экзамен биология 2013. I. Введение и общая характеристика жизни
 Скачать 1.53 Mb. Скачать 1.53 Mb.
|

|
Функциями лизосом являются: переваривание захваченных клеткой при эндоцитозе веществ или частиц (бактерий, других клеток) аутофагия — уничтожение ненужных клетке структур, например, во время замены старых органоидов новыми, или переваривание белков и других веществ, произведенных внутри самой клетки автолиз — самопереваривание клетки, приводящее к ее гибели (иногда этот процесс не является патологическим, а сопровождает развитие организма или дифференцировку некоторых специализированных клеток). Пример: При превращении головастика в лягушку, лизосомы, находящиеся в клетках хвоста, переваривают его: хвост исчезает, а образовавшиеся во время этого процесса вещества всасываются и используются другими клетками тела. растворение внешних структур (см, например, остеокласты) - Пероксисомы - небольшие (размером 0,3—1,5 мкм) овальной формы тельца, ограниченные мембраной, содержащие гранулярный матрикс, в центре которого часто видны кристаллоподобные структуры, состоящие из фибрилл и трубочек. Пероксисомы особенно характерны для клеток печени и почек. Во фракции пероксисом обнаруживаются ферменты окисления аминокислот, при работе которых образуется перекись водорода. то универсальный мембранный органоид клетки, диаметром примерно 0,15-0,25нм. Главной функцией пероксисом является расщепление длиннорадикальных жирных кислот. Хотя в целом они могут выполнять и другие функции. Пероксисомы в клетке образуются только за счет деления материнских пероксисом, поэтому , если в клетку по каким-то причинам не попали пероксисомы, то клетка погибает из-за накопления жирных кислот. Мембрана пероксисом имеет типичное жидкостно-мозаичное строение и может увеличиваться за счет переносимых сюда специальными белками переносчиками сложных липидов и белков. Функции. -Расщепление жирных кислот. В пероксисомах содержаться ферменты , относящиеся к группе ферментов оксидоредуктаз, которые начинают расщепление жирных кислот с отщепления остатков уксусной кислоты и образуют внутри радикала жирной кислоты двойную связь и как побочный продукт образуется перекись водорода.. -Пероксисомы участвуют в детоксикации многих вредных веществ, например, спиртов, альдегидов и кислот. Эта функция характерна для клеток печени, причем пероксисомы в печени имеют более крупные размеры. Детоксикация ядов веществ происходит за счет их окисления. -Пероксисомы способны катализировать окисление уратов, т.к. в них находится фермент уратоксидаза. 11. Аппарат Гольджи — важная органелла, которая присутствует практически в каждой эукариотической клетке. Пожалуй, единственными клетками, в которых отсутствует этот комплекс, являются эритроциты позвоночных животных. Функции этой структуры весьма разнообразны. Именно в цистернах аппарата скапливают все вырабатываемые клеткой соединения, после чего происходит их дальнейшая сортировка, модификация, перераспределение и транспорт. Строение- собой совокупность мембранных цистерн, которые тесно прилегают друг к другу, напоминая стопку. Структурное и функциональной единицей здесь считается диктиосома. Диктиосома самостоятельная часть аппарата Гольджи, которая состоит из 3 – 8 тесно прилегающих друг к другу цистерн. Стопка этих мембранных цистерн окружена системой мелкий вакуолей и пузырьков — именно таким образом осуществляется транспорт веществ, а также связь диктиосом между собой и другими клеточными структурами. Как правило, животные клетки имеют только одну диктиосому, в то время как в растительных структурах их может быть много. В диктиосоме принято разделять два конца — цис- и транс-стороны. Цис–сторона обращена в сторону ядра и гранулярной эндоплазматической сетки. Сюда в виде мембранных пузырьков транспортируются синтезированные белки и другие соединения. На этом конце диктиосомы постоянно образуются новые цистерны. Функции-Здесь осуществляется модификация новосинтезированных белковых молекул. В большинстве случаев к протеиновой молекуле присоединяется углеводный, сульфатный или фосфорный радикал. Таким образом, аппарат Гольджи отвечает за формирование белкой плазматической мембраны, ферментов и белков лизосом. -Аппарат Гольджи отвечает за транспорт модифицированных белков в определенные участки клетки. -От транс-стороны постоянно отделяются небольшие пузырьки, в которых содержатся готовые протеины. -Здесь происходит образование и транспорт всех ферментов лизосом. -В полостях цистерн происходит накопление липидов, а в дальнейшем и образование липопротеидов — комплекса белковой и липидной молекулы. -Аппарат Гольджи растительной клетки отвечает за синтез полисахаридов, которые затем идут на образование клеточной стенки растения, а также слизи, пектинов, гемицеллюлозы и восков. После деления растительной клетки комплекс Гольджи берет участие в формировании клеточной пластинки. В сперматозоиде эта органелла берет участие в образовании ферментов акросомы, с помощью которых происходит разрушение оболочек яйцеклетки при оплодотворении. В клетках представителей простейших комплекс Гольджи отвечает за образование сократительных вакуолей, которые регулируют осмотическое давление 12. - Митохондрии - полуавтономные двумембранные структуры продолговатой формы. Наружная мембрана гладкая, а внутренняя имеет складки - кристы, увеличивающие ее поверхность. Внутри митохондрия заполнена матриксом, в котором находятся кольцевая молекула ДНК, РНК, рибосомы. Количество митохондрий в клетках различно, с ростом клеток их число увеличивается в результате деления. Митохондрии — это «энергетические станции» клетки. В процессе дыхания в них происходит окончательное окисление веществ кислородом воздуха. Выделяющаяся энергия запасается в молекулах АТФ, синтез которых происходит в этих структурах. Пластиды являются основными цитоплазматическими органеллами клеток автотрофных растений. Название происходит от греческого слова «plastos», что в переводе означает «вылепленный». Главная функция пластид – синтез органических веществ, благодаря наличию собственных ДНК и РНК и структур белкового синтеза. В пластидах также содержатся пигменты, обусловливающие их цвет. Все виды данных органелл имеют сложное внутреннее строение. Снаружи пластиду покрывают две элементарные мембраны, имеется система внутренних мембран, погруженных в строму или матрикс. Классификация пластид по окраске и выполняемой функции подразумевает деление этих органоидов на три типа: хлоропласты, лейкопласты и хромопласты. Пластиды водорослей именуются хроматофорами. Хлоропласты – это зеленые пластиды высших растений, содержащие хлорофилл – фотосинтезирующий пигмент. Представляют собой тельца округлой формы размерами от 4 до 10 мкм. Химический состав хлоропласта: примерно 50% белка, 35% жиров, 7% пигментов, малое количество ДНК и РНК. У представителей разных групп растений комплекс пигментов, определяющих окраску и принимающих участие в фотосинтезе, отличается. Это подтипы хлорофилла и каротиноиды (ксантофилл и каротин). При рассматривании под световым микроскопом видна зернистая структура пластид – это граны. Под электронным микроскопом наблюдаются небольшие прозрачные уплощенные мешочки (цистерны, или граны), образованные белково-липидной мембраной и располагающиеся в непосредственно в строме. Причем некоторые из них сгруппированы в пачки, похожие на столбики монет (тилакоиды гран), другие, более крупные находятся между тилакоидами. Благодаря такому строению, увеличивается активная синтезирующая поверхность липидно-белково-пигментного комплекса гран, в котором на свету происходит фотосинтез. Хромопласты – пластиды, окраска которых бывает желтого, оранжевого или красного цвета, что обусловлено накоплением в них каротиноидов. Благодаря наличию хромопластов, характерную окраску имеют осенние листья, лепестки цветов, созревшие плоды (помидоры, яблоки). Данные органоиды могут быть различной формы – округлой, многоугольной, иногда игольчатой. Лейкопласты представляют собой бесцветные пластиды, основная функция которых обычно запасающая. Размеры этих органелл относительно небольшие. Они округлой либо слегка продолговатой формы, характерны для всех живых клеток растений. В лейкопластах осуществляется синтез из простых соединений более сложных – крахмала, жиров, белков, которые сохраняются про запас в клубнях, корнях, семенах, плодах. Под электронным микроскопом заметно, что каждый лейкопласт покрыт двухслойной мембраной, в строме есть только один или небольшое число выростов мембраны, основное пространство заполнено органическими веществами. В зависимости от того, какие вещества накапливаются в строме, лейкопласты делят на амилопласты, протеинопласты и элеопласты. Все виды пластид имеют общее происхождение и способны переходить из одного вида в другой. Так, превращение лейкопластов в хлоропласты наблюдается при позеленении картофельных клубней на свету, а в осенний период в хлоропластах зеленых листьев разрушается хлорофилл, и они трансформируются в хромопласты, что проявляется пожелтением листьев. В каждой определенной клетке растения может быть только один вид пластид. 13. Немембранные органеллы: 1) Рибосомы - мелкие тельца грибовидной формы, в которых идет синтез белка. Они состоят из рибосомальной РНК и белка, образующего большую и малую субъединицы. 2) Цитоскелет - опорно-двигательная система клетки, включающая немембранные образования, выполняющие как каркасную, так и двигательную функции в клетке. Эти нитчатые или фибриллярные могут быстро возникать и так же быстро исчезать. К этой системе относятся фибриллярные структуры(5-7нм) и микротрубочки (состоят из 13 субъединиц). 3) Клеточный центр состоит из центриолей (длинна 150нм, диаметр 300-500 нм), окруженных центросферами. Центриоли состоят из 9 триплетов микротрубочек. Функции: - образование нитей митотического веретена деления. – Обеспечение расхождения сестринских хроматид в анафазе митоза. 14. Ядро – структура, обеспечивающая генетическую детерминацию и регуляцию белкового синтеза. Строение: ядерная оболочка, хроматин, ядерный сок, ядрышко. В ядрышке выделяют нитчатый и зернистый компоненты. Нитчатый компонент состоит из белка и гигантской РНК – предшественницы, которая затем образует более мелкие рРНК. В процессе созревания фибриллы преобразуются в зёрна (гранулы). Функции: обеспечивает образование и созревание рРНК. 15. В среднем в хроматине около 40% приходится на ДНК и около 60% - на белки, среди которых специфические ядерные белки-гистоны составляют от 40 до 80% от всех белков, входящих в состав выделенного хроматина. Кроме того, в состав хроматиновой фракциям входят мембранные компоненты, РНК, углеводы, липиды, гликопротеиды. Вопрос о том, насколько эти минорные компоненты входят в структуру хроматина, еще не решен. Так, РНК может представлять собой транскрибируемую РНК, которая еще не потеряла связь с матрицей ДНК. Другие же минорные компоненты могут относиться к веществам соосажденных фрагментов ядерной оболочки. Упаковка ДНК в хроматин обеспечивает многократное сокращение линейных размеров ДНК, необходимое для размещения её в ядре. При этом надо оставить доступными определённые последовательности ДНК для регуляторных факторов и ферментов транскрипции. Эти задачи решаются на уровне упаковки ДНК в хроматин, которая происходит в несколько этапов. Наиболее изученными является три первых уровня упаковки: (1) накручивание ДНК на нуклеосомы с образованием нуклеосомной нити диаметром 10 нм, (2) компактизация нуклеосомной нити с образованием так называемой 30-нм фибриллы и (3) сворачивание последней в гигантские (50 — 200 тысяч п. н.) петли, закреплённые на белковой скелетной структуре ядра — ядерном матриксе[15]. Одним из самых последних уровней упаковки в митотическую хромосому некоторые исследователи считают уровень так называемой хромонемы, толщина которой составляет около 0,1 — 0,3 мкм[16]. В результате дальнейшей компактизации диаметр хроматиды достигает ко времени метафазы 700 нм. Значительная толщина хромосомы (диаметр 1400 нм) на стадии метафазы позволяет, наконец, увидеть её в световой микроскоп. Конденсированная хромосома имеет вид буквы X (часто с неравными плечами), поскольку две хроматиды, возникшие в результате репликации, соединены между собой в районе центромеры (подробнее о судьбе хромосом при клеточном делении см. статьи митоз и мейоз). 16. Сохраняя преемственность в ряду клеточных поколений, хромосомы в зависимости от периода и фазы клеточного цикла меняют свое строение. В интерфазе они образуют ядерные структуры, названные в связи с их способностью прокрашиваться основными красителями хроматином. При переходе клетки к митозу, особенно в метафазе, хроматин приобретает вид хорошо различимых отдельных интенсивно окрашенных телец — хромосом. Наиболее распространенной является точка зрения, согласно которой хроматин (хромосома) представлен спирализованными нитями. При этом выделяется несколько уровней спирализации (компактизации) хроматина. В зависимости от состояния хроматина выделяют эухроматиновые участки хромосом, отличающиеся меньшей плотностью упаковки в неделящихся клетках и потенциально транскрибируемые, и гетерохроматиновые участки, характеризующиеся компактной организацией и генетической инертностью. В их пределах транскрипции биологической информации не происходит.Морфология хромосом. Метафазная хромосома состоит из двух продольных нитей ДНП — хроматид, соединенных друг с другом в области первичной перетяжки (центромеры). 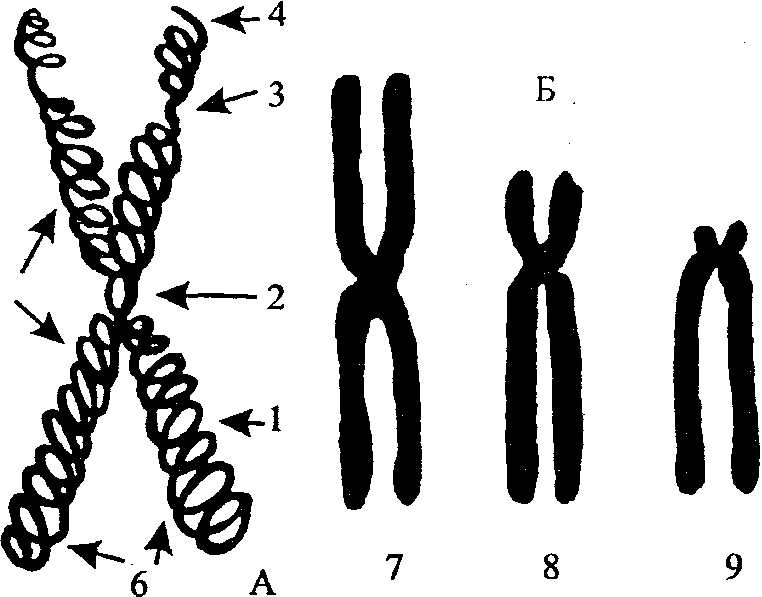 Рис. 9.1 . Схема строения метафазной хромосомы (А) и типы хромосом (Б). А: 1 — плечо, 2 — центромера, 3 — вторичная перетяжка, 4 — спутник, 5 — хроматиды, 6 — теломеры. Б: 7 — метацентрическая, 8 — субметацентрическая, 9 — акроцентрическая хромосомы. Уровни укладки хромосом Резкое укорочение длины хромосом при конденсации достигается за счёт нескольких уровней их укладки.
17. Конститутивный гетерохроматин содержится в околоцентромерных и теломерных участках всех хромосом, а также на протяжении некоторых внутренних фрагментов отдельных хромосом Он образован только нетранскрибируемой ДНК. Вероятно, его роль заключается в поддержании общей структуры ядра, прикреплении хроматина к ядерной оболочке, взаимном узнавании гомологичных хромосом в мейозе, разделении соседних структурных генов, участии в процессах регуляции их активности. Примером факультативного гетерохроматина служит тельце полового хроматина, образуемое в норме в клетках организмов гомогаметного пола (у человека гомогаметным является женский пол) одной из двух Х-хромосом. Гены этой хромосомы не транскрибируются. Образование факультативного гетерохроматина за счет генетического материала других хромосом сопровождает процесс клеточной дифференцировки и служит механизмом выключения из активной функции групп генов, транскрипция которых не требуется в клетках данной специализации. В связи с этим рисунок хроматина ядер клеток из разных тканей и органов на гистологических препаратах различается. Примером может служить гетерохроматизация хроматина в ядрах зрелых эритроцитов птиц. Перечисленные уровни структурной организации хроматина обнаруживаются в неделящейся клетке, когда хромосомы еще недостаточно компактизованы, чтобы быть видимыми в световой микроскоп как отдельные структуры. Лишь некоторые их участки с более высокой плотностью упаковки выявляются в ядрах в виде хроматиновых глыбок Компактные участки гетерохроматина сгруппированы около ядрышка и ядерной мембраны Метафазная хромосома. Вступление клетки из интерфазы в митоз сопровождается суперкомпактизацией хроматина. Отдельные хромосомы становятся хорошо различимы. Этот процесс начинается в профазе, достигая своего максимального выражения в метафазе митоза и анафазе (см. разд. 2.4.2). В телофазе митоза происходит декомпак-тизация вещества хромосом, которое приобретает структуру интерфазного хроматина. Описанная митотическая суперкомпактизация облегчает распределение хромосом к полюсам митотического веретена в анафазе митоза 18. хромосома состоит из двух плеч и центромеры, или первичной перетяжки, выполняющей функцию механического центра хромосомы при делении. Центромера является областью хромосомы, к которой при делении клетки прикрепляется нить веретена деления, разводящая хромосомы к полюсам клетки. Кроме первичной перетяжки некоторые хромосомы имеют вторичную перетяжку, не связанную с процессом прикрепления нитей веретена. Месторасположение вторичной перетяжки в хромосоме связано с образованием ядрышка, а этот участок хромосомы называют ядрышковым организатором. Длинное плечо хромосомы обозначается латинской буквой «q», короткое — «р». Концевые участки хромосом имеют сегменты, препятствующие склеиванию хромосом своими концами, и тем самым способствуют сохранению их целостности. Эти сегменты были названы теломерами. Если центромера расположена в центре хромосомы и как бы делит ее на равные части, то такая хромосома носит название равноплечной, или метацентрической. Если одно плечо незначительно больше другого — субметацентрической. Хромосому с почти терминальным (концевым) положением центромеры, когда одно плечо значительно больше другого, называют акроцентрической. Некоторые акроцентрические хромосомы имеют так называемые спутники — участки, соединенные с остальной частью хромосомы тонкой нитью хроматина. Такие хромосомы называют спутничковыми. Размер спутника относительно длины всей хромосомы постоянен для каждой конкретной хромосомы. В кариотипе человека спутники имеются у пяти пар хромосом: у 13-й, 14, 15, 21 и 22-й. |