методичка Физиология почки и ВСО. Физиология почки и водносолевого обмена
 Скачать 1.96 Mb. Скачать 1.96 Mb.
|

Кровоснабжение почкиКровоснабжение в почке играет особую роль, поскольку не только обеспечивает клеточный метаболизм, но и принимает непосредственное участие в мочеобразовании. В 1 минуту через сосуды обеих почек у человека проходит около 1200 мл крови, т.е. около 20-25% крови, выбрасываемой сердцем в аорту. Так как масса почек у человека составляет всего лишь 0,43% массы тела, очевиден исключительно высокий уровень органного кровотока (рис.3) Величина почечного плазмотока и кровотока определяется методом очищения по ПАГ (руководство к проведению лабораторных работ). Через сосуды коры почки протекает 91-93% крови, поступающей в почку, остальное ее количество снабжает мозговое вещество почки. Кровоток в коре почки в норме составляет 4-5 мл/г ткани. Важной особенностью почечного кровотока является высокий уровень саморегуляции – кровоток остается постоянным при изменении артериального давления более, чем в два раза (например, с 90 до 190 мм рт.ст.). 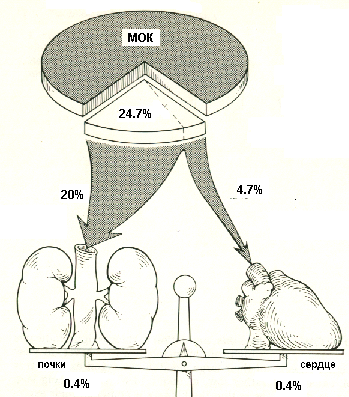 Рисунок 3 Сравнение почечного и коронарного кровотока Артерии почки отходят от брюшного отдела аорты, что обеспечивает высокий уровень артериального давления в приносящих артериолах, по которым кровь поступает в клубочек, содержащий разветвленную капиллярную сеть. Кровь от клубочка оттекает по выносящей артериоле, которая вновь распадается на вторичную сеть капилляров, оплетающих проксимальные и дистальные канальцы (перитубулярные капилляры). Далее по венам кровь покидает почку и поступает в нижнюю полую вену. Из клубочков юкстамедуллярных нефронов выносящая артериола доставляет кровь в мозговое вещество, где образуются прямые сосуды (vasa recta), глубоко спускающиеся в него вместе с петлями Генле и участвующие в осмотическом концентрировании мочи. Таким образом, кровоснабжение почек устроено по типу двух последовательных систем сосудов с регулируемым сопротивлением. Механизмы мочеобразованияОсновные этапы процесса мочеобразованияМочеобразование складывается из трех основных процессов, представленных на рис.4.
Клубочковая фильтрацияОбразование мочи в почке начинается с ультрафильтрации плазмы крови в почечных клубочках. Жидкость проходит из просвета кровеносных капилляров в полость капсулы клубочка через клубочковый фильтр. 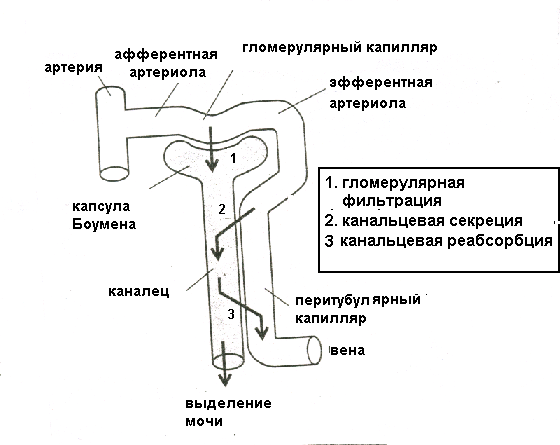 Рисунок 4 Основные процессы, обеспечивающие образование мочи Рассмотрим подробнее структуру этого фильтра и силы, обеспечивающие процесс фильтрации. Фильтрующая мембрана. Фильтрующая мембрана состоит из трех слоев: эндотелия капилляров, базальной мембраны и внутреннего листка капсулы Шумлянского - Боумена, который образован эпителиальными клетками – подоцитами. (Рис.5). Клетки эндотелия капилляров имеют очень тонкие периферические участки, в просвет сосуда выступает лишь область клетки, где находится ядро. Боковые части клетки пронизаны довольно крупными отверстиями, обычно затянутыми тонкими диафрагмами. При нормальной скорости кровотока крупные молекулы белка образуют над этими порами барьерный слой, что служит препятствием для прохождения через поры не только глобулинов, но и альбуминов. Таким образом, фенестрированный эндотелий капилляров ограничивает прохождение через клубочковый фильтр форменных элементов и белков, но свободно пропускает низкомолекулярные вещества, растворенные в плазме крови. Следующий барьер гломерулярного фильтра – базальная мембрана. Ее «поры» ограничивают прохождение молекул в зависимости от размера, формы и заряда. Так как мембрана имеет сетчатую структуру, образованную тонкими нитями, происходит ограничение прохождения молекул размером более 3,4 нм. Отрицательно заряженная стенка пор затрудняет прохождение молекул с одноименным зарядом. Поры не являются круглыми, что также существенно для ограничения фильтрации альбуминов. 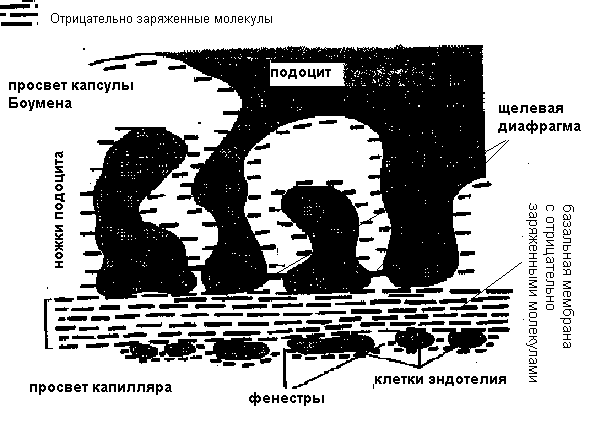 Рисунок 5 Структура клубочкового фильтра Последним барьером на пути фильтруемых веществ служат подоциты. Их отростки («ножки») прилегают к базальной мембране со стороны капсулы клубочка, между ножками подоцитов находятся пространства, по которым течет фильтруемая жидкость. Однако и в этом случае существует заслон на пути фильтруемых веществ – щелевые мембраны, перегораживающие пространство между ножками подоцитов. Они ограничивают прохождение альбуминов и других молекул с большой молекулярной массой. Поверхность ножек соседних отростков покрыта отрицательно заряженными сиалогликопротеинами, ограничивающими прохождение отрицательно заряженных частиц. Поскольку подоциты содержат внутри отростков актомиозиновые миофибриллы, они могут сокращаться и расслабляться, действуя как микронасосы, откачивающие фильтрат в полость капсулы. Такой многослойный фильтр обеспечивает сохранение белков в крови и образование практически безбелковой первичной мочи, в которой содержится большинство неорганических ионов и растворенных низкомолекулярных органических веществ почти в той же концентрации, что и в плазме. Движущей силой фильтрации является эффективное фильтрационное давление (Рф). Оно создаётся разностью между гидростатическим давлением крови в капиллярах клубочка (Pг) и противодействующими ему силами – онкотическим давлением белков плазмы крови (Рон) и гидростатическим давлением жидкости в капсуле клубочка (Рк). Соответственно, формула для расчета имеет следующий вид: Рф = Рг - (Рон +Рк). Подставим числовые значения давлений и произведем расчет: Рф= 70 мм рт.ст. – (30мм рт.ст.+20мм рт.ст.)=20мм рт.ст. Таким образом, эффективное фильтрационное давление равняется 20 мм рт.ст. Образовавшийся безбелковый фильтрат по своему составу близок плазме крови и имеет такую же, как и плазма, концентрацию осмотически активных веществ – 300 мосм/л. В обеих почках человека за 1 минуту образуется 110-130 мл ультрафильтрата. Таким образом, каждый мл плазмы из 600 мл, проходящих через сосуды почки за 1 минуту (величина почечного плазмотока), теряет примерно 1/5 часть своего объема. Объем профильтровавшейся за минуту первичной мочи принято называть скорость клубочковой фильтрации (СКФ). Метод определения СКФ и почечного плазмотока основан на принципе очищения (подробное описание метода дано в руководстве к лабораторным работам). Фильтрация считается довольно стабильным процессом, однако СКФ может изменяться при различных физиологических состояниях и при патологии. Регуляция почечного кровотока и СКФ происходит при участии симпатических нервов, ренин-ангиотензиновой системы и других факторов. За сутки образуется огромное количество первичной мочи – 180 л, окончательной мочи выделяется лишь 1,5-2,0 л. Остальная жидкость подвергается реабсорбции в почечных канальцах. В результате реабсорбции обратно в кровь возвращается большая часть воды и растворенных в ней веществ, «провалившихся» через фильтр и представляющих ценность для организма. Результатом сложной работы канальцев, в которых существует своеобразное «разделение труда», и явится образование окончательной мочи, состав и количество которой будет определятся водно-солевым балансом организма. Канальцевая реабсорбцияВ канальцах почки происходят два следующих этапа мочеобразования – процессы реабсорбции и секреции. Реабсорбция – процесс обратного всасывания веществ из просвета канальцев в кровь, при этом их выделение с мочой уменьшается. Секреция – процесс, обратный реабсорбции, в результате которого продукты, подлежащие выведению (экскреции), транспортируются в просвет канальцев; при этом их выделение с мочой увеличивается. Локализация важнейших транспортных процессов представлена на рис. 6. В основе реабсорбции и секреции лежат процессы мембранного транспорта через стенки канальцев. Они универсальны, и не отличаются от тех, что обеспечивают перенос веществ через другие плазматические мембраны (при всасывании в кишечнике, транспорте в капиллярах). 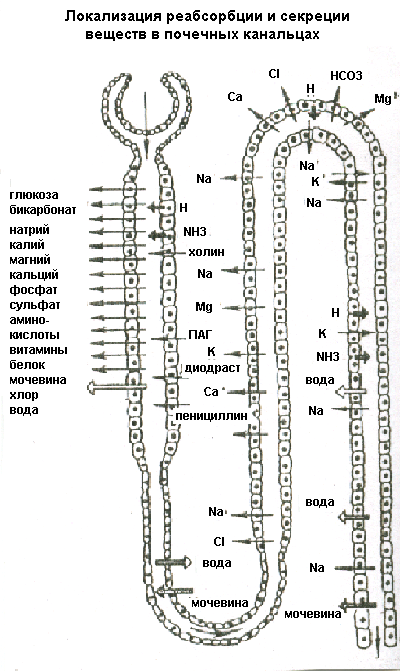 Рисунок 6 Реабсорбция и секреция в почечных канальцах. Направление стрелок указывает на направленность процесса. По многообразию транспортных процессов, их интенсивности, специфичности, избирательности - почки можно назвать уникальным органом. Проксимальная реабсорбция Образовавшийся в клубочках ультрафильтрат далее поступает в проксимальные канальцы. Эпителиальные клетки, образующие стенки проксимальных канальцев, как и все клетки, способные транспортировать вещества, имеют асимметричное строение, то есть характеризуются направленностью процессов от апикальной к базальной поверхности клетки. Апикальная мембрана клетки, обращенная в просвет канальца, имеет щеточную каемку, почти в 40 раз увеличивающую поверхность всасывания и обладающую большой сорбционной способностью. Базальная мембрана клеток образует складки, пространство между которыми называется базальным лабиринтом. Именно туда и поступает реабсорбированная жидкость, прежде чем попасть в перитубулярные капилляры. Между собой клетки соединяются так называемыми плотными контактами или плотными соединениями. На всем остальном протяжении они разделены довольно широким межклеточным пространством - базолатеральным лабиринтом. 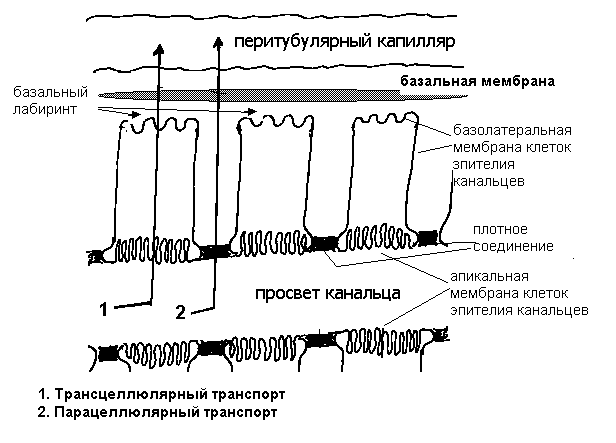 Рисунок 7 Схема строения эпителия проксимальных канальцев Из рисунка.7 видно, что для реабсорбции растворённых веществ и воды из просвета канальца в базальный лабиринт и далее в кровь, есть два пути: под номером 1 показан первый путь - трансцеллюлярный – через клетку. В этом случае вещество на своем пути должно преодолеть две плазматические мембраны (апикальную и базальную) и цитоплазму клетки. Второй путь реабсорбции - парацеллюлярный, между клетками - показан под номером 2. Он проходит через зоны плотных контактов. При таком транспорте могут быть использованы механизмы диффузии, осмоса и перенос вещества вместе с растворителем. Рассмотрение реабсорбции в проксимальном канальце следует начать с механизмов реабсорбции Nа, поскольку именно с Nа прямо или косвенно связана реабсорбция других веществ. Процесс реабсорбции натрия можно разделить на 3 этапа: прохождение через апикальную мембрану, движение через клетку к базальной мембране и эвакуация из клетки через базальную мембрану в межклеточное пространство. Рассмотрим их поэтапно. Апикальный транспорт. Вход Nа+ в клетку через апикальную мембрану представляет собой пассивный процесс. Он происходит по электрохимическому и концентрационному градиенту. Эти градиенты создаются благодаря активному транспорту натрия из клетки через базальные и базолатеральные мембраны. В апикальной мембране клеток имеются Nа+-каналы и Nа+-переносчики, облегчающие пассивный вход Nа+. Дело в том, что липидная основа мембраны непроницаема для гидрофильного иона Nа+ даже при наличии большого электрохимического градиента и отрицательного заряда на внутренней поверхности клеточной мембраны. Чтобы ионы Nа+ могли проникнуть через клеточную мембрану, в ней имеются гидрофильные белки - облегчители (пермеазы), образующие каналы, по которым проходит Nа+. Схема транспорта Nа+ в клетках проксимальных канальцев представлена на рис.8. Следующая группа механизмов апикального поступления Nа+ осуществляется с помощью вторично-активного транспорта. Котранспортёр (переносчик) может переносить Nа+ и какое-либо второе вещество в одном направлении по механизму симпорта. Примером такого вида транспорта является совместный перенос Nа+ с глюкозой и Nа+ с аминокислотами. По другому варианту вещество, например Н+, может выходить из клетки в обмен на ион Nа+, который движется в клетку: этот механизм называется противотранспорт или антипорт. Транспорт Na+ может быть сопряжён с транспортом бикарбоната и фосфатов. Вошедший в клетку Nа+ не смешивается с общим Nа+ клетки, а продвигается к местам эвакуации по специальной транспортной системе каналов, не нарушая клеточную внутреннюю среду. Б 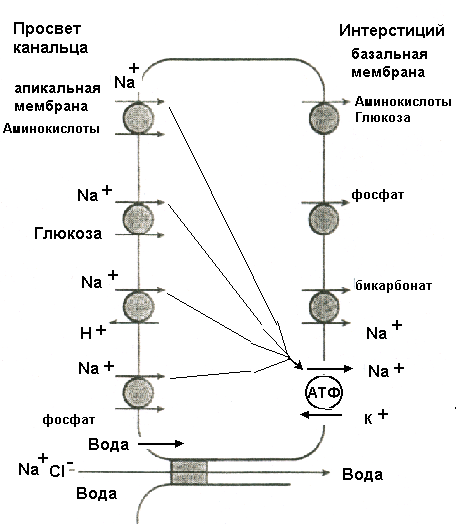 азальный транспорт. Через базальную и базолатеральную мембраны Nа+ транспортируется активно против электрохимического и концентрационного градиента с помощью Nа+–К+насосов. При этом ион Nа+ обменивается на ион К+. Главная роль в работе насосов принадлежит ферменту Nа+/К+ - АТФазе, которая вызывает распад молекулы АТФ, что и дает энергию, необходимую для реабсорбции. Такой вид транспорта называется первично-активный. То обстоятельство, что Nа+ постоянно откачивается из клетки, весьма важно, т.к. благодаря этому концентрация Nа+ в клетке остается низкой, что и обеспечивает совместно с электрохимическим потенциалом поступление в клетку новых порций натрия. Мы рассмотрели как Nа+ реабсорбируется через клетку (трансцеллюлярно), но некоторое количество Nа+ может проходить через зоны плотных контактов (парацеллюлярно) совместно с ионами Cl-. азальный транспорт. Через базальную и базолатеральную мембраны Nа+ транспортируется активно против электрохимического и концентрационного градиента с помощью Nа+–К+насосов. При этом ион Nа+ обменивается на ион К+. Главная роль в работе насосов принадлежит ферменту Nа+/К+ - АТФазе, которая вызывает распад молекулы АТФ, что и дает энергию, необходимую для реабсорбции. Такой вид транспорта называется первично-активный. То обстоятельство, что Nа+ постоянно откачивается из клетки, весьма важно, т.к. благодаря этому концентрация Nа+ в клетке остается низкой, что и обеспечивает совместно с электрохимическим потенциалом поступление в клетку новых порций натрия. Мы рассмотрели как Nа+ реабсорбируется через клетку (трансцеллюлярно), но некоторое количество Nа+ может проходить через зоны плотных контактов (парацеллюлярно) совместно с ионами Cl-. Вслед за электролитами пассивно по осмотическому градиенту из канальцев устремляется вода, она переносится частично через зоны клеточных контактов, частично через клетку по специальным водным каналам. Рисунок 8 Схема транспорта натрия в клетках проксимального канальца Двигаясь, вода захватывает и уносит в своем потоке растворенные в канальцевой жидкости вещества (главным образом Nа+, Cl- и мочевину). Этот механизм переноса называется «следование за растворителем» или «перенос веществ вместе с растворителем». В проксимальном канальце реабсорбируется большая часть профильтровавшегося Nа+ (65-80%) и 80% воды. Отличительной особенностью реабсорбции в проксимальном канальце является то, что вслед за Nа+ и другими осмотически активными веществами в эквивалентных количествах реабсорбируется вода, поэтому жидкость в проксимальном канальце остается изоосмотичной плазме крови и ее осмотическая концентрация составляет 300 мосм/л. Реабсорбция глюкозы. Через апикальную мембрану глюкоза поступает посредством системы симпорта с Nа+. Движение глюкозы опосредованно участием переносчика и является вторично-активным транспортом, поскольку энергия, необходимая для переноса глюкозы через апикальную мембрану, вырабатывается за счет транспортирующих Nа+ насосов. Через базальную мембрану глюкоза покидает клетку путем облегченной диффузии (рис 9). 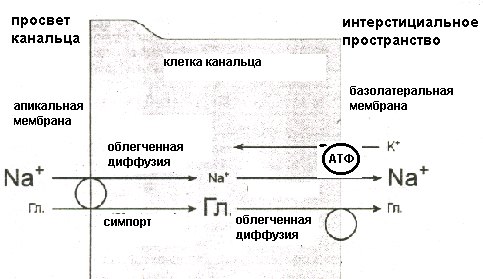 Рисунок 9 Механизм реабсорбции глюкозы в проксимальном канальце При нормальной концентрации глюкозы в крови (3,3-5,5 ммоль/л) вся фильтрируемая глюкоза практически полностью (100%) реабсорбируется клетками проксимальных канальцев и в окончательной моче она отсутствует. При повышении содержания глюкозы в крови с 5 до 10 ммоль/л глюкоза появляется в моче (глюкозурия). В этом случае оказывается превышен почечный порог - количество профильтровавшейся глюкозы превышает реабсорционную способность канальцев, транспортные системы максимально насыщаются, и избыток глюкозы выводится с мочой. Реабсорбционная способность канальцев определяется по глюкозе с использованием клиренсового метода (метод очищения, описание метода в руководстве к лабораторным работам). Подобно глюкозе в проксимальных канальцах почти полностью реабсорбируются аминокислоты. Реабсорбция аминокислот осуществляется путем вторично-активного транспорта, совместно с Nа+. Однако для аминокислот в апикальной мембране имеется не один переносчик, как для глюкозы, а 5-7 видов переносчиков, специфичных для различных групп аминокислот. В проксимальных канальцах реабсорбируются низкомолекулярные белки, которые в небольшом количестве (примерно 1,8 г/сут.) фильтруются и поступают в проксимальные канальцы. Фактически весь профильтровавшийся белок реабсорбируется, и экскреция его с мочой ничтожно мала (до 100 мг/сут.). При заболевании почек количество белков в моче может возрастать до 50,0 г/сут. (протеинурия). Реабсорбция белка происходит путем эндоцитоза. Молекула белка адсорбируется на апикальной мембране, мембрана впячивается, образуя вакуоли. Эти вакуоли отщепляются от мембраны клетки, сливаются в клетке с лизосомами, где под действием лизосомальных ферментов белок расщепляется до аминокислот. Продукты расщепления затем покидают клетку. В ПК также реабсорбируются бикарбонат, калий, фосфаты, витамины. Nа+ и другие реабсорбированные вещества, пройдя через стенку ПК, поступают в базальный и базолатеральный лабиринты. Затем жидкость эвакуируется в кровь перитубулярных капилляров, но механизм этот не совсем ясен. Существует гипотеза, согласно которой онкотическое давление в этих капиллярах значительно превышает онкотическое давление крови, поступившей в клубочки, поскольку в процессе фильтрации кровь теряет часть плазмы, и оттекающая от клубочка кровь, несколько сгущается. Другая гипотеза главное значение придает гидростатическому давлению, возникшему в базальном лабиринте из-за скопления в нем большого объема жидкости. Возможно, справедливы обе гипотезы. В клетках проксимальных канальцев реабсорбируется 100% глюкозы и аминокислот, белки, 80% Nа+, 80% воды, 80% бикарбоната, витамины и другие вещества. Но при всей сложности и многообразии транспортных процессов в проксимальных канальцах они являют собой чудо экономичности и эффективности. Оособенности проксимальной реабсорбции:
Осмотическое концентрирование и разведение мочи.Итак, после прохождения через проксимальные канальцы канальцевая жидкость поступает в петлю Генле и дистальный сегмент нефрона. С их участием связана одна из важнейших гомеостатических функций почки – осмотическое концентрирование мочи. Дело в том, что почки человека в нормальных условиях, как правило, производят гиперосмотическую по отношению к плазме крови мочу, т.е. работают в режиме концентрирования: осмолярность окончательной мочи колеблется от 600 до 900 мосм/л, т.е. в 3 раза может превышать осмолярность плазмы. Способность почки концентрировать мочу и экономить тем самым воду, возникла в эволюции с выходом животных на сушу, что позволило им перейти из водной среды обитания к наземному образу жизни и продолжить путь эволюционного развития в новом более перспективном направлении.  Рисунок 10 Взаимодействие петли Генле и собирательной трубки при образовании концентрированной мочи. В процессе осмотического концентрирования мочи принимают участие: петля Генле, дистальный каналец, собирательная трубка, сосуды и интерстиций мозгового вещества, которые функционируют как единая поворотно - противоточно-множительная система. В концентрировании мочи участвуют юкстамедуллярные нефроны с длинными петлями Генле, которые, как и собирательные трубки, глубоко проникают в мозговой слой почки. Процесс окончательного концентрирования мочи происходит в собирательных трубках, а условия для этого создаются работой всей поворотно – противоточно - множительной системы. Эта система создает гиперосмолярность мозгового вещества и при действии антидиуретического гормона (АДГ) заставляет воду переходить из собирательной трубки в интерстиций, а затем в кровеносные сосуды мозгового вещества, в результате чего образуется концентрированная моча. Ключевым вопросом при осуществлении концентрирования мочи является вопрос о том, каким образом интерстициальная жидкость мозгового вещества становится гиперосмолярной. Попытаемся ответить на него. Из рис.10 видно, что из проксимального канальца жидкость, изоосмотическая плазме крови, с концентрацией 300 мосм/л ,поступает в тонкое нисходящее колено петли Генле и, продвигаясь по нему, начинает терять воду; в результате ее осмотическая концентрация прогрессивно нарастает и на изгибе петли в сосочке достигает своего максимума (1400мосм/л). Затем она поворачивает и течет по восходящему колену в противоположном направлении (отсюда название поворотно-противоточная система). При этом происходит ее разбавление и уменьшение осмолярности до 100 мосм/л. Эти процессы обусловлены разными функциональными свойствами стенок этих участков петли: «маховиком» этой системы является толстый восходящий отдел петли, который совершенно непроницаем для воды, но активно реабсорбирует Nа+ и Cl-. В апикальной мембране клеток этого отдела транспорт Nа+ осуществляется одновременно с ионами К+ и двумя ионами Cl- с помощью Nа+, К+,2Cl-, котранспортера. Через базальную мембрану клеток Nа+ переносится активно. NаCl поступает в интерстиций и обуславливает выход воды из нисходящего колена петли, стенки которого, напротив, обладают высокой проницаемостью для воды, но не пропускает ионы Nа+. Таким образом, реабсорбция хлорида натрия восходящей частью петли “отвечает” за реабсорбцию воды в нисходящей части. Разберем это на простой схеме (рис.11). Представим себе, что петля Генле заполнена неподвижной жидкостью, которая поступила из проксимального канальца, тогда осмотическая концентрация в любой части петли Генле равна 300 мосм/л. (1). Теперь допустим (2), что активная транспортная система в восходящей части петли осуществляет реабсорбцию хлорида натрия в интерстициальное пространство до тех пор, пока не установится предельный градиент (скажем 200 мосм/л) между жидкостью, находящейся в восходящем колене петли Генле и жидкостью интерстициального пространства. (Стенки этой части петли непроницаемы для воды). Теперь существует разница в осмолярности между жидкостью в нисходящем колене (300 мосм/л) и окружающей каналец интерстициальной жидкостью (400 мосм/л). Поскольку стенки нисходящего колена петли хорошо проницаемы для воды, вода пассивно выходит из его просвета в интерстициальное пространство, в котором существует более высокая осмолярность, возникшая вследствие реабсорбции хлорида натрия в восходящем колене, и осмолярность жидкости в нисходящем колене увеличивается до 400 мосм/л. (3). Теперь усложним схему: пусть жидкость в петле не стоит на месте, а непрерывно движется, тогда по мере продвижения вниз по нисходящему колену из нее уходит все больше воды, концентрация внутриканальцевой жидкости все больше увеличивается и, поскольку устанавливается осмотическое равновесие, концентрация интерстициальной жидкости увеличивается на ту же величину (200 мосм/л). Градиент в 200 мосм/л, так называемый поперечный или горизонтальный градиент, поддерживается на каждом из «этажей» мозгового вещества. 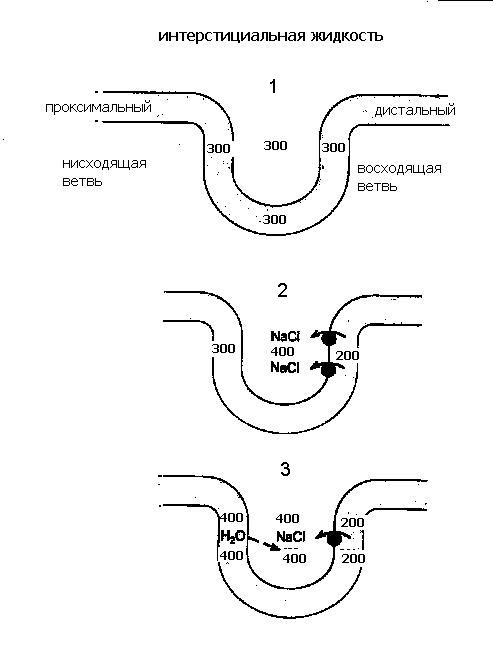 Рисунок 11 Схема взаимодействия нисходящего и восходящего колена в процессе концентрирования канальцевой жидкости. Таким образом, между соседними участками нисходящего и восходящего колена разность осмотической концентрации невелика. По ходу петли и по длине почечного сосочка – по вертикали – этот одиночный эффект нарастает, суммируется (умножается), и в результате формируется значительно больший перепад осмотического давления – так называемый корково-сосочковый вертикальный осмотический градиент. 300 мосм/л кора 6  00 мосм/л 00 мосм/л900 мосм/л 1200 мосм/л 1400 мосм/л сосочек Поэтому система называется ещё и множительной. Таким образом, нисходящее и восходящее колено петли тесно соприкасаются друг с другом, расположены параллельно, оказывают друг на друга влияние и функционируют как единый сопряженный механизм – противоточно-поворотно-множительная система. Благодаря работе этой системы в интерстиции мозгового вещества создается корково-сосочковый осмотический градиент. Следует отметить, что осмолярность интерстиция создается не только ионами хлорида натрия. Примерно половина осмолярности обусловлена присутствием в нем мочевины. У мочевины имеется свой кругооборот в почке, в частности из собирательной трубки она пассивно диффундирует в интерстиций мозгового вещества и тем самым, увеличивает его осмолярность. Прямые сосуды мозгового вещества, подобно коленам петли Генле, тоже образуют поворотно-противоточную систему. Они расположены параллельно петлям Генле и в них происходят такие же изменения осмолярности, что и в петле. Это сохраняет продольный осмотический градиент в мозговом веществе, не позволяя ему вымываться. По мере того, как канальцевая жидкость двигалась по нисходящему колену, ее осмолярность нарастала и в районе изгиба достигала 1400 мосм/л, так же как и осмолярность интерстиция, но по мере движения по восходящему колену она вновь разбавилась до 100 мосм/л. Далее в дистальном канальце происходит реабсорбция NаCl, воды и др. веществ и осмотическая концентрация вновь становится – 300 мосм/л, но по-прежнему канальцевая жидкость изоосмотична крови, т.е.в петле концентрирование не произошло. Из дистального канальца жидкость поступает в собирательную трубку, где и будет происходить формирование окончательной мочи и процесс ее концентрирования. Неслучайно собирательные трубки расположены параллельно петлям Генле и прямым сосудам. Они транзитом проходят через все зоны почки и на всем протяжении окружены интерстицием с прогрессивно нарастающим в направлении от коры к сосочку осмотическим давлением. Как видно из рисунка 10, осмолярность интерстициальной жидкости на каждом уровне идентична этой величине в нисходящем колене и СТ. Другими словами вокруг СТ на каждом “этаже” мозгового вещества имеется горизонтальный осмотический градиент в 200 мосм/л, а по вертикали - мощный корково-сосочковый осмотический градиент, созданный поворотно - противочной системой петли Генле. Таким образом, можно сказать, что петля Генле «работает» на собирательную трубку, создавая в интерстиции мозгового вещества зону гиперосмии. Это и будет та сила, которая способна вытянуть воду из собирательной трубки и произвести концентрирование мочи. Когда канальцевая жидкость поступает в собирательную трубку, ее осмолярность находится на том же уровне, что и осмолярность интерстициальной жидкости в этой зоне почки. В районе сосочка величина осмолярности достигает максимума (у человека она равна 1400 мосм/л), поэтому максимальная осмотическая концентрация мочи у человека тоже может достигать 1400 мосм/л. Резюмируя вышеизложенное, можно представить следующую схему основных процессов, обеспечивающих осмотическое концентрирование мочи. Основным элементарным актом в концентрирующей системе почки является создание поперечного (горизонтального) осмотического градиента между восходящим и нисходящим коленами петли Генле. В результате их взаимодействия, и за счет противоточного перемещения канальцевой жидкости и крови, устанавливается определенная величина продольного (вертикального) осмотического градиента. Осмолярность окончательной мочи будет зависеть от проницаемости стенок собирательных трубок для воды, основным регулятором которой является АДГ. Если АДГ много, проницаемость стенок собирательной трубки для воды увеличивается и вода, подчиняясь осмотическому градиенту, существующему в интерстиции мозгового вещества, реабсорбируется и поступает в интерстиций и далее в кровь. Жидкость в собирательных трубках приходит в состояние равновесия с окружающим гиперосмолярным интерстицием, выделяется концентрированная моча. Если АДГ мало, стенки собирательных трубок становятся водонепроницаемыми, образуется гипотоническая моча, диурез увеличивается. Таким образом, окончательный объём и состав мочи обусловлены функцией СТ. Их роль в осмотическом концентрировании и разведении мочи определяется как особенностями их анатомического расположения в почке, так и воздействием АДГ на проницаемость их стенок для воды. Другими словами, то, что произойдет с мочой в собирательной трубке и определит окончательное ее концентрирование. В отличие от проксимального канальца, где Nа+ и вода реабсорбировались вместе, в дистальном канальце и собирательной трубке вода и Nа+ реабсорбируются независимо. Именно это обстоятельство и позволяет дистальному отделу нефрона производить как концентрированную, так и разбавленную мочу. Дистальная реабсорбция называется факультативной (необязательной). Таким образом: 1. В процессе осмотического концентрирования мочи принимают участие петля Генле, дистальный каналец, собирательная трубка, сосуды и интерстиций мозгового вещества. Их объединение в единый концентрирующий аппарат почки обусловлено их взаимным расположением и общностью протекающих в них процессов. 2. Процесс окончательного концентрирования мочи происходит в собирательной трубке за счет факультативной реабсорбции воды. 3. Условия для нее создает корково – сосочковый осмотический градиент мозгового вещества, созданный поворотно-противоточно-множительной системой петли Генле. 4. Факультативная реабсорбция воды в собирательной трубке регулируется АДГ. 5. В дистальном сегменте нефрона натрий и вода реабсорбируются независимо. Механизм действия АДГ на почкуАДГ увеличивает процесс факультативной реабсорбции воды в собирательной трубке, изменяя проницаемость ее стенок. В результате уменьшается объем образующейся мочи – диурез (отсюда и происходит название гормона – антидиуретический) и выделяется концентрированная моча. АДГ синтезируется в нейросекреторных клетках гипоталамических ядер (супраоптического и паравентрикулярного). По отросткам этих клеток с током аксоплазмы гормон перемещается в заднюю долю гипофиза, откуда и поступает в кровь. Клеточный механизм действия АДГ в настоящее время изучен достаточно хорошо (рис.12). АДГ действует на главные клетки собирательных трубок, связывается сV2 рецепторами на базолатеральной мембране клеток, в результате активируется аденилатциклаза (АЦ), которая стимулирует образование цАМФ и запускается каскад реакций, приводящий к увеличению водной проницаемости. Условно эффекты АДГ можно разделить на апикальный и базальный. В апикальной мембране цепь процессов завершается встраиванием в мембрану клеток особого белка аквапорина. Аквапорин образует в гидрофобной мембране каналы (поры), по которым вода свободно перемещается из просвета собирательной трубки в интерстиций. В настоящее время известны 8 видов аквапоринов, часть из которых «работает» и в других отделах канальцев. Что касается базального эффекта, то согласно гипотезе А.Г. Гинецинского, в процесс вовлекаются ферменты гидролиза ГАГ (гликозаминогликанов), входящих в состав межклеточного вещества. Таким ферментом, в частности, является гиалуронидаза, а субстратом – гиалуроновая кислота. В результате происходит деполимеризация (разжижение) базальной мембраны, что облегчает движение воды по осмотическому градиенту. 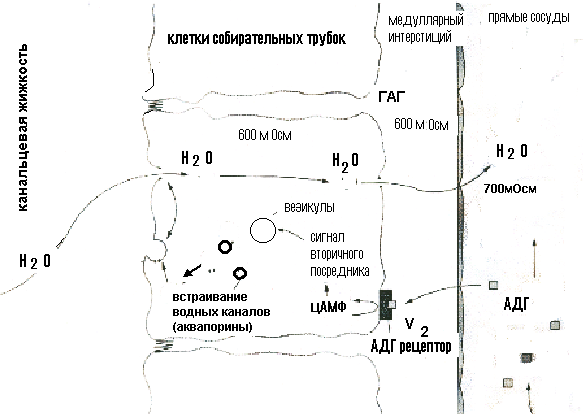 Рисунок 12 Механизм действия АДГ на клетки собирательных трубок Схема, иллюстрирующая эффекты АДГ 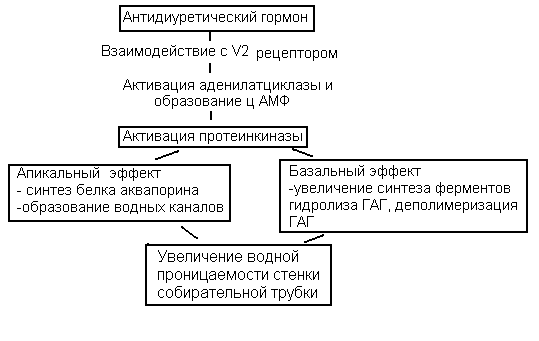 Существует заболевание, связанное с недостаточной секрецией АДГ. Оно называется несахарный диабет и характеризуется выделением значительных количеств мочи (до 25 л в сутки). Канальцевая секреция Секреция это процесс переноса веществ через клетки из крови в просвет канальцев. В почке секретируются органические кислоты (парааминогиппуровая кислота (ПАГ), диодраст), органические основания (холин, гуанидин), неорганические вещества (калий и водород), конечные продукты обмена (мочевина, мочевая кислота, креатинин), Важно отметить, что секретируются многие лекарственные препараты и некоторые красители (пенициллин, салицилаты, сульфаниламиды, фуросемид, этакриновая кислота, морфин, хинин, атропин, фенолрот). Величина канальцевой секреции определяется методом очищения (руководство к проведению лабораторных работ). |