БХ 5 1 и 2 часть. Гормоны поджелудочной железы
 Скачать 1.33 Mb. Скачать 1.33 Mb.
|

2 часть1. Гормоны надпочечников 1.1 Гормоны коры надпочечников: классификация по химической структуре, схема основных этапов синтеза из холестерола, механизм действия Надпочечники являются парными эндокринными железами. Они состоят из коркового вещества и мозгового вещества. В корковом веществе синтезируются стероидные гомоны (кортикостероиды), а в мозговом – гормоны, являющиеся производными аминокислот. Стероидные гормоны коры надпочечников делятся на три группы: глюкокортикоиды, минералокортикоиды и половые гормоны. Холестерол является источником для синтеза всех стероидных гормонов. Структурные особенности различных стероидных гормонов можно иллюстрировать следующей схемой: 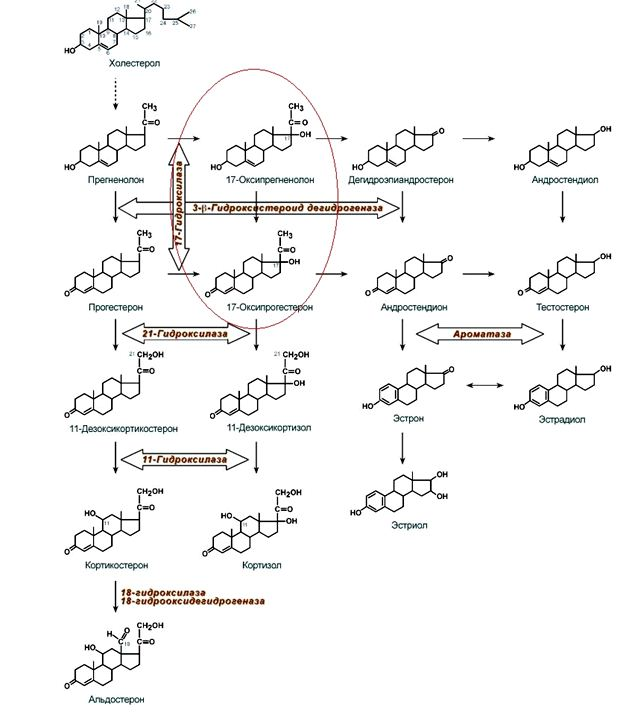 Рис. Схема синтеза стероидных гормонов (источник заимствования – ресурсы Сети Интернет). Синтез гормонов коры надпочечников В гломерулярной зоне коры надпочечников (краевой) синтезируются минералокортикоиды, в пучковой (средней) — глюкокортикоиды, а в ретикулярной (сетчатой) – половые гормоны. Источником для синтеза стероидных гормонов является холестерол (см. рис.). Он может либо синтезироваться в надпочечниках, либо поступать в надпочечники из печени в составе ЛПНП. Первые две реакции являются общими в синтезе всех стероидных гормонов 17-гидроксилаза не экспрессируется в гломерулярной зоне коры надпочечников, поэтому синтез кортикостерона и альдостерона запускают лишь 21- и 11-гидроксилазы. В результате кортикостерон обладает слабой активностью как глюко-, так и минералокортикоида (см. рис.). Напомним, что у глюкокортикоидов преобладают эффекты на углеводный и белковый обмены. Минералокортикоиды, преимущественно, контролируют обмен Na+ и K+. Стимуляторами секреции глюкокортикоидов являются физический и эмоциональные стрессы, состояние тревоги, страха, волнения и боль. Основной контроль за синтезом и секрецией глюкокортикоидов реализует гипоталамо-гипофизарная система по принципам нисходящих стимулирующих сигналов и отрицательной обратной связи от самих периферических гормонов и их тканей-мишеней. Мишенями для действия глюкокортикоидов (ГК) является целый ряд тканей: печень, жировая ткань, почки, кожа и ее производные, костная и мышечная ткани, лимфоидная ткань, соединительная. . Влияние на клеточный метаболизм мощное, выражается в их гипергликемическом действии, мобилизации липидов, активации катаболизма коллагена, других белков в тканях-мишенях. Являются антагонистами инсулина. Снижают проницаемость клеточных мембран для глюкозы в общих с ним тканях-мишенях. Стимулируют глюконеогенез, активируя синтез его ключевых ферментов, а также аминотрансфераз в печени. Это позволяет потреблять в качестве субстратов глюконеогенеза аминокислоты, высвобождаемые в результате распада белков из лимфоидной, эпителиальной, мышечной, соединительной, костной тканей. В жировой ткани, активируя синтез ТАГ- и других липаз, тем самым оказывают пермиссивное действие на глюкагон, адреналин, СТГ, АКТГ (англ. permission - позволение). Глюкокортикоиды в норме увеличивают перемещение лимфоцитов, моноцитов, эозинофилов и базофилов в лимфоидную ткань. Они подавляют синтез ферментов – фосфолипазы А2 и циклооксигеназы – ключевых ферментов синтеза эйкозаноидов. Для ГК характерна супрессия активности иммунной системы, регуляция, воспалительных и аллергических реакций. Превращение холестерола в прегненолон в результате отщепления шеститиуглеродного фрагмента от боковой цепи (фермент С21-22 десмолаза). Десмолаза является ключевым ферментом в биосинтезе стероидов. Фермент активен в фосфорилированном виде. Его фосфорилирование стимулирует АКТГ. 2. Окисление и изомеризация прегненолона в прогестерон. Прегненолон является предшественником всех стероидных гормонов (см. рис.). В желтом теле (в яичниках) синтез стероидов останавливается на этапе образования прогестерона. Во всех других стероидпродуцирующих тканях прогестерон является только промежуточным продуктом синтеза других стероидов. Дальнейшие преобразования прогестерона проходят следующим образом: сначала прогестерон подвергается гидроксилированию в положениях С17 или С21, в результате чего образуются различные функциональные классы стероидов. Дальнейшее гидроксилирование прогестерона в положениях С21 или С11 приводит к образованию минералокортикоидов, в С17 — глюкокортикоидов. Ввиду общности природы и происхождения, гормоны стероидной группы отличают выраженные липофильные свойства. Поступая в кровь, связываются со специфическим транспортным белком – транскортином или альбуминами крови. Реализация механизмов действия гормонов данной природы в клетках-мишенях происходит по внутриклеточному пути. Это обуславливает длительность развития их эффектов, но продолжительность действия, что связано с активацией матричных синтезов клеточных белков, проникающим в клетку гормоном. Глюкокортикоиды Кортизол наиболее активный глюкокортикоид, является основным гормоном данной группы у человека. Он формирует ОН-группу в 17-м положении (как и кортизон), а затем подобно кортикостерону, получает гидроксильный радикал у С11-атома углерода. Формирование гидроксильного радикала по С17-углероду – ключевая реакция синтеза группы глюкокортикоидов и половых гормонов, ее реализует 17-гидроксилаза (см. рис.). Затем происходит гидроксилирование по С21, С11-атомам. 17-гидроксилаза не экспрессируется в гломерулярной зоне коры надпочечников, поэтому синтез кортикостерона и альдостерона запускают лишь 21- и 11-гидроксилазы. В результате кортикостерон обладает слабой активностью как глюко-, так и минералокортикоида (см. рис.). Напомним, что у глюкокортикоидов преобладают эффекты на углеводный и белковый обмены. Минералокортикоиды, преимущественно, контролируют обмен Na+ и K+. Стимуляторами секреции глюкокортикоидов являются физический и эмоциональные стрессы, состояние тревоги, страха, волнения и боль. Основной контроль за синтезом и секрецией глюкокортикоидов реализует гипоталамо-гипофизарная система по принципам нисходящих стимулирующих сигналов и отрицательной обратной связи от самих периферических гормонов и их тканей-мишеней. Мишенями для действия глюкокортикоидов (ГК) является целый ряд тканей: печень, жировая ткань, почки, кожа и ее производные, костная и мышечная ткани, лимфоидная ткань, соединительная. . Влияние на клеточный метаболизм мощное, выражается в их гипергликемическом действии, мобилизации липидов, активации катаболизма коллагена, других белков в тканях-мишенях. Являются антагонистами инсулина. Снижают проницаемость клеточных мембран для глюкозы в общих с ним тканях-мишенях. Стимулируют глюконеогенез, активируя синтез его ключевых ферментов, а также аминотрансфераз в печени. Это позволяет потреблять в качестве субстратов глюконеогенеза аминокислоты, высвобождаемые в результате распада белков из лимфоидной, эпителиальной, мышечной, соединительной, костной тканей. В жировой ткани, активируя синтез ТАГ- и других липаз, тем самым оказывают пермиссивное действие на глюкагон, адреналин, СТГ, АКТГ (англ. permission - позволение). Глюкокортикоиды в норме увеличивают перемещение лимфоцитов, моноцитов, эозинофилов и базофилов в лимфоидную ткань. Они подавляют синтез ферментов – фосфолипазы А2 и циклооксигеназы – ключевых ферментов синтеза эйкозаноидов. Для ГК характерна супрессия активности иммунной системы, регуляция, воспалительных и аллергических реакций. Минералокортикоиды Альдостерон – основной минералокортикоид. Секреция гормона стимулируется ангитензином II – олигопептидом, формируемом в результате частичного протеолиза белка плазмы крови – ангиотензиногена Вначале под действием ренина из ангиотензиногена формируется ангиотензин I. Секреция самого ренина стимулируется при снижении фильтрационного давления в почках. Постоянная и независимая от почечного кровотока секреция ренина (базовая) поддерживается симпатической нервной системой. Далее ангиотензин I под действием ангиотензинпревращающего фермента (АПФ) превращается в ангиотензин II. Ангиотензин II связывается с поверхностью клеток клубочковой зоны коры надпочечников и стимулирует фосфорилирование регуляторного фермента синтеза стероидов - С21–22 десмолазы. Помимо активации секреции альдостерона ангиотензин II – мощный вазоконстриктор, также стимулятор центра жажды в головном мозге. Под действием минералокортикоидов повышается реабсорбция натрия, хлоридов и бикарбонатов в дистальных канальцах почек, а также увеличивается экскреция калия и водорода. Реализуя внутриклеточный механизм передачи гормонального сигнала, они стимулируют синтез Na+,K+-АТФаз на базальной мембране эпителиальных клеток дистальных канальцев и собирательных трубочек почек. Активируют синтез ферментов ЦТК и увеличение количества нарабатываемой в клетке энергии для работы Na+,K+-АТФаз. Формируют натриевые каналы для транспорта ионов через апикальную мембрану клеток почечного эпителия. Благодаря минералокортикоидам регулируется баланс электролитов и воды в организме, что влияет на величину артериального давления. Вызываемая ними гипернатриемия стимулирует в задней доле гипофиза секрецию вазопрессина (АДГ). Тот, в свою очередь, также усиливает спазм сосудов и свой уникальный эффект, стимулирующий реабсорбцию воды почками. Вместе с ангиотензинами и АДГ, эффекты альдостерона ведут к повышению системного артериального давления, следовательно, фильтрационное давление в нефронах увеличивается, поэтому секреция ренина прекращается (см. рис. 3), а давление стабилизируется. При стенозе почечной артерии даже при избыточно высоком системном давлении повышения фильтрационного давления не будет. Наблюдается избыточная секреция ренина и гиперактивность ренин-ангиотензиновой системы. Следовательно, для коррекции синдрома артериальной гипертензии таким пациентам показано применение ингибиторов АПФ. Андрогены коры надпочечников Андрогены надпочечников – представители группы мужских половых гормонов. Основными железами, синтезирующими андрогены, являются яички, в частности, клетки Лейдига. В них проходит синтез тестостерона из холестерол. Уникальным андрогеном коры является дегидроэпиандростерон (ДГЭА), синтезируемый в пучковой и сетчатой зонах из предшественника 17α-гидроксипрегненолона. Из 17α-гидроксипрогестерона или непосредственно из ДГЭА в коре формируется андростендион. Андрогенная активность надпочечниковых стероидов в основном обусловлена их способностью преобразовываться в тестостерон. В самих надпочечниках образуется очень мало тестостерона, преобладающими мужскими половыми гормонами являются дегидроэпиандростерон-сульфат (ДГЭА-С), ДГЭА и андростендион. Поэтому в биохмических исследованиях показателем продукции андрогенов в коре надпочечников считается именно ДГЭА. Кроме андрогенов, в коре надпочечников синтезируются в небольшом количестве женские половые гормоны, такие как прогестерон и его производные. Большое значение половые гормоны надпочечников имеют в детском возрасте, когда половые железы еще недостаточно активны. Благодаря половым гормонам коры надпочечников в этот период жизни стимулируется развитие половых признаков и полового поведения. Кроме того, данные гормоны являются анаболиками белкового обмена в скелетных мышцах, костной ткани и коже. Усилению эффектов тестостерона способствует его восстановление в периферических тканях 5α-редуктазой в дигидротестостерон (ДГТ). Наибольшая активность фермента зарегистрирована в простате, семенных пузыьках, в коже наружных половых органов. ДГТ обладает большим сродством к внутриклеточным рецепторам, связь их комплексов (гормон-рецептор) с ДНК стабильнее. Мишенями для тестостерона являются вольфовы структуры, сперматогонии, мышцы, кости, мозг, почки, кожа, связочный аппарат. Помимо контроля над формированием первичных половых признаков в эмбриогенезе, ДГТ и тестостерон ответственны за формирование вторичных половых признаков и сперматогенез. Мощный анаболический эффект андрогенов и стимуляция процессов репликации, транскрипции, трансляции в мышцах, костях, мозге, почках, повышение либидо – это прежде всего эффекты самого тестостерона. Конечный эффект тестостерона определяется его концентрацией, регулирующейся лютеинизирующим гормоном (ЛГ). Продукцию гонадолиберинов и самого тропного гормона тестостерон угнетает согласно механизмам отрицательной обратной связи. 1.2. Адреналин - гормон мозгового вещества надпочечников Мозговой слой надпочечников – часть вегетативной нервной системы. Его хромаффинные клетки реализуют продукцию катехоламинов, преимущественно адреналина (см. рис. 4). Аминокислота тирозин - непосредственный субстрат для их синтеза. Поэтому адреналин, как и йодтиронины щитовидной железы, классифицируется как гормон – производное аминокислоты тирозина  Адреналин, как и близкие ему по структуре дофамин и норадреналин, имеет катехоловое ядро, поэтому все трое составляют группу катехоламинов. Синтез фермента фенилэтаноламин-N-метилтрансферазы, метилирующей норадреналин в адреналин, стимулируют глюкокортикоиды. Следовательно, гормоны хронического стресса активируют синтез адреналина – гормона «острого стресса». Действительно, метаболические эффекты катехоламинов обеспечивают срочную адаптацию организма к резко возрастающей потребности в энергосубстаратах, связанной с его ответом на стресс, подобно эволюционным реакциям «бегства или нападения». Метаботропные рецепторы аденилатциклазного типа экспрессируют: печень, мышцы, жировая ткань. Стимулируя в гепатоцитах распад гликогена, адреналин реализует свое гипергликемическое действие. Оно обусловлено цАМФ-зависимой активацией фосфорилазы гликогена путем ее фосфорилирования. Одновременно фосфорилированию подвергается и гликогенсинтетаза, что сопровождается ее инактвацией. Следовательно, стимулируя β2-адренергические рецепторы в гепатоците, адреналин одновременно активирует гликогенолиз и подавляет гликогенез. В миоцитах обмен гликогена стимулируется подобным образом. Скорость липолиза в жировой ткани зависит от активности его ключевого фермента – ТАГ-липазы. Связываясь с β1-адренорецепторами адипоцитов, адреналин, подобно гепатоцитам, активирует в жировой ткани цАМФ-зависимую протеинкиназу, реализуя активацию ТАГ-липазы путем ее фосфорилирования. Помимо влияния на углеводный и липидный виды обмена, адреналин через стимуляцию β1-адренергических рецепторов увеличивает силу, частоту сердечных сокращений, минутный объем сердца. Через β2-адренергические рецепторы он реализует выраженное бронходилятирующее действие; расслабление гладких мышц матки, мочевого пузыря, сосудов, т.ч. коронарных. Противоположные эффекты на гладкие мышцы сосудов (вазоконстрикторные), на мышцы мочевого пузыря, матки катехоламины реализуют через α1-адренорецепторы. Их же активация в экзокринных железах (слюнных, потовых, аксиллярных, ладонных и подошвенных) позволяет адреналину усиливать секрецию слюны и пота. Вместе с норадреналином они через α1- и α2-адренорецепторы стимулируют вазоконстрикцию сосудов брюшной полости, кожи, слизистых оболочек. Перераспределение крови в сосудистом русле обеспечивает улучшение мозгового кровообращения, рост системного артериального давления. Период полураспада катехоламинов составляет 1 - 2 минуты. Их инактивируют катехол-О-метилтрансфераза (КОМТ) и моноаминооксидаза (МАО). МАО экспрессируется в печени, почках, желудке, кишечнике, нервной ткани, головном мозге, сердце, половых железах, надпочечниках, тромбоцитах. Описаны изоферменты МАО – МАО-А в нервной ткани и МАО-В – в других тканях. Натрийуретические факторы - антагонисты ренин-ангиотензиновой системы и альдостерона Считаем уместным вспомнить об антагонистичной альдостерону группе пептидных факторов, эффективно снижающей давление путем вазодилятации и регуляции обмена Na+ и K+. Это система атриопептинов - факторов, подавляющих реабсорбцию натрия в почках, стимулирующая, наоборот, его фильтрацию и выведение почками. Это т.н. натрийуретические факторы – предсердный, мозговой и сосудистый. Из них наиболее мощным гипотоником и калийсберегающим фактором является предсердный (предсердный атриопептин А, предсердный натрийуретический фактор А или ANPА). По сути - это гормон сердечного происхождения (вырабатывается секреторными кардиомиоцитами, сконцентрированными в ушках предсердий, желудочках сердца). Эти нейроэндокринные клетки синтезируют препрогормон, состоящий из 151 аминокислоты. В результате его частичного протеолиза крдиомиоциты могут формировать атриопептин А (из 28 аминокислот) и атриопептин В (состоит из 32 аминокислот). Натрийуретические факторы (полипептиды) синтезируются апудоцитами легких, мозга, вегетативных ганглиев. Однако их структуры подобны сердечным, группа пептидов называется атриопептинами. В частности, в мозге вырабатывется преимущественно атриопептин С (22 аминокислоты). Секреция атриопептинов возрастает пи гиперволемии, переходе в горизонтальное положение, солевой нагрузке, физической активности, при растяжении предсердий. Их секрецию стимулируют глюкокортикоиды, альдостерон, вазопрессин. Они же, наоборот, подавляют ренин-ангиотензин-альдостероновую систему, снижают продукцию АДГ (вазопрессина). Действуют на мишени (сосуды, почки, минералокортикоидную зону надпочесников) через мембранные рецепторы, представляющие трансмембранную гуанилатциклазу. Период их полураспада или эффективной циркуляции в крови составляет 3 – 5 минут. |