоэтических клеток. Кроме этого большую роль играет строма костного мозга, которая создает гемопоэтическое микроокружение, необходимое для развития, дифференциации и созревания клеток. Таким образом регуляция гемопоэза представляет собой единую систему, состоящую из нескольких взаимосвязанных звеньев каскадного механизма, которая реагирует на изменяющиеся условия внешней и внутренней среды и различные патологические состояния (при сильной анемии – снижении содержания эритроцитов, снижении содержания лейкоцитов, тромбоцитов, факторов свертывания крови, острой кровопотери и т.д.). Угнетение гемопоэза происходит под действием ингибирующих факторов. К ним относятся продукты образуемые клетками на последних этапах созревания ( простагландины, цитокины и др.) №47 Общая характеристика иммунной системы и иммуноцитов. Иммунная система объединяет органы и ткани, в которых происходит образование и взаимодействие клеток — иммуноцитов, выполняющих функцию распознавания генетически чужеродных субстанций (антигенов) и осуществляющих специфические реакции защиты. Иммунитет — это способ защиты организма от всего генетически чужеродного — микробов, вирусов, от чужих клеток или генетически измененных собственных клеток. Иммунная система обеспечивает поддержание генетической целостности и постоянства внутренней среды организма, выполняя функцию распознавания «своего» и «чужого». В организме взрослого человека она представлена: красным костным мозгом - источником стволовых клеток для иммуноцитов, центральным органом лимфоцитопоэза (тимус), периферическими органами лимфоцитопоэза (селезенка, лимфатические узлы, скопления лимфоидной ткани в органах), лимфоцитами крови и лимфы, а также популяциями лимфоцитов и плазмоцитов, проникающими во все соединительные иэпителиальные ткани. Все органы иммунной системы функционируют как единое целое благодаря нейрогуморальным механизмам регуляции, а также постоянно совершающимся процессам миграции и рециркуляции клеток по кровеносной и лимфатической системам. Главными клетками, осуществляющими контроль и иммунологическую защиту в организме, являются лимфоциты, а также плазматические клетки и макрофаги. Постоянно перемещающиеся лимфоциты осуществляют «иммунный надзор». Они способны «узнавать» чужие макромолекулы бактерий и клеток различных тканей многоклеточных организмов и осуществлять специфическую защитную реакцию. Для понимания роли отдельных клеток в иммунологических реакциях необходимо прежде всего дать определение некоторым понятиям иммунитета. |