|
Фибус Левин предположил, что нк состоят из мономеров нуклеотидов. А в 1930 г
Назовите основные этапы в истории изучения нуклеиновых кислот. В каких экспериментах была доказана роль ДНК в хранении и передаче наследственной информации?
Нуклеиновые кислоты (НК) входят в состав клеток всех живых организмов. Они играют главную роль в хранении, передаче наследственной информации и ее реализации в процесса синтеза белков.
- НК были обнаружены были обнаружены в XIX веке щвейцарским врачом Иоганом Фридрихом Мишером.
- В 1868-69 гг. он выделил из лейкоцитов неизвестное высокомолекулярное вещество, которые было названо нуклеином.
- В 1879-88 гг. немецкий ученый Альбрехт Коссель и Эмиль Фишер нашли в составе нуклеина пиримидиновые и пуриновые азотистые основания. Тогда же немецкий ученый Рихард Альтман впервые ввел термин «Нуклеиновая кислота» и разработал удобный способ выделения этих веществ.
- В 1909 г. Фибус Левин предположил, что НК состоят из мономеров – нуклеотидов. А в 1930 г. установил различия нуклеотидов по типу углеводной компоненты. НК, содержащие рибозу – РНК, дезоксирибозу – ДНК.
- В 1934 г. Торбьёрн Оскар Касперссон вместе с Эйнаром Хаммерстеном показали, что НК — это гетерополимеры, мономерами которых являются нуклеотидные звенья.
- Одновременно с изучением структуры нуклеиновых кислот проводились исследования, направленные на установление их биологической роли.
- В 1881 г. Эдуард Захариас доказал, что нуклеин содержится в хромосомах, которые уже тогда считались ответственными за наследственные признаки. Однако идея о том, что нуклеиновые кислоты являются материальной основой наследственности, в науке не утвердилась. Так, советский биолог Н.К. Кольцов считал, что за передачу наследственности отвечает белковая компонента хромосом. Считалось также, что РНК встречается только у растений, а ДНК – у животных.
- И только в 1936 г. благодаря работам Ж. Браше и Дж. Дэвидсона, а также А.Н. Белозерского стало ясно, что оба вида нуклеиновых кислот присуще как для животных, так и для растений.
- Ф. Гриффит (1928 г.), О. Эвери (1944 г.), А. Херши (1952 г.) опытным путем доказали, что нуклеиновые кислоты обеспечивают хранение, передачу и реализацию наследственной информации в процессе синтеза белка в клетке. Они пометили ДНК и капсид радиоактивным фосфором. Заразили вирусом кишечную палочку и пришли к выводу, что внутрь клетки попадает ДНК, а капсид остается в питательной среде. Таким образом, было доказано, что ДНК является носителем наследственной информации.
- В 1950-е г. Э. Чаргафф установил строгие количественные соотношения азотистых оснований в молекулах ДНК, при помощи бумажной хроматографии которые получили название правил Чаргаффа.
- Важнейшие сведения были получены благодаря рентгеноструктурным исследованиям биомолекул. Л. Полинг продемонстрировал возможность спирализации белковых молекул. Первые рентгенограммы молекул ДНК, полученные английскими биофизиками М.Уилкинсом и Р. Франклин свидетельствовали о наличии в их пространственной структуре элементов периодичности.
- Основываясь на этих открытиях, в 1953 г. американский генетик Джеймс Уотсон и английский физик Френсис Крик сформулировали основные представления о структуре двойной спирали ДНК. Модель ДНК Уотсона-Крика позволила объяснить многие фундаментальные биологические феномены, такие как структурная организация молекул ДНК, способ хранения и точного копирования генетической информации, возможность изменения структуры генов в процессе эволюции.
- В 1957 г. Ф.Крик сформулировал центральную догму молекулярной биологии (рис.1), согласно которой наследственная информация ( генетическая) информация реализуется однонаправленно. Заключенная в последовательностях нуклеотидов ДНК информация о первичной структуре белка сначала переписывается на молекулу РНК (процесс транскрипции), а затем в процессе биосинтеза белка реализуется в последовательности аминокислотных остатков полипептидной цепи (процесс трансляции). Согласно этой концепции, ДНК является хранилищем информации о структуре белка. Посредником между ними является РНК, реализующая генетическую информацию, заключенную. в ДНК, в структуру белка.
- В конце 1950 – начале 1960-х годов были выделены различные типы РНК:
Матричная
Транспортная
Рибосомная РНК
- Открыты рибосомы, состоящие из РНК и белков.
В этот период были раскрыты молекулярные механизмы наследования признаков, расшифрован генетический и изучены матричные процессы:
репликации (удвоения) ДНК
транскрипции (биосинтез РНК)
трансляции (биосинтез белка)
Доказательства роли ДНК в передаче наследственной информации. Опыты Херши и Чейз.
Херши и Чейз работали с бактериофагами (конкретно с Т2). Пометили ДНК и капсид радиоактивным фосфором. Заразили вирусом кишечную палочку и пришли к выводу, что внутрь клетки попадает ДНК, а капсид остается в питательной среде. Таким образом, было доказано, что ДНК является носителем наследственной информации. Позже было обнаружено, что у некоторых видов вирусов наследственная информация содержится в РНК. Но тем неменее носителем генитческой информация всегда остается нуклеиновые кислоты.
Опишите строение нуклеотидов; какие химические связи формируют и стабилизируют молекулы НК. Сформулируйте правила Чаргаффа.
Каждый нуклеотид состоит из азотистого основания, пятиуглеродного сахара (пентозы) и остатка фосфорной кислоты. В состав нуклеотида может входить одно из пяти азотистых оснований: аденин (А), гуанин (Г), цитозин (Ц), тимин (Т) или урацил (У). Аденин и гуанин относятся к пуриновым основаниям, цитозин, тимин и урацил — к пиримидиновым. Молекула ДНК состоит из двух полинуклеотидных цепей, соединенных между собой водородными связями. Эти связи возникают между азотистыми основаниями нуклеотидов противоположных цепей: между аденином и тимином возникают две водородные связи, а между гуанином и цитозином — три.
Первичная структура-последовательность расположения мононуклеотидов в полинуклеотидной цепи ДНК и РНК. Нуклеотиды в молекулах нуклеиновых кислот соединены сложноэфирной связью, образованной остатком фосфорной кислоты одного мононуклеотида и 3’-гидроксильной группой остатка пентозы другого мононуклеотида (3’,5’-фосфодиэфирная связь).
Вторичная структура.ДНК состоит из двух цепей, закрученных вокруг одной и той же оси, образующих правовращающую спираль. Внутри спирали находятся азотистые основания, снаружи - углеводные компоненты (рис. 12). Обе цепи в молекуле ДНК имеют противоположную полярность: межнуклеотидная связь в одной цепи имеет направление 5’®3’, в другой – 3’®5’.
В молекуле ДНК основания уложены парами: пуриновые из одной цепи и пиримидиновые из другой. Взаимодействие пар А-Т и Г-Ц называют комплементарностью, а соответствующие азотистые основания – комплементарными (рис. 13). Цепи ДНК комплементарны друг другу. Стабильность
А-Т пар обеспечивается двумя водородными связями, а пар Г-Ц-тремя.
Изучая состав ДНК, Чаргафф (1949г.) установил важные закономерности соотношения АО в ДНК, что в дальнейшем помогло раскрыть их структуру.
1) сумма пуриновых оснований = сумме пиримидиновых оснований.
А + Г = Ц + Т или 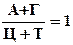
2) соединение А = соединению Т
А = Т или 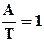
3) соединение Г = соединению Ц
Г = Ц или 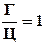
4) сумма А и Ц = сумме Г и Т
А+Ц = Г+Т или 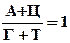
5) изменчива только сумма А+Т и Г+Ц
если А+Т > Г+Ц, то это АТ-тип ДНК
если наоборот, то это ГЦ-тип ДНК.
Эта разница выражается через коэффициент специфичности, который является важной характеристикой молекулы ДНК.
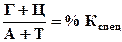 (в % от суммы Г+Ц от 0,54 до 0,94 для живых, то есть преобладает АТ-тип; от 0,45 до 2,57 – для микроорганизмов). (в % от суммы Г+Ц от 0,54 до 0,94 для живых, то есть преобладает АТ-тип; от 0,45 до 2,57 – для микроорганизмов).
Правила Чаргаффа характерны только для ДНК, но не свойственны РНК.
Опишите модель ДНК, предложенную Уотсоном и Криком. Какие существуют формы
Структура ДНК, в модели Уотсона и Крика, представляет собой двухцепочечную антипараллельную правостороннюю спираль. Сахарофосфатный остов цепей ДНК образует внешнюю часть спирали, в то время как азотистые основания находятся внутри и образуют пары, связанные водородными связями, которые удерживают две цепочки ДНК вместе.
В приведенной ниже модели оранжевые и красные атомы обозначают фосфаты в сахарофосфатном остове, а синие атомы внутри спирали принадлежат азотистым основаниям.
В модели Уотсона и Крика две нити двойной спирали ДНК удерживаются вместе водородными связями между азотистыми основаниями на противоположных нитях. Каждая пара оснований лежит перпендикулярно нити, образуя «ступеньку» на «лестнице» молекулы ДНК.
Пары оснований не состоят из какой-то случайной комбинации оснований. Вместо этого, каждому A находящемуся на одной нити, соответствует T, находящийся на противоположной нити (и наоборот). Точно так же G, находящийся на одной цепи, всегда будет иметь в качестве пары C на противоположной цепи. Эти связи A-T и G-C известны как комплементарные пары оснований.
Диаграмма, иллюстрирующая попарное соединение оснований между собой A-T и G-C. А и Т находящиеся напротив друг друга на двух нитях спирали, и их функциональные группы образуют две водородные связи, которые удерживают нити вместе. Точно так же G и C находятся напротив друг друга на двух нитях, и их функциональные группы образуют три водородные связи, которые удерживают цепи вместе.
Формы:
В-форма ДНК – основной тип двойной спирали. В двойной спирали на один виток приходится 10 пар нуклеотидов (п.н.), длина витка – 3,4 нм. Расстояние между соседними нуклеотидами составляет 0,34 нм, диаметр спирали – 2,0 нм.
^ А-форма ДНК образуется при дегидратации. Характеризуется более широкой и короткой спиралью, основания имеют более сильный наклон к оси спирали (+19). На один виток спирали приходится 11 пар оснований. Расстояние между соседними нуклеотидами составляет 0,23 нм, длина витка – 2,5 нм, диаметр спирали – 2,3 нм. А-форму имеет спираль РНК-ДНК в комплексе матрица-затравка, а также спираль РНК-РНК и шпилечные структуры РНК (2’-гидроксильная группа рибозы не позволяет молекулам РНК образовывать В-форму). А-форма ДНК обнаружена в спорах. Установлено, что А-форма ДНК в 10 раз устойчивее к действию УФ-лучей, чем В-форма.
^ Z-форма ДНК – это двойная спираль, закрученная влево. На один виток этой формы приходится 12 пар оснований. Расстояние между соседними нуклеотидами составляет 0,38 нм, длина витка – 4,56 нм, диаметр Z-ДНК – 1,8 нм. Z- форма ДНК обнаружена в клетках прокариот и эукариот. Обычно Z-форма образуется в участках ДНК, где пурины чередуются с пиримидинами (например, 5’-ГЦГЦГЦ-3’), или в повторах 5’-ЦГЦГЦГ-3’, содержащих метилированный цитозин.
В клетках ДНК обычно существует в В-форме, которая является самой стабильной, но отдельные участки ДНК могут находиться в А-форме, Z-форме или даже в другой форме. Некоторые участки молекулы ДНК со специфическими последовательностями образуют изгибы, что имеет важное значение для регуляции экспрессии генов.
Опишите три модели репликации ДНК. Каким способом происходит удвоение ДНК в живой клетке.
Три модели репликации ДНК
Существовали три основные модели репликации ДНК, которые были предложены научным сообществом после открытия структуры ДНК.
Полуконсервативная репликация. В этой модели две нити ДНК раскручиваются и отсоединяются друг от друга, далее каждая из них выступает в качестве матрицы для синтеза новой комплементарной нити. В результате чего получаются две молекулы ДНК, каждая из которых содержит одну исходную и одну новую цепи.
Консервативная репликация. В этой модели результатом репликации ДНК является одна молекула, содержащая обе исходных цепи ДНК (идентичная исходной молекуле ДНК), и вторая молекула, состоящая из двух новых цепей (которые являются точными копиями цепей исходной молекулы).
Дисперсионная репликация. В дисперсионной модели результатом репликации ДНК являются две молекулы ДНК, каждая из которых представляет собой смесь, или «гибрид» родительской и дочерней ДНК. В этой модели каждая отдельная нить подобна лоскутному одеялу и состоит из фрагментов исходной и новой ДНК.
Перед каждым клеточным делением при абсолютно точном соблюдении нуклеотидной последовательности происходит самоудвоение (редупликация) молекулы ДНК. Редупликация начинается с того, что двойная спираль ДНК временно раскручивается. Это происходит под действием фермента ДНК-полимеразы в среде, в которой содержатся свободные нуклеотиды. Каждая одинарная цепь по принципу химического сродства (А - Т, Г - Ц) притягивает к своим нуклеотидным остаткам и закрепляет водородными связями свободные нуклеотиды, находящиеся в клетке. Таким образом, каждая полинуклеотидная цепь выполняет роль матрицы для новой комплиментарной цепи. В результате получаются две молекулы ДНК, у каждой из них одна половина происходит от родительской молекулы, а другая является вновь синтезированной, т.е. две новые молекулы ДНК представляют собой точную копию исходной молекулы.
Опишите эксперименты Мезельсона и Сталя по доказательству полуконсервативного способа репликации ДНК.
Мезельсон и Сталь ставили свои знаменитые эксперименты по репликации ДНК, используя бактерии E. coli в качестве модельной системы.
Они начали с наращивания E. coli на питательной среде, также известной как питательный бульон, содержащей "тяжелый" изотоп азота, N15 (изотоп - это просто вариант химического элемента, который отличается от других вариантов количеством нейтронов в ядре атома.) При выращивании на среде, содержащей «тяжёлый» азот N15, бактерии используют его для синтеза новых молекул, в том числе ДНК.
После многих поколений, выросших на среде с изотопом N15 азотистые основания ДНК бактерий были отмечены тяжелым N15. Затем бактерии были помещены на среду, содержащую «легкий» изотоп N14 и позволяли расти на ней еще в течение нескольких поколений. ДНК, синтезированная после переключения, должна была состоять из N14 поскольку это был единственный азот, доступный для её синтеза.
Мезельсон и Сталь знали, с какой частотой делились клетки E. coli , поэтому они могли собирать небольшие образцы каждого поколения бактерий, а затем выделять из них ДНК. После чего они измеряли плотность ДНК (и, косвенно, содержание в ней изотопов N15 и N14, используя центрифугирование в градиенте плотности.
Этот метод разделяет молекулы, такие как, например, ДНК, на фракции при помощи вращения их на высоких скоростях в присутствии других молекул, например, хлорида цезия. В результате от верха вращающейся пробирки до её донышка образуется градиент плотности. Центрифугирование в градиенте плотности позволяет обнаруживать очень малые различия в плотности, такие, как, например, между ДНК меченой тяжелым изотопом N15 и легким изотопом N14.
Поколение 0
ДНК, выделенная из клеток в начале эксперимента («поколение 0», прямо перед переходом к среде с изотопом N14 давала одну фракцию после центрифугирования. Этот результат был вполне логичным, поскольку в этот момент ДНК бактерий должна была содержать только тяжелый N15. Поколение 1
ДНК, выделенная из бактерий первого поколения (после одного цикла репликации ДНК), также показывала одну фракцию при центрифугировании. Однако эта фракция располагалась уже выше, что указывало на промежуточную плотность между ДНК с тяжелым изотопом N15 и ДНК с легким изотопом N14.
Эта промежуточная фракция стала для Мезельсона и Сталя свидетельством того, что молекулы ДНК, полученные на первом этапе репликации, являются гибридом лёгкой и тяжелой ДНК. В следствие чего этот результат соответствует дисперсионной и полуконсервативной моделям, но не соответствует консервативной модели.
Консервативная модель предсказала бы две отдельные фракции в этом поколении (фракция для тяжелой исходной молекулы ДНК и фракция для легкой новой молекулы ДНК).
Поколение 2
Информация о втором поколении позволила Мезельсону и Сталю определить, какая из оставшихся моделей (полуконсервативная или дисперсионная) была действительно правильной.
После того, как ДНК бактерий второго поколения была центрифугирована, она показала две фракции. Первая фракция находилась в той же позиции, что и промежуточная фракция из первого поколения, а вторая была выше (указывая на то, что ДНК отмечена только легким изотопом N14.
Полученный Мезельсоном и Сталем результат подтвердил, что ДНК копировалась полуконсервативно. Две различные градиентные фракции — гибридной молекулы и лёгкой молекулы — можно ожидать именно при полуконсервативной репликации (как показано на диаграмме ниже). Напротив, при дисперсионной репликации все молекулы должны иметь фрагменты как старой так и новой ДНК, что делает невозможным получение чистых «легких» молекул.
Поколения 3 и 4
В полуконсервативной модели ожидается, что каждая гибридная молекула ДНК из второго поколения будет давать одну гибридную молекулу и одну легкую молекулу в третьем поколении, тогда как каждая легкая молекула ДНК будет давать при репликации только легкие молекулы.
Таким образом, в третьем и четвертом поколениях мы ожидаем, что гибридная фракция будет становиться всё более узкой (поскольку тогда она представляет меньшую долю всей ДНК), а вторая фракция будет становиться всё шире (так как она представляет всё большую долю).
Дайте общую характеристику процесса репликации ДНК.
Каковы основные различия в репликации ДНК у про- и эукариот? Перечислите ферменты репликации ДНК и их функции.
Репликация ДНК – это молекулярный процесс точного копирования молекул ДНК (ее нуклеотидной последовательности). С помощью механизма репликации происходит точная передача генетической информации от клетки к клетке и, таким образом, все клетки многоклеточного организма являются носителями одной и той же наследственной информации. Процесс синтеза ДНК сопровождается множеством событий и является, как правило, точным. Из одной молекулы ДНК синтезируются две идентичные дочерние молекулы. Этот процесс становится возможным благодаря структурным особенностям молекулы ДНК: - двухцепочечная структура; - комплементарность и антипараллельность. Основными характеристиками репликации являются:
- синтез ДНК является полуконсервативным, так как каждая цепь служит матрицей для синтеза дочерней цепи;
- репликация носит двунаправленный характер; -
- синтез новой цепи осуществляется только в направлении 5'→3'; -
- в репликации участвует значительное количество белковых факторов.
Отличия репликации у эукариот связаны с нуклеосомным строением хромосом. ДНК наматывается на частички нуклеосом и межнуклеосомные фрагменты ДНК включают примерно 200 нуклеотидов. Поэтому фрагменты Оказаки у эукариот короче, чем у прокариот и составляют - 100-200 нуклеотидов. В связи с нуклеосомным строением меньше и скорость синтеза ДНК (50-100 нуклеотидов в сек). Нуклеосомы перед репликацией распадаются и формируются заново на новых цепях на расстоянии 200-400 нуклеотидов от репликативной вилки. Репликация идет в двух взаимопротивоположных направлениях с множеством репликонов. У эукариот не 1 репликон, как у прокариот, а множество: у дрожжей – 500, у млекопитающих – 20-30 тыс.
Точка начала репликации представлена специфичной последовательностью нуклеотидов, которая называется автономной последовательностью репликации, точкой ori. Участок ДНК который содержит точку ori который способен к независимой репликации называется репликон. У прокариот каждая молекула ДНК представляет один репликон, у эукариот ДНК содержит больше точек начала (οri), и соответственно больше репликонов.
В репликации ДНК участвует большое количество белковых факторов и ферментов.
ДНК-геликазы осуществляют деспирализацию и денатурацию участка ДНК с использованием энергии гидролиза АТФ и образование репликационной вилки. В связи с наличием двух репликационных вилок, существуют две геликазы, которые передвигаются в разных направлениях от точки инициации репликации.
Праймаза – фермент, который инициирует синтез ДНК за счет синтеза небольшого фрагмента, состоящего из 11-12 рибонуклеотидов, - праймера или затравки (РНКпраймер). Геликаза совместно с праймазой образует комплекс - праймосома.
Топоизомераза I разрывает только одну из двух цепей двойной спирали ДНК, расщепляя фосфодиэфирные связи, что предупреждает суперспирализацию молекулы ДНК. Топоизомераза II ковалентно связывается с обеими цепями двойной спирали ДНК и вносит в нее на время двухцепочечный разрыв.
Белки, связывающиеся с одноцепочечной ДНК (SSB – белки), это факторы, стабилизирующие одноцепочечную ДНК в области денатурации и препятствующие комплементарному спариванию двух цепей или различных участков одной и той же цепи, имеющих палиндромные последовательности
ДНК-полимеразы - ферменты, способные синтезировать новые цепи ДНК с матричной цепи. Было обнаружено три класса полимераз у прокариот (I, II, III) и полимеразы α, β, γ, δ, ε у эукариот.
ДНК-лигаза – это фермент, связывающий участки вновь синтезированной ДНК, посредством образования фосфодиэфирных связей 3'→5'.
Отдельные этапы репликации ДНК: инициация, элонгация, терминация.
Инициация включает следующие процессы:
присоединение реплисомы к точке инициации репликации и деспирализация участка двойной спирали ДНК геликазами;
синтез РНК-праймеров ферментом праймазой, выполняющей функции РНК-полимеразы;
присоединение дезоксирибонуклеотидов, комплементарных матрице, к 3'-концу праймера ДНК-полимеразой.
ДНК-гираза в точки оri гидролизует фосфодиэфирную связь → образование репликативной вилки → образование репликативного глазка с помощью ДНК-хеликазы.
SSB-белки стабилизируют репликативную вилку → ДНК-праймаза строит праймер → ДНК-полимераза достраивает нить
Элонгация характеризуется удлинением вновь синтезируемых цепей, которое осуществляется ДНКполимеразой, входящей в состав реплисомы, во время ее продвижения по матрице:
непрерывный рост лидирующей цепи;
прерывистый синтез фрагментов Оказаки на отстающей цепи;
контроль ошибок спаривания оснований во время репликации и их исправление при помощи 3'→5' экзонуклеазы из состава ДНК-полимеразы.
ДНК-полимераза продолжает удлинять нить из дезоксирибонуклеотидов на лидирующей нити, а также строить фрагменты фрагменты Оказаки на отстающей нити ДНК по мере движения репликативной вилки.
3. Терминация включает следующие процессы:
удаление РНК-праймеров 5'→3' экзонуклеазой, представляющей собой субъединицу ДНК-полимеразы;
заполнение недостающих участков ДНК-полимеразой;
связывание фрагментов вновь синтезированной ДНК с помощью лигаз.
ДНК-полимераза II удаляет праймеры и достраивает фрагменты ДНК → ДНК-лигаза соединяет фрагменты между собой.
Термины: репликативная вилка, репликативный глазок, репликоп, точка ori, праймеры, отстающая цепь, лидирующая цепь, фрагменты Оказаки.
Репликационная вилка (репликативная вилка) — Y-образная структура, перемещающаяся вдоль родительской спирали ДНК и характеризующаяся местным расхождением двух её цепей, в пределах которой происходит активная репликация ДНК.
Репликационные вилки образуются не в любых местах ДНК, а только в точках начала репликации, состоящих из определенной последовательности нуклеотидов (около 300 штук). Такие места распознаются специальными белками, после чего образуется так называемый репликационный глаз, в котором расходятся две цепи ДНК.
Репликон — единица репликации ДНК, от точки ее начала и до точки ее окончания.
Точки ori (англ. оrigin – начало) = точки начала репликации, в формируются репликативные вилки (100 – 1000 и больше)
Праймер — короткий фрагмент нуклеиновой кислоты или связана молекула, служит исходным пунктом репликации ДНК. Праймер нужен потому, что ни одна ДНК-полимераза (фермент, который катализирует репликацию ДНК) не может начать синтез новой молекулы ДНК с одноцепочечной матрице, поскольку нужна двухцепочечная участок для присоединения к ДНК, а также они способны только присоединять нуклеотид к уже имеющейся — ОН группы на 3 'конце другого нуклеотида.
Одна новая цепь, которая проходит от 5' к 3'-концу по направлению к вилке репликации, является простой. Эта цепь создается непрерывно, потому что ДНК-полимераза движется в том же направлении, что и вилка репликации. Эта непрерывно синтезируемая цепь называется лидирующей цепью.
В другой новой цепи, которая направлена от репликационной вилки от 5'-конца к 3'-концу, всё немного сложнее. Эта цепь строится фрагментами, потому что по мере движения вилки ДНК-полимераза удаляется от неё, и ей в какой-то момент приходится отрываться и снова присоединяться к новому открывшемуся фрагменту. Эта цепь, достраивающаяся фрагментами, называется отстающей цепью.
Фрагменты Оказаки (англ. Okazaki fragments) — относительно короткие фрагменты ДНК (с РНК-праймером на 5'-конце), которые образуются на отстающей цепи в процессе репликации ДНК. Лидирующая цепь может быть синтезирована на основании одного праймера, в то время как отстающая цепь нуждается в новом праймере для каждого из коротких фрагментов Оказаки.
Термины «экспрессия генов», понятия «ген» и «оперон». Каковы различия экпрессии генов у про- и эукариот.
Экспрессия генов — это процесс, в ходе которого наследственная информация от гена (последовательности нуклеотидов ДНК) преобразуется в функциональный продукт — РНК или белок.
Ген — структурная и функциональная единица наследственности, контролирующая развитие определённого признака или свойства.
Оперон — функциональная единица генома у прокариот , в состав которой входят цистроны ( гены , единицы транскрипции), кодирующие совместно или последовательно работающие белки и объединенные под одним (или несколькими) промоторами . Такая функциональная организация позволяет эффективнее регулировать транскрипцию этих генов.
Главное отличие между прокариотической и эукариотической экспрессией генов заключается в том, что полная экспрессия прокариотических генов происходит в цитоплазме, тогда как часть экспрессии эукариотических генов происходит внутри ядра, тогда как покой происходит в цитоплазме. Кроме того, регуляция экспрессии прокариотических генов в основном происходит на уровне транскрипции, тогда как регуляция экспрессии эукариотических генов происходит на различных этапах экспрессии генов.
Центральная догма молекулярной биологии. Какие существуют специальные способы передачи генетической информации в клетке?
Мы говорим о конце 50-х годов, более точно — о 1958 годе, когда Крик сформулировал эту центральную догму.
Таким образом, центральная догма — это постулат, что в живой клетке происходит направленный поток информации: ДНК — РНК — белок. Крик подчеркивал, что его центральная догма — это не позитивное, а негативное утверждение, и главное содержание центральной догмы состоит в том, что не происходит обратного потока информации, белок не может изменить генетическую информацию.
В этом состоит основной смысл центральной догмы: белок не в состоянии изменить информацию в РНК или в ДНК, поток идет в одну сторону.
Помимо общих способов передачи генетической информации выделяются специальные , начну с обратной транскрипции)
Итак,. Обратная транскрипция — это процесс образования двуцепочечной ДНК на матрице одноцепочечной РНК. Данный процесс называется обратной транскрипцией, так передача генетической информации при этом происходит в «обратном» направлении, относительно транскрипции.
Идея обратной транскрипции вначале была очень непопулярная, так как противоречила центральной догме молекулярной биологии, которая предполагала, что ДНК транскрибируется в РНК и далее транслируется в белки.
Однако, в 1970 году, en:Howard Temin и en:David Baltimore независимо друг от друга открыли фермент, названный обратной транскриптазой (ревертазой), и возможность обратной транскрипции была окончательно принята.
Репликация РНК — копирование цепи РНК на комплементарную ей цепь РНК с помощью фермента РНК-зависимой РНК-полимеразы. Таким способом реплицируются вирусы, содержащие одноцепочечную (например, пикорнавирусы, к которым относится вирус ящура) или двуцепочечную РНК.
Прямая трансляция белка на матрице ДНК (ДНК — белок)
Прямая трансляция была продемонстрирована в клеточных экстрактах кишечной палочки, которые содержали рибосомы, но не иРНК. Такие экстракты синтезировали белки с введённых в систему ДНК, и антибиотик неомицин усиливал этот эффект.[3][4]
Она состоит в том, что в клетке имеется направленный поток информации от ДНК, которая представляет собой исходный генетический текст, состоящий из четырех букв: A, T, G и C. Этот текст записан в виде последовательностей этих букв, которые называются нуклеотидами, в двойной спирали ДНК, в одной из двух цепей. Этот текст транскрибируется. Процесс называется транскрипцией. Синтезируется РНК, которая идентична этому тексту, с той разницей, что в РНК вместо T стоит U, но они очень похожи, и РНКовый текст является слепком с соответствующего ДНКового текста.
Эта РНК, которая называется информационной РНК, или мРНК, транслируется с помощью генетического кода в виде последовательности аминокислотных остатков в белках — перевод текста нуклеиновых кислот ДНК и РНК из 4-буквенного текста в 20-буквенный текст аминокислот. Канонических аминокислот, или природных аминокислот, — 20, а букв в тексте нуклеиновых кислот — 4, поэтому происходит перевод из 4-буквенного алфавита в 20-буквенный с помощью генетического кода, в котором каждой тройке нуклеотидов отвечает какая-то аминокислота. Всего можно сделать из 4 букв 64 комбинации по 3 буквы, а аминокислот 20, поэтому код должен был быть вырожденным, и в то время он еще не был известен, даже не начали его расшифровывать, когда Крик сформулировал свою центральную догму. Но уверенность в том, что код должен существовать, уже была, и к тому времени уже было показано, что он триплетный, что именно 3 буквы в нуклеиновых кислотах отвечают одной аминокислоте. Эти 3 буквы, которые отвечают аминокислоте, получили название кодонов. Этих кодонов 64, а они кодируют 20 аминокислот. Значит, каждой аминокислоте — почти каждой — отвечает по несколько кодонов. Есть аминокислоты, которым отвечает только один кодон, но, как правило, больше чем один.
Перечислите виды РНК и их функции.
|
|
|
 Скачать 264.99 Kb.
Скачать 264.99 Kb.