Физиология как наука
 Скачать 0.79 Mb. Скачать 0.79 Mb.
|

Происхождение электрических явлений в тканяхНа уровне клетки регистрируется потенциал мембраны (ПД) - разность потенциалов между наружной и внутренней поверхности мембраны в каждый данный момент времени. Стационарно, как показатели электрического состояния клетки регистрируют 2 вида потенциала мембраны (ПМ): потенциал покоя (ПП) и потенциал действия (ПД). Потенциал покоя(ПП) - это разность потенциалов между наружной и внутренней поверхности мембраны в состоянии покоя, т.е. в покое мембрана поляризована. Если на клетку нанести раздражение достаточной силы, клетка придет в новое, активное состояние. Если силы раздражителя недостаточно, чтобы сместить ПМ до некого критического уровня, то происходит возращение ПМ к исходному уровню, т.е. к уровню ПП. Возникшие изменения ПМ называются- локальный ответ. Если силы раздражителя достаточно, чтобы сместить ПМ до критического уровня деполяризации, то произойдет формирования потенциала действия(ПД), что свидетельствует о возбуждении клетки переходе ее в деятельное состояние. Потенциал действия и потенциал покоя -это электрические явления, регистрируемые на уровне клетки. На уровне ткани регистрируются следующие биоэлектрические явления: В состоянии покоя: - токи покоя (повреждения) - (ТП), - токи градиента основного обмена (ТГОО). В состоянии возбуждения: - токи действия (ТД). 6. Понятие о потенциале покоя. Роль ионов К+, Na+, Ca+2, Cl- в происхождении мембранного потенциала. Калий-натриевый насос, его значение. Уравнения Нернста и Гольдмана, расчет величины мембранного потенциала. Потенциал покоя (ПП) - это разность потенциалов между наружной и внутренней поверхности мембраны в состоянии покоя, т.е. в покое мембрана поляризована. Происхождение ПП обусловлено: 1. Неравномерным распределением ионов калия и натрия между цитоплазмой и межклеточной жидкостью. В клетке - калия порядка 400 мкмоль/литр, вне клетки – 10, соответственно, натрия в клетке - 50 и 460 - вне клетки - в состоянии покоя. 2. Избирательная проницаемость клеточной мембраны в покое для натрия и калия. В покое - высокая проницаемость для калия, а для натрия в покое она практически отсутствует небольшая. В покое за счет процесса облегченной диффузии через неуправляемые медленные калиевые каналы за счет градиента концентрации - калий постоянно выходит из клетки во внеклеточное пространство, это формирует постоянный выходящий калиевый ток. Он является причиной разности потенциалов в покое и обуславливает ПП. Постоянному выходящему калиевому току противодействует работа калиевая часть калий-натриего насоса, которая обеспечивает постоянное возвратное поступление 2 молекул калия из внешней среды в клетку. В покое скорости этих двух процессов невелики. В реальных условиях в клетке возникает некое равновесное состояние между выходящим калиевым током и входящим калиевым током. Это формирует некий равновесный потенциал /ЕК/, который формирует по существу ту реальную разность потенциалов, которая существует между наружной и внутренней поверхностью клетки, если бы ее создавал один вид ионов. Его величина, описывается уравнением Нернста: 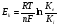 где: R- газовая постоянная, Т- абсолютная температура, F- число Фарадея, Ке - концентрация свободных ионов калия в наружном растворе, Кi - их концентрация в цитоплазме, n - валентность, ln - натуральный логарифм. По этой формуле Нернста можно подсчитать вклад калия в формирование потенциала мембраны в состоянии покоя, а так же - возбуждения. Реально равновесный потенциал для калия в покое равен минус 90 милливольт. Натрий, его равновесный потенциал в покое - плюс 60 милливольт. Хлор, для него равновесный потенциал равен - минус 70. Гольдман - американский ученый - создал объединительное уравнение, в котором суммировал эти показатели потенциалов для натрия, калия, хлора, только в эти уравнения введено одно добавление - коэффици ент проницаемости мембраны для иона, который характеризует скорость диффузии каждого из ионов. Расчет с использованием уравнения Гольдмана показывает, что в состоянии покоя потенциал мембраны составляет минус 70 милливольт. Реальные измерения ПП – 70-80 мВ. Таково электрическое состояние мембраны клетки в покое - она поляризована.
Если на клетку нанести раздражение достаточной силы, клетка придет в новое, активное состояние. При нанесении раздражения увеличивается проницаемость мембраны для натрия. Это происходит за счет энергии раздражителя и связано с активацией небольшого числа натриевых каналов. Возникает небольшое усиление входящего натриевого тока, интенсивность входящего натриевого тока пропорционально силе раздражителя. За счет этого процесса происходит уменьшение полярности мембраны по сравнению с исходным (с 70 мВ до 40-50 мВ). Эти изменения называются - пассивная деполяризация или частичная деполяризация, выраженность и скорость этого процесса зависит от силы раздражителя. Если силы раздражителя недостаточно, чтобы сместить ПМ до некого критического уровня, то происходит возращение ПМ к исходному уровню, т.е. к уровню ПП. Возникшие изменения ПМ называются - локальный ответ. Если силы раздражителя достаточно, чтобы сместить ПМ до критического уровня деполяризации, то произойдет формирования потенциала действия (ПД), что свидетельствует о возбуждении клетки переходе ее в деятельное состояние. Критический уровень деполяризации/КУД/- это такая величина разности потенциалов (40-50 мВ), при которой активируется большое количество потенциалзависимых быстрых натриевых каналов, проницаемость мембраны для натрия становится максимальной и перестает быть зависимой от силы раздражителя. Возникает лавинообразный входящий натриевый ток, который быстро (доли мс) смещает потенциал мембраны до 0 (активная деполяризация - потеря полярности), а затем его силы хватает чтобы изменить знак мембраны на противоположный - плюс 10-20 мВ. (Смена знака потенциала мембраны называется овершут или реверсия потенциала). Входящий натриевый ток формирует восходящую часть пика (спайка) потенциала действия, наличие которого указывает на сформировавшееся возбуждение клетки. Амплитуда пика не зависит от силы раздражителя - закон «все или ничего», будем рассматривать в следующей лекции. Наличие восходящей части пика ПД свидетельствует, что клетка перешла в новое функциональное состояние – состояние возбуждения, т.е. в деятельное состояние. Вторая половина ПД (нисходящая) состоит из трех частей: 1. Нисходящая часть пика ПД (от острия пика до КУД), формируется быстро (за доли мс), 2. Положительный следовой потенциал (от КУД до ПП) формируется медленнее (несколько мс), 3. Отрицательный следовой потенциал(несколько мс). 1 и 2 части обеспечиваются процессом реполяризации, 3 часть - процессом гиперполяризации. Процесс реполяризации - возвращение, восстановление полярности мембраны клеток, которое для них характерно в покое. Процесс реполяризации обусловлен: 1. активацией потенциалзависимых быстрых калиевых каналов, которая(активация) возникает при ПМ 0- плюс 5 мВ), что приводит к возникновению значительного по объему выходящего калиевого тока. 2. быстрой инактивацией потенциалзависимых натриевых каналов, которая возникает сразу после достижения высшего значения ПД (+10,+20мв). Это блокирует входящий натриевый ток. 3. значительной активацией калий-натриевого насоса (увеличение скорости оборота), которая обеспечивает удаление избытка натрия в клетке, возникшего в фазу деполяризации. Эти три процесса обеспечивают возвращение ПМ до уровня ПП. Следует заметить, что эти три процесса ионного транспорта инертны и не инактивируются мгновенно при достижении ПМ уровня ПП, что приводит к избыточному перемещению ионов и, как следствие, к избыточной поляризации мембраны (гиперполяризации), за счет которой (гиперполяризации) и формируется отрицательный следовой потенциал. Затем ПМ мембраны клетка приходит в исходное состояние. Следует иметь ввиду, что при формировании ПД выходящий калиевый ток, осуществляемый через неуправляемые медленные калиевые каналы по объему ничтожно мал по сравнению с объемом перемещения натрия и калия через быстрые потенциалзависимые каналы, так как процесс формирования ПД происходит за мс. ВАЖНО. Включение других потенциалзависимых каналов существенно влияет на форму и продолжительность потенциала действия. Так, активация долговременных потенциалзависимых кальциевых каналов (L-тип), их много в кардиомиоцитах, формирует фазу плато, клетка длительно деполяризована. Потенциал действия всегда, сформировавшись, распространяется на соседние участки клетки, локальный ответ остается там, где он возник: он не способен распространяться. 8. Раздражимость и возбудимость… Возбуждение - процесс, который возникает только при действии раздражителя. Раздражители - факторы, которые воздействуют на биологические клетки. Качественно (по природе) раздражители делятся на: - физические (электромагнитные волны, электрический ток, механические воздействия, температура), - химические. По биологической значимости: -адекватные (присущи для восприятия данному виду рецептора) -неадекватные (не являются естественными с точки зрения природы или силы раздражения). Все раздражители (по силе) делятся на: пороговые, подпороговые, надпороговые Порог раздражителя - та минимальная сила, при действии которой возникает возбуждение. Законы раздражения Действие раздражителя описывается несколькими законами: 1. Закон силы раздражения: Чем больше сила раздражения, тем, до известных пределов, сильнее ответная реакция. Раздражители имеют нижний предел - подпороговое раздражение не вызывает ответной реакции. Возбудимые ткани работают только на пороговых и надпороговых раздражителях. Но есть сила раздражения для любого биологического раздражителя, которая способна вызывать max эффект - оптимальная сила (оптимум частоты и силы раздражения). Если сила больше, чем оптимальная, то ответная реакция ниже - пессимум частоты или силы раздражения. 2. Закон длительности раздражения: Чем длительнее раздражение, необходимое для возникновения возбуждения, тем сильнее, до известных пределов, ответная реакция живых систем. Есть зависимость между силой раздражения и временем, в течение которого этот раздражитель должен действовать, чтобы вызвать ответную реакцию. Зависимость выражается гиперболой, следовательно, даже сильные раздражители, действуя кратковременно, либо не способны вызвать ответную реакцию, либо - слабую ответную реакцию и наоборот. График "сила-время" Особенно чётко зависимость между силой и временем в прослеживается в диапазоне промежуточных величин. 3. Закон градиента силы: Величина ответной реакции и её характер зависят ещё и от интенсивности/крутизны/ нарастания действия силы. Более интенсивное нарастание силы раздражения вызывает больший ответ. При этом длительное действие раздражителей одной и той же по величине силы, приводит к развитию аккомодации - явления, которое выражается в понижении чувствительности ткани к раздражению, уменьшению возбудимости ткани. Механизм этого явления бкдет рассмотрен в следующей лекции. 4. Закон "всё или ничего": Если раздражитель меньше пороговой силы, он никогда не вызовет ПД (потенциал действия) - "ничего". Но какой бы силы ни был надпороговый раздражитель, он всегда будет вызывать max для данного состояния электрическую реакцию, т.е. max пик ПД - "всё". Ответная реакция, её характер зависят от скорости химических процессов обеспечивающих ответные реакции, так называемые скорости активационных и инактивационных /восстановительных/ процессов. Введенский назвал свойство клеток, тканей, связанное со скорость активационных и инактивационных процессов - лабильность (функциональная подвижность)-свойство клетки, ткани, отражающее их максимальные возможности. Мера лабильности - это максимальная частота, которую способна воспроизвести ткань или клетка. Характеризуется способностью ткани отвечать ПД на каждое раздражение. У каждой ткани лабильность различна: в синапсах - 40-50 раз в сек., в периферических нервах - до 20000 раз в сек. Если лабильность ткани превышена, то ткань отвечает либо снижением ответной реакции, либо, если Вы долго будете принуждать ткань работать в режиме большем, чем лабильность - гибелью (это своего рода защитная реакция). Вот почему раздражение выше - по силе или по частоте - чем то, которое вызывает максимальный ответ - вызывает снижение ответа - пессимум (то, о чем мы говорили чуть раньше - при разборе закона силы - вот почему сверхсильные раздражители не дают сверхсильной реакции - они дают в здоровом организме снижение эффекта - это своеобразная защитная реакция). Частота раздражения близкая или совпадающая с величиной лабильности вызывает максимальный ответ, т.е. является оптимальной/ оптимум частоты раздражения/ ВОЗБУДИМОСТЬ Под возбудимостью понимают способность отвечать на раздражение формированием электрической активности /потенциалом действия/. У различных тканей возбудимость различна. У одной ткани возбудимость может изменяться в процессе жизнедеятельности, возбудимость у живой возбудимой ткани есть всегда, в не зависимости от действия раздражителя. Возбуждение это состояние, это реализованная возбудимость. Меры возбудимости. Для оценки возбудимости в каждой лаборатории функциональной диагностики существует специальный аппарат, называемый хронаксиметром (от слова - “хронос” - время). Это - прибор, который позволяет оценить возбудимость. Итак, к мерам возбудимости относятся: 1. Порог раздражения - первая базисная мера раздражителя любой природы. Порог раздражения - см. выше. Но для количественной оценки возбудимости в медицине используют не любой раздражитель, а используют электрический ток. Именно с помощью электрического тока тестируют мышцы, нервы, синапсы. Электрический ток точно дозируется - электрический ток можно легко дозировать, при чем по двум показателям: по силе и по времени действия. С другими раздражителями иначе: например, химический - можно дозировать по силе (концентрации), но нельзя - по длительности, так как для его отмывания нужно время. С помощью электрического тока получены еще 3 меры возбудимости, одна из которых используется в медицине. 1. Базисная мера - это реобаза. Это - минимальная сила постоянного тока, которая, действуя длительное, но определенное время, способна вызвать ответную реакцию. Недостаток этой меры - определение времени трудно определимо - оно расплывчато. 2. Полезное время - то время, которое должна действовать сила тока в 1 реобазу, чтобы вызвать ответную реакцию. Но и эта мера возбудимости не нашла своего применения в медицинской практике, потому что, как показывает график, она находится на очень пологой части кривой "сила - время" и любая неточность (небольшая неточность) вела к большой ошибке. 3. Поэтому в практику была введена еще одна мера - хронаксия. Это - минимальное время, в течение которого должна действовать сила тока в 2 реобазы, чтобы вызвать ответную реакцию. На графике - это тот участок кривой, где зависимость между силой и временем точно прослеживается. Посредством хронаксии определяют возбудимость нервов, мышц, синапсов. Этим методом определяют, где же наступило поражение нервно-мышечной системы: на уровне мышцы, нервов, синапсов или центральных образований. Нормальная возбудимость в покое принимается за 100 %. Возбудимость характеризуется разностью между потенциалом мембраны и КУДом. Период начального изменения возбудимости при формировании ПД называется периодом супернормальной возбудимости. В момент достижения КУД наступает максимальная проницаемость мембраны для натрия. В этот момент натрий потоком идет в клетку. Если в момент пика нанести новое раздражение на клетку, то клетка на нее не ответит, каким бы сильным раздражителем не пользовались. Натрий потоком идет в клетку, и нет таких сил, чтобы это остановить, выкачать натрий из клетки и снова его закачать. В этот момент возбудимость у клетки будет равна нулю (фаза абсолютной рефрактерности). По мере реполяризации будет происходить процесс восстановления возбудимости. Это называется фазой относительной рефрактерности (клетку могут возбудить только чрезвычайно сильные раздражители). Отрицательный следовой потенциал обусловлен гиперполяризацией мембраны. А раз поляризация избыточна, то возбудимость будет пониженной - это фаза субнормальной возбудимости (ниже нормальной возбудимости). |