Физиология. Лек 2 физиология бактерий лекция. Физиология микроорганизмов Микроорганизмам, как и всем живым существам, присущи процессы питания, дыхания, роста и размножения
 Скачать 0.53 Mb. Скачать 0.53 Mb.
|

|
Ферменты бактерий. В бактериальной клетке происходят многочисленные реакции, как биосинтетические, направленные на синтез соединений, необходимых для организации структуры бактерии, так и производящие энергию, процессы ассимиляции и диссимиляции. Все эти реакции катализируются соответствующими ферментами. Ферменты являются белками и обладают специфичностью при распознавании соответствующего вещества и последующем превращении его. Большая часть ферментов связана с определенными структурами бактериальной клетки. Так, в цитоплазматической мембране находятся окислительно-восстановительные ферменты, которым принадлежит основная роль в дыхании клетки, ферменты, обеспечивающие доставку питательных веществ, и др. Ферменты, связанные с делением клетки, обнаруживаются в мезосомах, клеточной стенке, в месте образования перегородки. У бактерий по характеру вызываемых ими превращений обнаруживаются следующие основные группы ферментов: г и д р о л а з ы, вызывающие расщепление протеинов, углеводов, липидов путем присоединения молекул воды; оксидоредуктазы, катализирующие окислительно-восстановительные реакции; трансфера з ы, осуществляющие перенос отдельных атомов, от молекулы к молекуле; л и а з ы, отщепляющие химические группы негидролитическим путем; изомеразы, участвующие в углеводном обмене; л и г а з ы, способствующие биосинтетическим реакциям клетки. Ферменты бактерий классифицируются на экзоферменты и эндоферменты. Экзоферменты выделяются бактериальной клеткой в окружающую среду для внеклеточного переваривания. Этот процесс осуществляется с помощью гидролаз, которые расщепляют макромолекулы питательных веществ до простых соединений — глюкозы, аминокислот, жирных кислот. Такие соединения могут свободно проходить через оболочку клетки и с помощью пермеаз передаваться в цитоплазму клетки для участия в метаболизме, являясь источниками углерода и энергии. Некоторые экзоферменты выполняют защитную функцию, например, пенициллиназа, выделяемая многими бактериями, делает клетку недосягаемой для антибиотика — пенициллина. Эндоферменты катализируют метаболические реакции, происходящие внутри клетки. Ферменты бактерий классифицируются также на конститутивные и индуцибельные. Конститутивными называются такие ферменты, которые синтезируются клеткой независимо от наличия субстрата в среде, индуцибельные ферменты образуются бактериями только при наличии в среде соответствующего индуцирующего соединения, т. е. субстрата данного фермента. Например, в геноме кишечной палочки заложена способность разлагать лактозу, но только при наличии в среде лактозы клеткой синтезируется фермент, катализирующий ее гидролиз. Известны также ферменты, которые получили название аллостерических . Кроме активного центра у них имеется регуляторный или аллостерический центр, который в молекуле фермента пространственно разделен с активным центром. Аллостерическим (от греч. allos - иной, чужой) он называется потому, что молекулы, связывающиеся с этим центром, по строению ( стерически ) не похожи на субстрат, но оказывают влияние на связывание и превращение субстрата в активном центре, изменяя его конфигурацию. Молекула фермента может иметь несколько аллостерических центров. Вещества, связывающиеся с аллостерическим центром, называют аллостерическими эффекторами. Они влияют через аллостерический центр на функцию активного центра: или облегчают ее, или затрудняют. Соответственно аллостерические эффекторы называются положительными (активаторы) или отрицательными (ингибиторы). Аллостерические ферменты играют важную роль в тонкой регуляции метаболизма бактерий. Поскольку практически все реакции в клетке катализируются ферментами, регуляция метаболизма сводится к регуляции интенсивности ферментативных реакций. Патогенные бактерии обладают наряду с ферментами обмена также ферментами агрессии, являющимися факторами вирулентности. К таким ферментам относятся гиалуронидаза, дезоксирибонуклеаза, коллагеназа, н е й р а м и и и д аза, и др. Гиалуронидаза стрептококков, например, расщепляет гиалуроновую кислоту в мембранах клеток соединительных тканей макроорганизма , что способствует распространению возбудителей и их токсинов в организме, обуславливая высокую инвазивность этих бактерий. Плазмокоагулаза является главным фактором патогенности стафилококков, так как участвует в превращении протромбина в тромбин, который вызывает образование фибриногена, в результате чего каждая бактерия покрывается пленкой, предохраняющей ее от фагоцитоза. Ферменты бактерий обладают высокой специфичностью, и именно это свойство широко используется при идентификации и определении видов микроорганизмов. Наибольшее значение имеет определение сахаролитических (ферментация сахаров) и протеолитических (разложение белков) свойств. Споры Одной из особенностей микроорганизмов является их способность к спорообразованию. Споры образуются при неблагоприятных условиях существования (высушивание, недостаток питательных веществ, изменение рН среды и т. д.), причем из одной клетки формируется только одна спора. Таким образом, образование спор не связано с процессом размножения, а является своеобразным приспособлением к переживанию в неблагоприятных условиях. По принятой номенклатуре спорообразующие аэробы носят название бацилл, а спорообразующие анаэробы — клостридии. Процесс спорообразования проходит ряд стадий, в течение которых в определенном месте клетки цитоплазма, нуклеоид, рибосомы концентрируются, уплотняются, покрываются мембраной, а затем плотной, плохо проницаемой многослойной оболочкой, включающей кальциевые соли дипиколиновой кислоты, обусловливающей термоустойчивость спор. Споры длительное время могут сохраняться в покое, оставаясь жизнеспособными. Так, в почве споры патогенных микроорганизмов (возбудителя сибирской язвы, столбняка и др.) могут сохраняться десятками лет. При попадании в благоприятную среду споры очень быстро прорастают — из 1 споры возникает 1 бактериальная клетка, которая начинает размножаться. Спорообразование — видовое свойство палочек, а форма и расположение формирующейся споры по отношению к вегетативной части клетки является дифференциально-диагностическим признаком. Форма спор может быть овальной или круглой, расположение центральное (возбудитель сибирской язвы), субтерминальное — ближе к концу палочки (возбудители газовой гангрены, ботулизма) и терминальное — на конце (возбудитель столбняка).  В зрелой споре различимы: центральный, плохо окрашиваемый участок (спороплазма), двухслойная ЦПМ и оболочка споры. 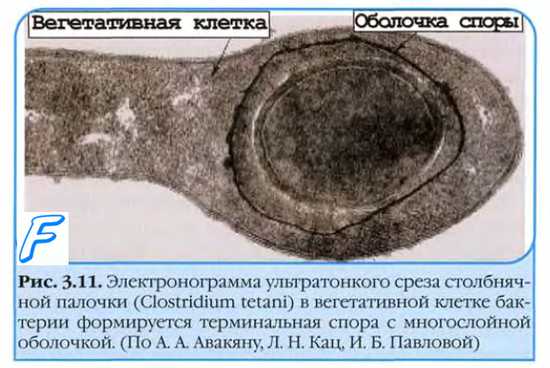 Спороплазма (протопласт споры) включает цитоплазму, бактериальную хромосому, системы белкового синтеза и некоторые другие (например, анаэробного энергообразования). Оболочка споры двухслойная: пространство между слоями заполняют гликопептидные полимеры, сходные с пептидогликанами, образующие сетчатую структуру (кортекс), проявляющую высокую чувствительность к лизоциму. Внутренний слой (стенка споры) образован пептидогликанами, аналогичными таковым \ вегетирующей клетки. Внешний слой (собственно оболочка) образуют кератиноподобные белковые структуры с низкой проницаемостью. Споруляция бактерий. Стадии споруляции у бактерий. Расположение спор у бактерий. Процесс споруляции (спорообразования) начинается сразу после возникновения дефицита питательных веществ и продолжается приблизительно 8 ч. Никаких внешних источников питания или энергии при этом не требуется. Споруляцию стимулирует внесение в среду глюкозы, фосфора и NH4; угнетает внесение пептона, лактозы, NaCl, CaCl2 (у бактерий рода Bacillus— DL-аланина). Споруляция контролируется особыми генами. Их число вариабельно у различных видов и может достигать 70. Детали спорообразования служат видовыми признаками, но его принципиальные закономерности одинаковы для всех бактерий. Подготовительная стадия споруляции сопровождается прекращением деления и увеличением количества липидных включений. Стадия предспоры споруляции обычно начинается бурно. В клетке появляется эллиптическая оболочка, окружающая участок цитоплазмы с изменёнными плотностью и тинкториальными свойствами. Подобное образование обозначают терминами «предспора», или «примордиальная спора». Третья стадия споруляции включает появление оболочки (обычно в течение 10 мин после образования предспоры) и ещё большее увеличение коэффициента светопреломления. Стадия созревания споры сопровождается её уплотнением и снижением метаболической активности клетки. Поскольку споры сильно преломляют свет, то распознавание их при микроскопии не представляет затруднений, В спорных случаях прибегают к специальным методам окрашивания. Прорастание спор бактерий. Активация споры. Покоящиеся ( некультивируемые ) формы бактерий. После созревания и лизиса родительской клетки спора освобождается, сохраняясь в покоящемся состоянии до попадания в благоприятные условия. • Прорастание споры в оптимальных условиях осуществляется в течение 2-3 ч; процент проросших спор увеличивается после соответствующей предварительной обработки. Например, споры могут быть активированы кратковременным прогреванием. Споры кислотоустойчивы, поэтому окрашиваются по методу Ожешки или по методу Циля-Нильсена в красный, а вегетативная клетка в синий цвет. Спора долго сохраняется из-за наличия многослойной оболочки, дипиколината кальция, низкого содержания воды и вялых процессов метаболизмов. В благоприятных условиях споры прорастают, проходя три последовательные стадии: активация, инициация, прорастание. • Прорастанию предшествует активация споры. Её инициируют различные химические вещества, повышение температуры и влажности. Под воздействием автолизинов происходит расщепление кортекса, поглощение воды и набухание. Внешне процесс проявляется увеличением («вздутием») споры и уменьшением коэффициента светопреломления. При этом в споре происходят глубокие физиологические изменения: усиливается дыхание, увеличивается активность ферментов, происходит выделение аминокислот, дипиколиновой кислоты и пептидов (потеря сухой массы споры может достигать 20-30%). В этот период спора утрачивает терморезистентность. Затем спора лопается в произвольном месте и из неё выходит вегетативная клетка, снабжённая у подвижных видов жгутиковым аппаратом. Покоящиеся ( некультивируемые ) формы бактерий Специализированные анабиотические формы (такие как эндо- и экзоспоры, миксоспоры, цисты) известны лишь для ограниченного круга микроорганизмов. Однако как у спорообразующих микроорганизмов (Bacillus cereus) на фоне подавления спорообразования, так и у споронеобразующих (Pseudomonas carboxidoflava, Micrococcus luteus, Escherichia coli) бактерий внеклеточные микробные метаболиты, проявляющие свойства индукторов анабиоза, вызывают образование цистоподобных рефрактерных клеток (термин связан с переходом целой клетки в покоящуюся форму с увеличенной способностью данных клеток по сравнению с обычными сильно преломлять свет). В этих клетках снижение метаболической активности (гипометаболизм) сопровождается развитием резистентности к экстремальным воздействиям. Эти формы отличаются нерегистрируемым уровнем эндогенного дыхания, повышенной терморезистентностью и специфической ультраструктурой. По сравнению с вегетативными клетками, цистоподобные рефрактерные клетки обладают рядом особенностей ультраструктурной организации: у них снижена плотность рибосом в цитоплазме, цитоплазма приобретает мелкозернистую структуру, увеличена толщина клеточной стенки, в самой клеточной стенке появляются слоистость и некоторые плотные включения. Для споронеобразующих микроорганизмов цистоподобные рефрактерные клетки — единственная покоящаяся форма, способная сохраняться в течение нескольких лет, а для спорообразующих бактерий — альтернатива спорообразования. Рефрактерные клетки невозможно выявить с помощью традиционных бактериологических подходов. Капсула микроорганизмов Капсула возникает благодаря способности ряда бактерий (как сапрофитов, так и патогенных) создавать снаружи клеточной стенки скопление слизистого вещества. Образование капсулы часто является признаком вирулентности для патогенных бактерий, так как она позволяет клетке противостоять воздействию защитных механизмов макроорганизма. Такие бактерии образуют капсулу в организме (например, пневмококк, возбудитель сибирской язвы) и утрачивают это свойство при культивировании на питательных средах. Группа патогенных микроорганизмов, получивших название «капсульные бактерии», образует капсулу независимо от условий существования: и в организме, и при культивировании на питательных средах. Химический состав капсул различен: полисахариды у пневмококков, преимущественно полипептиды у возбудителя чумы. При микроскопии препаратов, содержащих бактерии в капсулах, последние имеют вид бесцветного ореола, окружающего клетки, так как вещество капсул плохо воспринимает красители. Микроорганизмы, у которых не выявляется четкой капсулы, могут содержать аналогичную структуру, обнаруживаемую в виде очень тонкого поверхностного слоя, получившего название микрокапсулы. Микрокапсула хорошо видна под электронным микроскопом. В чистых культурах бактерий капсула образуется реже. Она выявляется при специальных методах окраски мазка (например, по Бурри-Гинсу), создающих негативное контрастирование веществ капсулы: тушь создает темный фон вокруг капсулы. Капсула гидрофильна, препятствует фагоцитозу бактерий. Капсула антигенна: антитела против капсулы вызывают ее увеличение (реакция набухания капсулы). От капсулы следует отличать слизь - мукоидные экзополисахариды, не имеющие четких границ. Слизь растворима в воде. Бактериальные экзополисахариды участвуют в адгезии (прилипании к субстратам), их еще называют гликокаликсом. Культуральные свойства бактерий К культуральным или макроморфологическим свойствам относятся характерные особенности роста микроорганизмов на плотных и жидких питательных средах. На поверхности плотных питательных сред в зависимости от посева микроорганизмы могут расти в виде колоний, штриха или сплошного газона. Колонией называют изолированное скопление клеток одного вида, выросших из одной клетки (клон клеток). В зависимости от того, где растет микроорганизм (на поверхности плотной питательной среды, в толще ее), различают поверхностные, глубинные и донные колонии. Колонии, выросшие на поверхности среды, отличаются разнообразием, они видоспецифичны и их изучение используется для определения видовой принадлежности исследуемой культуры. При описании колоний учитывают следующие признаки: форму колонии - округлая, амебовидная, ризоидная , неправильная и т.д.; размер (диаметр) колонии - очень мелкие (точечные) (0,1-0,5 мм), мелкие (0,5-3 мм), средних размеров (3-5 мм) и крупные (более 5 мм в диаметре); поверхность колонии - гладкая, шероховатая, складчатая, морщинистая, с концентрическими кругами или радиально исчерченная; профиль колонии - плоский, выпуклый, конусовидный, кратерообразный и т.д.; прозрачность - тусклая, матовая, блестящая, прозрачная, мучнистая; цвет колонии (пигмент) - бесцветная или пигментированная (белая, желтая, золотистая, красная, черная), особо отмечают выделение пигмента в среду с ее окрашиванием; край колонии - ровный, волнистый, зубчатый, бахромчатый и т.д.; структура колонии - однородная, мелко или крупнозернистая, струйчатая; край и структуру колонии определяют с помощью лупы или на малом увеличении микроскопа, поместив чашку Петри с посевом на столик микроскопа крышкой вниз; консистенция колонии - определяют прикасаясь к поверхности петлей, колония может быть плотной, мягкой, врастающей в агар , слизистой (тянется за петлей), хрупкой (легко ломается при соприкосновении с петлей). Глубинные колонии чаще всего похожи на более или менее сплющенные чечевички (форма овалов с заостренными концами), иногда комочки ваты с нитевидными выростами в питательную среду. Образование глубинных колоний часто сопровождается разрывом плотной среды, если микроорганизмы выделяют газ. Донные колонии имеют обычно вид тонких прозрачных пленок, стелющихся по дну. Описание роста микроорганизмов при посеве штрихом включает его особенности: скудный, умеренный, обильный; сплошной с ровным или волнистым краем; диффузный; перистый; ризоидный ; древовидный. Характеризуют цвет, поверхность, консистенцию. Особенности колонии могут изменяться с возрастом, они зависят от состава среды, температуры культивирования. Рост микроорганизмов на жидких питательных средах учитывают, используя 4-7 суточные культуры, выращенные в стационарных условиях. В жидких питательных средах при росте микроорганизмов наблюдается помутнение среды, образование пленки или осадка. Рост бактерий с равномерным помутнением среды , что характерно для факультативных анаэробов. Степень помутнения может быть слабая, умеренная, сильная. Придонный рост бактерий характеризуется образованием осадка: скудного, обильного, рыхлого, слизистого, хлопьевидного, зернистого. Питательная среда может быть прозрачной или мутной. Пристеночный рост - образование зерен, рыхлых хлопьев на внутренней поверхности стенок сосуда. Питательная среда при этом остается прозрачной. Поверхностный рост бактерий характеризуется появлением на поверхности среды пленки: тонкой, плотной, рыхлой, гладкой, складчатой, влажной, сухой, кольцеобразной, сплошной. Такой рост наблюдается при культивирован ии аэ робных бактерий. При росте на полужидких (0,5-0,7% агара ) питательных средах подвижные микробы вызывают выраженное помутнение, неподвижные формы растут только по ходу посева уколом в среду. Нередко рост микробов сопровождается появлением запаха, пигментацией среды, выделением газа. Характерный запах культур некоторых видов бактерий связан с образованием различных эфиров ( уксусноэтилового , уксусноамилового и др.), индола, меркаптана, сероводорода, скатола, аммиака, масляной кислоты. Способность образовывать пигменты присуща многим видам микроорганизмов. Химическая природа пигментов разнообразна: каротиноиды , антоцианы, меланины. Если пигмент не растворим в воде, окрашивается только культуральный налет, если же он растворим, окрашивается и питательная среда. Считается, что пигменты защищают бактерии от губительного действия солнечных лучей, поэтому в воздухе так много пигментированных бактерий, кроме того, пигменты участвуют в обмене веществ этих микроорганизмов. В природе существуют, так называемые, фосфоресцирующие бактерии, культуры которых светятся в темноте зеленовато-голубоватым или желтоватым светом. Такие бактерии встречаются, главным образом, в речной или морской воде. К светящимся бактериям - фотобактериям - относятся аэробные бактерии (вибрионы, кокки, палочки). |