гормоны. Гормоны это органические вещества, которые образуются в тканях одного типа (эндокринные железы, или железы внутренней секреции), поступают в кровь, переносятся по кровяному руслу в ткани другого типа (тканимишени), где оказывают своё биологическое действие (т е.
 Скачать 216.79 Kb. Скачать 216.79 Kb.
|

|
Гормоны – это органические вещества, которые образуются в тканях одного типа (эндокринные железы, или железы внутренней секреции), поступают в кровь, переносятся по кровяному руслу в ткани другого типа (ткани-мишени), где оказывают своё биологическое действие (т. е. регулируют обмен веществ, поведение и физиологические функции организма, а также рост, деление и дифференцировку клеток). Классификация гормонов По химической природе гормоны делятся на следующие группы: 1)пептидные – гормоны гипоталамуса, гипофиза, инсулин, глюкагон, гормоны паращитовидных желез; 2)производные аминокислот – адреналин, тироксин; 3)стероидные – глюкокортикоиды, минералокортикоиды, мужские и женские половые гормоны; 4)эйкозаноиды – гормоноподобные вещества, которые оказывают местное действие; они являются производными арахидоновой кислоты (полиненасыщенная жирная кислота). По месту образования гормоны делятся на гормоны гипоталамуса, гипофиза, щитовидной железы, паращитовидных желёз, надпочечников (коркового и мозгового вещества), женские половые гормоны, мужские половые гормоны, местные или тканевые гормоны. По действию на биохимические процессы и функции гормоны делятся на: 1)гормоны, регулирующие обмен веществ (инсулин, глюкагон, адреналин, кортизол); 2) гормоны, регулирующие обмен кальция и фосфора (паратиреоидный гормон, кальцитонин, кальцитриол); 4 3)гормоны, регулирующие водно-солевой обмен (альдостерон, вазопрессин); 4)гормоны, регулирующие репродуктивную функцию (женские имужские половые гормоны); 5)гормоны, регулирующие функции эндокринных желёз (адренокортикотропный гормон, тиреотропный гормон, лютеинизирующий гормон, фолликулостимулирующий гормон, соматотропный гормон); 6)гормоны стресса (адреналин, глюкокортикоиды и др.); 7)гормоны, влияющие на ВНД (память, внимание, мышление, поведение, настроение): глюкокортикоиды, паратиреоидный гормон, тироксин, адренокортикотропный гормон) Свойства гормонов 1)Высокая биологическая активность. Концентрация гормонов в крови очень мала, но их действие сильно выражено, поэтому даже небольшое увеличение или уменьшение уровня гормона в крови вызывает различные, часто значительные, отклонения в обмене веществ и функционировании органов и может привести к патологии. 2)Короткое время жизни, обычно от нескольких минут до получаса, после чего гормон инактивируется или разрушается. Но с разрушением гормона его действие не прекращается, а может продолжаться в течение часов и даже суток. 3)Дистантность действия. Гормоны вырабатываются в одних органах (эндокринных железах), а действуют в других (тканяхмишенях). 4)Высокая специфичность действия. Гормон оказывает своё действие только после связывания с рецептором. Рецептор – это сложный белок-гликопротеин, состоящий из белковой и углеводной частей. Гормон связывается именно с углеводной частью рецептора. Причём строение углеводной части имеет уникальную химическую структуру и соответствует пространственному строению гормона. Поэтому гормон безошибочно, точно, специфично связывается только со своим рецептором, несмотря на малую концентрацию гормона в крови. Не все ткани одинаково реагируют на действие гормона. Высокой чувствительностью к гормону обладают те ткани, в которых 5 имеются рецепторы к данному гормону. В таких тканях гормон вызывает наиболее выраженные сдвиги в обмене веществ и функциях. Если рецепторы к гормону есть во многих, или почти во всех тканях, то такой гормон оказывает общее действие (тироксин, глюкокортикоиды, соматотропный гормон, инсулин). Если рецепторы к гормону присутствуют в весьма ограниченном числе тканей, то такой гормон обладает избирательным действием. Ткани, в которых имеются рецепторы к данному гормону, называются ткани-мишени. В тканях-мишенях гормоны могут воздействовать на генетический аппарат, мембраны, ферменты. Типы биологического действия гормонов 1)Метаболическое – действие гормона на организм проявляется регуляцией обмена веществ (например, инсулин, глюкокортикоиды, глюкагон). 2)Морфогенетическое – гормон действует на рост, деление и дифференцировку клеток в онтогенезе (например, соматотропный гормон, половые гормоны, тироксин). 3)Кинетическое или пусковое – гормоны способны запускать функции (например, пролактин – лактацию, половые гормоны – функцию половых желёз). 4)Корригирующее. Гормонам принадлежит важнейшая роль в адаптации человека к различным факторам внешней среды. Гормоны изменяют обмен веществ, поведение и функции органов так, чтобы приспособить организм к изменившимся условиям существования, т.е. осуществляют метаболическую, поведенческую и функциональную адаптацию, тем самым поддерживают постоянство внутренней среды организма. Механизм действия пептидных гормонов и адреналина Рецепторы к этим гормонам находятся на наружной поверхности клеточной мембраны, и гормон внутрь клетки не проникает. Действие гормона в клетку передается при помощи так называемых вторых посредников, к которым относятся циклический АМФ (цАМФ), циклический ГМФ (цГМФ), кальций, инозитолтрифосфат, диацилглицерол (диглицерид) и некоторые другие. В системе передачи регуляторного сигнала они называются 6 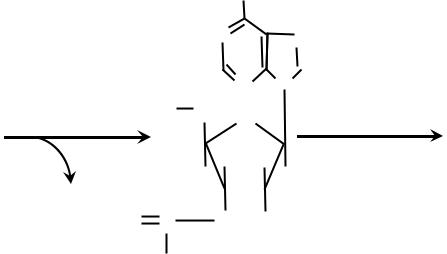 вторыми посредниками, потому что первым посредником является сам гормон. Каждый из вторых посредников активирует специфическую протеинкиназу. Протеинкиназы фосфорилируют ферменты, и это изменяет активность ферментов. Главным вторым посредником является цАМФ (рис. 1). Большинство гормонов действует через него. Другие посредники, действуя через свои протеинкиназы, могут изменять содержание цАМФ в клетке путём повышения или снижения активности ферментов, синтезирующих или разрушающих цАМФ.
ОН Циклический-3', 5'-АМФ Рис. 1. Метаболизм цАМФ цАМФ Циклический АМФ образуется в клетке из АТФ под действием аденилатциклазной системы (рис. 2). В состав аденилатциклазной системы входят: рецептор, G-белок и фермент аденилатциклаза. G- белок называется так потому, что он способен связывать гуаниловые нуклеотиды (ГТФ или ГДФ). Существует 2 разновидности G-белка: Gs - стимулирует аденилатциклазу и увеличивает образование цАМФ и Gi – ингибирует аденилатциклазу и уменьшает образование цАМФ. Белки Gs и Gi оказывают своё активирующее или ингибирующее действие только когда находятся в' активном состоянии. G-белок 7 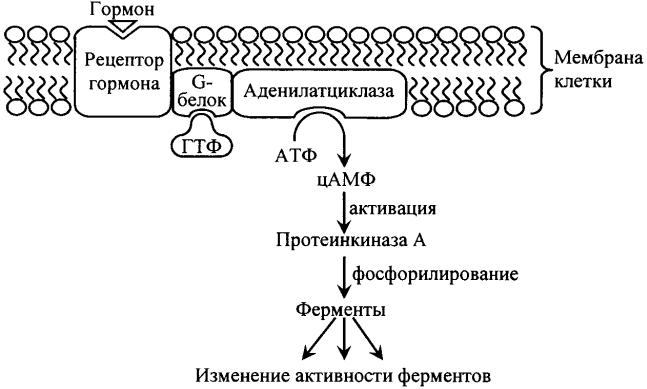 активен, когда он связан с ГТФ, и наоборот, связанный с ГДФ, G- белок неактивен. Пока гормон не действует на клетку, аденилатциклазная система неактивна; все её компоненты разобщены и с G-белком связан ГДФ. Однако после связывания гормона с рецептором происходит последовательное изменение конформации всех компонентов аденилатциклазной системы, G-белок обменивает ГДФ на ГТФ, переходит в активное состояние и активирует аденилатциклазу, которая из АТФ синтезирует цАМФ. Циклический АМФ, в свою очередь, активирует специфическую цАМФ–зависимую протеинкиназу (протеинкиназу А), которая фосфорилирует внутриклеточные ферменты, в результате чего изменяется активность ферментов. Рис. 2. Аденилатциклазная система Протеинкиназа состоит из 4 субъединиц (тетрамер), две из которых являются регуляторными, а две – каталитическими (рис. 3). В таком виде протеинкиназа неактивна. При связывании протеинкиназой 4 молекул цАМФ происходит отсоединение (диссоциация) каталитических субъединиц, которые фосфорилируют белки (ферменты), изменяя их активность. 8  Разрушается цАМФ фосфодиэстеразой. Рис. 3. Активация протеинкиназы А под действием цАМФ К – каталитические субъединицы, Р – регуляторные субъединицы цГМФ Циклический ГМФ образуется из ГТФ под действием гуанилатциклазы по аналогии с синтезом цАМФ. Циклический ГМФ активирует специфическую цГМФ–зависимую протеинкиназу или протеинкиназу G, которая фосфорилирует ферменты, что сопровождается изменением их активности. Разрушается цГМФ, как и цАМФ, фосфодиэстеразой. Кальций Концентрация Са2+ во внеклеточной жидкости в 10 000 раз больше, чем в цитоплазме. Такая концентрация кальция оказалась бы потенциально летальной для клетки. Клетка борется с избытком Са2+ в цитоплазме путём откачивания его наружу с помощью Са2+- АТФазы, расположенной в плазматической мембране. Под действием гормонального сигнала концентрация Са2+ в клетке возрастает в 10 и более раз за счет притока Са2+ из внеклеточной жидкости, а также выхода Са2+ из ЭПР и митохондрий. В клетке Са2+ связывается с белком кальмодулином и активирует специфическую Са2+-кальмодулин-зависимую протеинкиназу. В ответ на гормональный сигнал и повышение концентрации Са2+ в 9 + 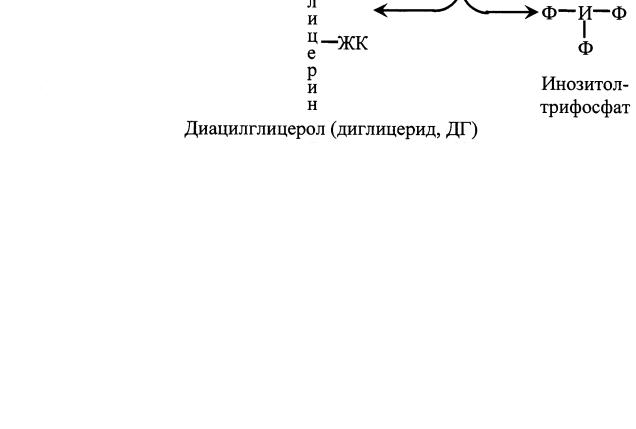 клетке специфическая протеинкиназа катализирует фосфорилирование множества внутриклеточных ферментов, регулируя, тем самым, их активность. Инозитолтрифосфат и диглицерид Эти вторые посредники являются производными фосфолипида мембран, фосфатидилинозитола. Указанный фосфолипид фосфорилируется в мембране под действием фермента фосфатидилинозитолкиназы при участии 2 молекул АТФ (рис. 4). Рис. 4. Схема образования диглицерида и инозитолтрифосфата из фосфатидилинозитола. ЖК – жирная кислота, И – инозитол, Ф – фосфат неорганический Образующийся фосфатидилинозитолдифосфат расщепляется под действием специфического мембраносвязянного фермента фосфолипазы С, в результате чего образуются два вторых посредника: диглицерид (диацилглицерол, ДАГ) и инозитолтрифосфат (ИФ3). Биологические эффекты этих двух вторых посредников реализуются по-разному. ДАГ активирует Са2+-зависимую протеинкиназу С, которая катализирует фосфорилирование внутриклеточных ферментов, изменяя их активность. ИФ3 связывается со специфическим рецептором на ЭПР, способствуя выходу из него Са2+ в цитоплазму. Далее Са2+ связывается с кальмодулином и активирует Са2+-кальмодулин-зависимую протеинкиназу, которая катализирует фосфорилирование ферментов, изменяя их активность (рис. 5). Рис. 5. Система вторых посредников – диацилглицерола и инозитолтрифосфата ЖК – жирная кислота, И – инозитол, Ф – фосфат неорганический 11 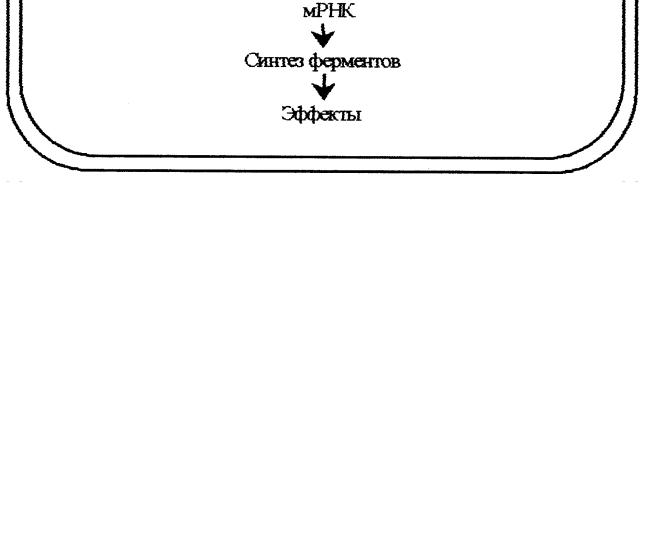 Механизм действия стероидных гормонов и тироксина Рецепторы к этим гормонам находятся внутри клетки, в цитоплазме. Гормон легко и быстро проникает через мембрану внутрь клетки, взаимодействует с рецептором с образованием гормон-рецепторного комплекса. Этот комплекс переносится в ядро, где связывается с ядерным хроматином в определенном участке ДНК. С этого участка ДНК начинается синтез специфических мРНК, которые затем выходят в цитоплазму и служат матрицей для синтеза ферментов или других белков, необходимых клетке и обеспечивающих ответ клетки на действие гормона (рис. 6). Рис. 6. Механизм действия стероидных гормонов и тироксина 12 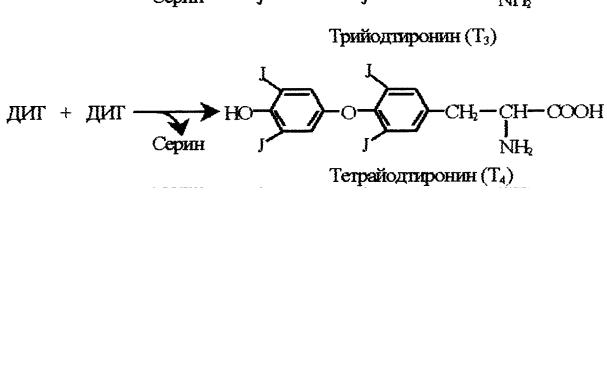 Таким образом, под действием пептидных гормонов и адреналина в клетке изменяется активность ферментов; стероидные гормоны и тироксин приводят к изменению количества ферментов. ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ Вщитовидной железе синтезируются тиреоидные гормоны – тироксин (Т4) и трииодтиронин (Т3). Для синтеза этих гормонов необходим иод, который активно захватывается из крови клетками фолликулов щитовидной железы. Тироксин и трииодтиронин являются производными аминокислоты тирозина. Тироксин содержит всвоей молекуле 4 атома иода, в состав трииодтиронина входит 3 атома иода. Вклетках эпителия фолликулов щитовидной железы находится белок тиреоглобулин. Это гликопротеин, содержащий много остатков аминокислоты тирозина (около 3 % от массы белка). Синтез тиреоидных гормонов идет из тирозина и атомов иода именно в составе молекулы тиреоглобулина и включает 2 стадии. На апикальных мембранах клеток фолликулов сначала происходит иодирование тирозина с образованием моноиодтирозина (МИТ) и дииодтирозина (ДИТ). Следующим этапом является конденсация МИТ и ДИТ с образованием Т3 и Т4 (рис. 7). Рис. 7. Схема синтеза тиреоидных гормонов 13 Такая иодированная молекула тиреоглобулина секретируется в просвет фолликула, в коллоид. Когда в щитовидную железу приходит сигнал в виде ТТГ (тиреотропного гормона), клетки фолликула захватывают капельки коллоида вместе с тиреоглобулином, ферменты лизосом протеазы гидролизуют белок до аминокислот, и готовые Т3 и Т4 поступают в кровь. В крови тиреоидные гормоны связываются с белкомпереносчиком и в таком виде транспортируются в ткани-мишени. Концентрация Т4 в крови в 10 раз больше, чем Т3, поэтому Т4 называют главной формой тиреоидных гормонов в крови. Но Т3 в 10 раз активнее, чем Т4. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||