ЭКЗАМЕН по Биологии вопросы 29-42. Исторический очерк
 Скачать 0.62 Mb. Скачать 0.62 Mb.
|

|
Цитогенетика кроссинговера Механизм кроссинговера «разрыв–воссоединение» Согласно теории Янссенса–Дарлингтона, кроссинговер происходит в профазе мейоза. Гомологичные хромосомы с гаплотипами хроматид АВ и ab образуют биваленты. В одной из хроматид в первой хромосоме происходит разрыв на участке А–В, тогда в прилежащей хроматиде второй хромосомы происходит разрыв на участке a–b. Клетка стремится исправить повреждение с помощью ферментов репарации–рекомбинации и присоединить фрагменты хроматид. Однако при этом возможно присоединение крест–накрест (кроссинговер), и образуются рекомбинантные гаплотипы (хроматиды) Abи аВ. В анафазе первого деления мейоза происходит расхождение двухроматидных хромосом, а во втором делении – расхождение хроматид (однохроматидных хромосом). Хроматиды, которые не участвовали в кроссинговере, сохраняют исходные сочетания аллелей. Такие хроматиды (однохроматидные хромосомы) называются некроссоверными; с их участием разовьются некроссоверные гаметы, зиготы и особи. Рекомбинантные хроматиды, которые образовались в ходе кроссинговера, несут новые сочетания аллелей. Такие хроматиды (однохроматидные хромосомы) называются кроссоверными, с их участием разовьются кроссоверные гаметы, зиготы и особи. Таким образом, вследствие кроссинговера происходит рекомбинация – появление новых сочетаний (гаплотипов) наследственных задатков в хромосомах. 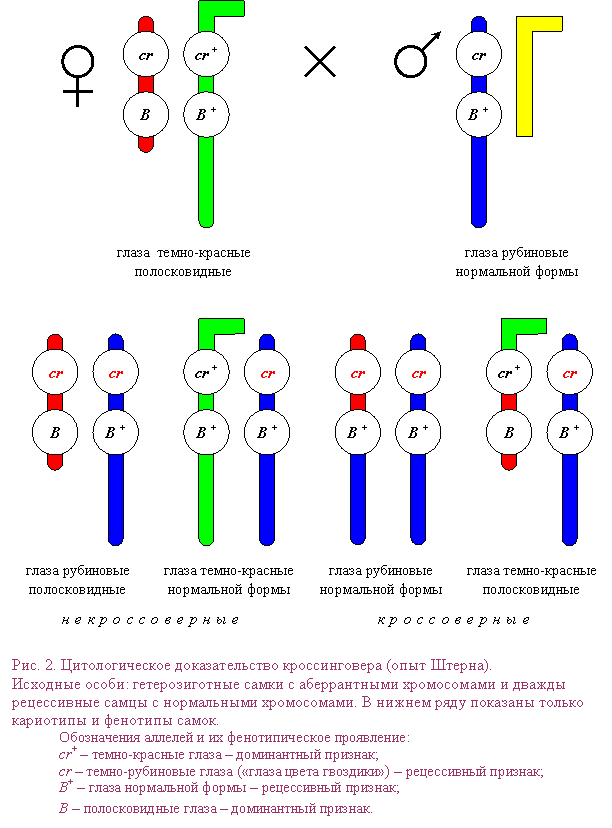 Примечание. Согласно другим теориям, кроссинговер связан с репликацией ДНК: или в пахитене мейоза, или в интерфазе (см. ниже). В частности, возможна смена матрицы в вилке репликации. Интерференция – это подавление кроссинговера на участках, непосредственно прилегающих к точке происшедшего обмена. Рассмотрим пример, описанный в одной из ранних работ Моргана. Он исследовал частоту кроссинговера между генами w (white – белые глаза), у (yellow – желтое тело) и m (miniature – маленькие крылья), локализованными в Х-хромосоме D. melanogaster.Расстояние между генами w и у в процентах кроссинговера составило 1,3, а между генами у и m – 32,6. Если два акта кроссинговера наблюдаются случайно, то ожидаемая частота двойного кроссинговера должна быть равна произведению частот кроссинговера между генами у и w и генами w и m. Другими словами, частота двойных кроссинговеров будет 0,43%. В действительности в опыте был обнаружен лишь один двойной кроссинговер на 2205 мух, т. е. 0,045%. Ученик Моргана Г. Меллер предложил определять интенсивность интерференции количественно, путем деления фактически наблюдаемой частоты двойного кроссинговера на теоретически ожидаемую (при отсутствии интерференции) частоту. Он назвал этот показатель коэффициентом коинциденции, т. е. совпадения. Меллер показал, что в Х-хромосоме дрозофилы интерференция особенно велика на небольших расстояниях; с увеличением интервала между генами интенсивность ее уменьшается и на расстоянии около 40 морганид и более коэффициент коинциденции достигает 1 (максимального своего значения). Двойной и множественный кроссинговер. Т. Морган предположил, что кроссинговер между двумя генами может происходить не только в одной, но и в двух и даже большем числе точек. Четное число перекрестов между двумя генами, в конечном счете, не приводит к их перемещению из одной гомологичной хромосомы в другую, поэтому число кроссинговеров и, следовательно, расстояние между этими генами, определенное в эксперименте, снижаются. Обычно это относится к достаточно далеко расположенным друг от друга генам. Естественно, что вероятность двойного перекреста всегда меньше вероятности одинарного. В принципе она будет равна произведению вероятности двух единичных актов рекомбинации. Например, если одиночный перекрест будет происходить с частотой 0,2, то двойной – с частотой 0,2 × 0,2 = 0,04. В дальнейшем, наряду с двойным кроссинговером, было открыто и явление множественного кроссинговера: гомологичные хроматиды могут обмениваться участками в трех, четырех и более точках. Цитологическое (цитогенетическое) доказательство кроссинговера. Прямые цитологические свидетельства обмена частей хромосом во время кроссинговера были получены в начале 30-х годов у дрозофилы и кукурузы. Рассмотрим опыт Штерна, проведенный на D. melanogaster. Обычно две гомологичные хромосомы морфологически неразличимы. Штерн исследовал Х-хромосомы, которые имели морфологические различия и, следовательно, были гомологичны не полностью. Однако гомология между этими хромосомами сохранялась на большей части их длины, что позволяло им нормально спариваться и сегрегировать в мейозе (то есть нормально распределяться по дочерним клеткам). Одна из Х-хромосом самки в результате транслокации, т. е. перемещения фрагмента Y-хромосомы, приобрела Г-образную форму. Вторая Х-хромосома была короче нормальной, так как часть ее была перенесена на IV хромосому. Были получены самки, гетерозиготные по указанным двум, морфологически различным, Х-хромосомам, а также гетерозиготные по двум генам, локализованным в Х-хромосоме: Bar (В) иcarnation (cr). Ген Bar – это полудоминантный ген, влияющий на количество фасеток и, следовательно, форму глаза (мутанты с аллелем В имеют полосковидные глаза). Ген cr контролирует окраску глаз (аллель cr+ обусловливает нормальную окраску глаз, а аллель cr – окраску глаз цвета красной гвоздики). Г-образная Х-хромосома несла аллели дикого типа В+ и cr+, укороченная хромосома – мутантные аллели В и cr. Самки указанного генотипа скрещивались с самцами, имевшими морфологически нормальную Х-хромосому с аллелями cr и В+. В потомстве самок было два класса мух с некроссоверными хромосомами (crB / crB+ и cr+B+ /crB+) и два класса мух, фенотип которых соответствовал кроссоверам (crB+ / crB+ и cr+B / crB+). Цитологическое исследование показало, что у кроссоверных особей произошел обмен участками Х-хромосом, и, соответственно, изменилась их форма. Все четыре класса самок имели по одной нормальной, т. е. палочковидной, хромосоме, полученной от отца. Кроссоверные самки содержали в своем кариотипе преобразованные в результате кроссинговера Х-хромосомы – длинную палочковидную или двуплечую с короткими плечами. Эти опыты, так же как и одновременно полученные аналогичные результаты на кукурузе, подтвердили гипотезу Моргана и его сотрудников о том, что кроссинговер представляет собой обмен участками гомологичных хромосом и что гены действительно локализованы в хромосомах. Генетические и цитогенетические карты хромосом Генетические карты хромосом схемы относительного расположения сцепленных между собой наследственных факторов - генов. Генетические карты хромосом отображают реально существующий линейный порядок размещения генов в хромосомах (см. Цитологические карты хромосом) и важны как в теоретических исследованиях, так и при проведении селекционной работы, т.к. позволяют сознательно подбирать пары признаков при скрещиваниях, а также предсказывать особенности наследования и проявления различных признаков у изучаемых организмов. ИмеяГенетические карты хромосом можно по наследованию «сигнального» гена, тесно сцепленного с изучаемым, контролировать передачу потомству генов, обусловливающих развитие трудно анализируемых признаков; например, ген, определяющий эндосперм у кукурузы и находящийся в 9-й хромосоме, сцеплен с геном, определяющим пониженную жизнеспособность растения. Многочисленные факты отсутствия (вопреки Менделя законам) независимого распределения признаков у гибридов второго поколения были объяснены хромосомной теорией наследственности. Гены, расположенные в одной хромосоме, в большинстве случаев наследуются совместно и образуют одну группу сцепления, количество которых, т. о., соответствует у каждого организма гаплоидному числу хромосом (см. Гаплоид). Американский генетик Т. Х. Морган показал, однако, что сцепление генов, расположенных в одной хромосоме, у диплоидных организмов (см. Диплоид) не абсолютное; в некоторых случаях перед образованием половых клеток между однотипными, или гомологичными, хромосомами происходит обмен соответственными участками; этот процесс носит название перекреста, или кроссинговера. Обмен участками хромосом (с находящимися в них генами) происходит с различной вероятностью, зависящей от расстояния между ними (чем дальше друг от друга гены, тем выше вероятность кроссинговера и, следовательно, рекомбинации). Генетический анализ позволяет обнаружить перекрест только при различии гомологичных хромосом по составу генов, что при кроссинговере приводит к появлению новых генных комбинаций. Обычно расстояние между генами на Генетические карты хромосом выражают как % кроссинговера (отношение числа мутантных особей, отличающихся от родителей иным сочетанием генов, к общему количеству изученных особей); единица этого расстояния - морганида - соответствует частоте кроссинговера в 1%. Генетические карты хромосом составляют для каждой пары гомологичных хромосом. Группы сцепления нумеруют последовательно, по мере их обнаружения. Кроме номера группы сцепления, указывают полные или сокращённые названия мутантных генов, их расстояния в морганидах от одного из концов хромосомы, принятого за нулевую точку, а также место центромеры. Составить Генетические карты хромосом можно только для объектов, у которых изучено большое число мутантных генов. Например, у дрозофилы идентифицировано свыше 500 генов, локализованных в её 4 группах сцепления, у кукурузы - около 400 генов, распределенных в 10 группах сцепления (рис. 1). У менее изученных объектов число обнаруженных групп сцепления меньше гаплоидного числа хромосом. Так, у домовой мыши выявлено около 200 генов, образующих 15 групп сцепления (на самом деле их 20); у кур изучено пока всего 8 из 39. У человека из ожидаемых 23 групп сцепления (23 пары хромосом) идентифицировано только 10, причём в каждой группе известно небольшое число генов; наиболее подробные карты составлены для половых хромосом. У бактерий, которые являются гаплоидными организмами, имеется одна, чаще всего непрерывная, кольцевая хромосома и все гены образуют одну группу сцепления (рис. 2). При переносе генетического материала из клетки-донора в клетку-реципиент, например приконъюгации, кольцевая хромосома разрывается и образующаяся линейная структура переносится из одной бактериальной клетки в другую (у кишечной палочки в течение 110-120 мин). Искусственно прерывая процесс конъюгации, можно по возникшим типам рекомбинантов установить, какие гены успели перейти в клетку-реципиент. В этом состоит один из методов построения Генетические карты хромосом бактерий, детально разработанных у ряда видов. Ещё более детализированы Генетические карты хромосом некоторых бактериофагов. См. также Генетика,Мутация. Цитологические карты хромосом, схематическое изображение хромосом с указанием мест фактического размещения отдельных генов, полученное с помощью цитологических методов. Цитологические карты хромосом составляют для организмов, для которых обычно уже имеются генетические карты хромосом. Каждое место расположения гена (локус) на генетической карте организма, установленное на основе частоты перекреста участков хромосом (кроссинговера), на Цитологические карты хромосом привязано к определённому, реально существующему участку хромосомы, что служит одним из основных доказательств хромосомной теории наследственности. Для построенияЦитологические карты хромосом используют данные анализа хромосомных перестроек (вставки, делеции и др.) и, сопоставляя изменения морфологических признаков хромосом при этих перестройках с изменениями генетических свойств организма, устанавливают место того или иного гена в хромосоме. Пользуясь методом хромосомных перестроек, амер. генетик К. Бриджес составил в 1935 подробную Цитологические карты хромосом плодовой мушки дрозофилы, наиболее полно генетически изученного организма. Гигантские хромосомы насекомых отряда двукрылых оказались самыми удобными для построения Цитологические карты хромосом т.к. наряду с большими размерами обладают чёткой морфологической очерченностью: каждый участок этих хромосом имеет свой определённый и чёткий рисунок, обусловленный характерным чередованием по длине ярко окрашиваемых участков (дисков) и слабо окрашиваемых (междисков). Цитологическими методами легко определить отсутствие участка хромосомы или перенос его в др. место. Сопоставление Цитологические карты хромосом с генетическими показало, что физическое расстояние между генами в хромосомах не соответствует генетическому (видимо, частота кроссинговера неодинакова в разных участках хромосом), поэтому плотность распределения генов на цитологических и генетических картах хромосом различна. Так было установлено важное генетическое явление — неравномерность частот перекреста по длине хромосомы. Линейное расположение генов и их последовательность, установленные генетическими методами, подтверждаются Цитологические карты хромосом Современные методы цитологии и генетики позволяют построить Цитологические карты хромосом многих организмов, в том числе человека. Вопрос 34 (Хромосомная теория наследственности) Хромосомная теория наследственности, теория, согласно которой хромосомы, заключённые в ядре клетки, являются носителями генов и представляют собой материальную основу наследственности, т.е. преемственность свойств организмов в ряду поколений определяется преемственностью их хромосом. Хромосомная теория наследственности возникла в начале 20 в. на основе клеточной теории и использования для изучения наследственных свойств организмов гибридологического анализа. В 1902 У. Сеттон в США, обративший внимание на параллелизм в поведении хромосом и менделевских т. н. «наследственных факторов», и Т.Бовери в Германии выдвинули хромосомную гипотезу наследственности, согласно которой менделевские наследственные факторы (название впоследствии генами) локализованы в хромосомах. Первые подтверждения этой гипотезы были получены при изучении генетического механизма определения пола у животных, когда было выяснено, что в основе этого механизма лежит распределение половых хромосом среди потомков. Дальнейшее обоснование Хромосомная теория наследственности принадлежит американскому генетику Т. Х. Моргану, который заметил, что передача некоторых генов (например, гена, обусловливающего белоглазие у самок дрозофилы при скрещивании с красноглазыми самцами) связана с передачей половой Х-хромосомы, т. е. что наследуются признаки, сцепленные с полом (у человека известно несколько десятков таких признаков, в том числе некоторые наследственные дефекты - дальтонизм, гемофилия и др.). Доказательство Хромосомная теория наследственности было получено в 1913 американским генетиком К. Бриджесом, открывшим нерасхождение хромосом в процессе мейоза у самок дрозофилы и отметившим, что нарушение в распределении половых хромосом сопровождается изменениями в наследовании признаков, сцепленных с полом. С развитием Хромосомная теория наследственности было установлено, что гены, расположенные в одной хромосоме, составляют одну группу сцепления (см. Сцепление генов) и должны наследоваться совместно; число групп сцепления равно числу пар хромосом, постоянному для каждого вида организмов (см. Кариотип); признаки, зависящие от сцепленных генов, также наследуются совместно. Вследствие этого закон независимого комбинирования признаков (см. Менделя законы) должен иметь ограниченное применение; независимо должны наследоваться признаки, гены которых расположены в разных (негомологичных) хромосомах. Явление неполного сцепления генов (когда наряду с родительскими сочетаниями признаков в потомстве от скрещиваний обнаруживаются и новые, рекомбинантные, их сочетания) было подробно исследовано Морганом и его сотрудниками (А. Г. Стёртевантом и др.) и послужило обоснованием линейного расположения генов в хромосомах. Морган предположил, что сцепленные гены гомологичных хромосом, находящиеся у родителей в сочетаниях Согласно представлениям школы Моргана, гены являются дискретными и далее неделимыми носителями наследственной информации. Однако открытие в 1925 советскими учёными Г. А. Надсоном и Г. С. Филипповым, а в 1927 американским учёным Г. Мёллером влияния рентгеновских лучей на возникновение наследственных изменений (мутаций) у дрозофилы, а также применение рентгеновских лучей для ускорения мутационного процесса у дрозофилы позволили советским учёным А. С. Серебровскому, Н. П. Дубинину и др. сформулировать в 1928-30 представления о делимости гена на более мелкие единицы, расположенные в линейной последовательности и способные к мутационным изменениям. В 1957 эти представления были доказаны работой американского учёного С. Бензера с бактериофагом Т4. Использование рентгеновских лучей для стимулирования хромосомных перестроек позволило Н. П. Дубинину и Б. Н. Сидорову обнаружить в 1934 эффект положения гена (открытый в 1925 Стёртевантом), т. е. зависимость проявления гена от места расположения его на хромосоме. Возникло представление о единстве дискретности и непрерывности в строении хромосомы. Хромосомная теория наследственности развивается в направлении углубления знаний об универсальных носителях наследственной информации - молекулах дезоксирибонуклеиновой кислоты (ДНК). Установлено, что непрерывная последовательность пуриновых и пиримидиновых оснований вдоль цепи ДНК образует гены, межгенные интервалы, знаки начала и конца считывания информации в пределах гена; определяет наследственный характер синтеза специфических белков клетки и, следовательно, наследственный характер обмена веществ. ДНК составляет материальную основу группы сцепления у бактерий и многих вирусов (у некоторых вирусов носителем наследственной информации является рибонуклеиновая кислота); молекулы ДНК, входящие в состав митохондрий, пластид и др. органоидов клетки, служат материальными носителями цитоплазматической наследственности. Хромосомная теория наследственности, объясняя закономерности наследования признаков у животных и растительных организмов, играет важную роль в с.-х. науке и практике. Она вооружает селекционеров методами выведения пород животных и сортов растений с заданными свойствами. Некоторые положения Хромосомная теория наследственности позволяют более рационально вести с.-х. производство. Так, явление сцепленного с полом наследования ряда признаков у с.-х. животных позволило до изобретения методов искусственного регулирования пола у тутового шелкопряда выбраковывать коконы менее продуктивного пола, до разработки способа разделения цыплят по полу исследованием клоаки - отбраковывать петушков и т.п. Важнейшее значение для повышения урожайности многих с.-х. культур имеет использование полиплоидии.На знании закономерностей хромосомных перестроек основывается изучение наследственных заболеваний человека. Вопрос 35(Наследования признаков человека сцепленных с полом) Известно, что Х-хромосомы генетически активны, как любая из аутосом. У-хромосомы — генетически инертны, у человека лишь некоторые гены, не являющиеся жизненно важными, локализуются в ней (гипертрихоз — волосатые уши, некоторые законы передачи признаков, сцепленных с X хромосомами, были впервые изучены Т.Морганом на дрозофилах. Кариотип самца дрозофилы несет пару различных хромосом, получивших название половых (ХУ), а самки — пару одинаковых половых хромосом — XX. Самки образуют только один тип гамет — X, а самцы два типа гамет Хи У. Ген, определяющий белый цвет глаз, находится у дрозофил в Х-хромосоме (рецессивный аллель), доминантный — формирует красные глаза. При скрещивании мух с белыми глазами (white) — «w» и мух с обычными темно-красными глазами — W+ были обнаружены различия результатов реципрокных скрещиваний. Два скрещивания, которые характеризуются взаимно противоположным сочетанием анализируемого признака и пола, называются реципрокными скрещиваниями. При скрещивании красноглазой самки и белоглазого самца (гомогаметный пол доминантен) в F, все мухи были красноглазые, а в F2 происходило расщепление 3 красноглазых : I белоглазой. Значит, если гомогаметный пол доминантен, то соблюдаются все законы Менделя: единообразие первого поколения и расщепление во втором поколении по фенотипу 3:1. При реципрокном скрещивании, когда гомогаметный пол был рецессивен (белоглазая самка и красноглазый самец), в первом поколении наблюдались иные результаты: все самки в F, были красноглазыми, а все самцы — белоглазыми. Такое наследование получило название крисс-кросс (или крест-накрест) наследования, сыновья наследуют фенотипический признак матери, а дочери — признак отца. Полученные результаты Морган объяснил тем, что ген «w» находит+ся в X-хромосоме, а У-хромосома генетически инертна или, по крайней мере, не содержит гена «w». Этот тип наследования получил название наследования, сцепленного с полом. Таким образом, ген «w» сцеплен с полом, т.е. находится в Х-хромосоме. Установленные Т.Морганом законы справедливы и для человека. Проиллюстрируем законы наследования признаков, сцепленных с полом, установленные Морганом, на следующем примере. В брак вступают женщина-дальтоник (рецессивный признак) и мужчина с нормальным цветовосприятием. Изучая цитологическую схему данного брака, видим, что сыновья единственную Х-хромосому получают от матери, следовательно, будут иметь ее фенотип по данному признаку (дальтоники). Дочери получают одну Х-хромосому от матери (несущую рецессивный аллель дальтонизма), а другую Х-хромосому от отца (несущую доминантный аллельцветовосприятия) и будут иметь нормальное зрение. Следовательно, фенотипический признак отца перешел к дочерям, а от матерей — к сыновьям (крисс-кросс наследование). В случае, если мать имеет нормальное зрение, а отец дальтоник, все дети будут иметь нормальное цветовосприятие, а если дочь с нормальным цветовосприятием, но носитель гена дальтонизма, выйдет замуж за здорового мужчину, то возможное соотношение фенотипов у детей будет 3 : 1. Сотрудник Т.Моргана К Бриджес обратил внимание на редкие отклонения от схемы крисс-кросс наследования. Бриджес предположил, что такие отклонения в наследовании могут быть связаны с нарушением расхождения хромосом в мейозе. Он исследовал хромосомы таких исключительных мух и убедился, что белоглазые самки имеют У-хромосому наряду с двумя XX, а красноглазые самцы — одну Х-хромосому. Тем самым было впервые доказано, что определенный ген w находится в конкретной хромосоме — X. В F, от скрещивания красноглазой самки и белоглазого самца изредка встречались мухи, у которых один глаз белый, а другой красный. При более внимательном рассмотрении оказывается, что эти мухи симметрично представлены женской и мужской половинками тела. Таких мух называют билатеральными гинандроморфами. При этом белый глаз находится на мужской половине. Эти особи возникают в результате потери одной Х-хромосомы при первом дроблении зиготы, которая должна дать начало самке. Потери хромосом могут происходить и на более поздних стадиях развития. Тогда появляются организмы-мозаики, у которых в разных пропорциях представлены участки тела, состоящие из клеток с неодинаковым числом хромосом. Бриджес в 1922 г. показал, что пол у дрозофилы определяется не числом Х-хромосом, а зависит от соотношения половых Х-хромосом и наборов аутосом (А). Такое соотношение было названо половым индексом. Если это соотношение равно : 2Х:2А=1, то из зиготы развивается самка, если оно равно 1 Х:2Азж0,5, то — самец. При этом У-хромосома в определении пола роли не играет. При промежуточном соотношении развиваются (2Х:ЗА=0,67) интерсексы, мухи, имеющие промежуточный фенотип. У части животных (пчел, муравьев и ос) существует особый тип определения пола, названный гапло-диплоидным. У этих особей нет половых хромосом. Самки развиваются из оплодотворенных яиц и диплоидны, а самцы — из неоплодотворенных яиц и гаплоидны. При сперматогенезе число хромосом не редуцируется. Человек является исключением из этого правила. Вопрос 36(механизмы генотипического определения ,и дифференциации признаков пола в развитии) У человека важнейшим фактором определения пола в эмбриональном развитии является хромосома Y. Независимо от числа хромосом X отсутствие хромосомы Y в кариотипе развивающегося организма не позволяет дифференцироваться мужскому полу. Относительно недавно было показано, что в дифференциации мужского пола особая роль принадлежит гену SRY (сокращение расшифровывается как «пол-определяющая область хромосомы Y»). Мутации в гене SRY приводят к тому, что у лиц с кариотипом XY развивается женский фенотип. При введении гена SRY в клетки эмбриона мыши женского пола происходит трансформация пола и у новорожденного мышонка пол оказывается мужским. Эмбриональные гонады могут дифференцироваться в яички и яичники. Если эмбриональные гонады находятся у плода мужского пола, то ген SRY, локализованный в Y-хромосоме, продуцирует фактор, детерминирующий дифференцировку эмбриональных яичек. Эмбриональные яички секретируюг гормоны, определяющие развитие мужских половых органов. В отсутствие гена SRY эмбриональные гонады дифференцируются в яичник. Установлено, что, кроме гена SRY, несколько аутосомных генов играют определенную роль в мужской дифференциации пола. Один из них — ген супрессор опухоли Вильмса, кодирует фактор транскрипции, связывающийся со специфическими ДНК-последовательностями. Точковые мутации в этом гене иногда приводят к формированию гонадно противоположного пола. Второй ген такого же типа - SOX9. Мутации в гене SOX9 вызывают дисплазию, при которой у индивидуумов 46,ХУ фенотипически развивается женский пол. Нет сомнений в том, что современные представления о генетическом контроле становления пола далеко не совершенны. Это относится не только к тому, какие гены вовлекаются в данный процесс, но и к тому, на каких стадиях развития и в каких тканях эти гены проявляются. Развитие признаков пола, как и любых других признаков организма, определяется генотипом и факторами внешней среды. Так как организмы генетически бисексуальны, процесс дифференциации пола оказывается сложным. Бисексуальная основа организма в принципе позволяет изменять направление его развития, т. е. переопределять пол в онтогенезе. Вслед за определением пола следует дифференциация, т. е. развитие половых различий: формирование воспроизводительной системы, а также физиологического и биохимического механизмов, обеспечивающих скрещивание. Зачаточные индифферентные в половом отношении гонады у эмбрионов животных имеют двойственную природу. Они состоят из внешнего слоя — кортекса (cortex), из которого в процессе дифференциации развиваются женские половые клетки, и из внутреннего слоя — медуллы (medulla), из которого развиваются мужские гаметы (рис. 122). В ходе дифференциации пола идет развитие одного из слоев гонады и подавление другого. У мужского пола быстрее развивается медуллярная ткань, которая подавляет деятельность кортикального слоя, в результате гонады превращаются в семенники. У женского пола ускоряется развитие кортикального слоя, в силу чего подавляется формирование медуллярного слоя и гонады превращаются в яичники. В соответствии с этими преобразованиями дифференцируются и половые пути, которые тоже закладываются одинаковыми у особей обоих полов. Процесс дифференциации пола у многих животных обусловлен гормонами, которые выделяются не только эндокринными железами, но кортикальным и медуллярным слоями полового зачатка, а в последующем — и половыми железами. Эти вещества отличаются по характеру своего действия и по времени их выработки. У особей мужского пола преобладает «мужское вещество» — медулларин, у женского — кортикальный гормон. Дальнейшая дифференциация пола, особенно развитие вторичных половых признаков, также идет под влиянием различных гормонов. Уровень гормональной секреции контролируется генами, точнее, их балансом. Преобладание генов, определяющих мужской пол, в общем балансе приводит к повышению активности мужских гормонов и к дифференциации мужского пола, обратное соотношение генов — к развитию женского пола. Смена активности гормональной секреции то одного, то другого пола в онтогенезе приводит к развитию интерсексуальных форм. На дифференциацию пола у высших растений значительное влияние оказывают растительные гормоны — ауксины. Вопрос 37(Множественные аллели,
Все рассмотренные до сих пор примеры относились к таким точкам хромосомы — локусам, которые могут быть заняты одним из аллелей — доминантным или рецессивным. Однако во многих, если не в большинстве локусов могут быть еще и такие аллели, которые отличаются по своему фенотипическому проявлению как от доминантного, так и от рецессивного генов. Три или большее число генов, которые могут находиться в одном и том же локусе, т. е. занимать одно и то же положение в гомологичных хромосомах, называют серией множественных аллелей. Все они обусловливают различные фенотипы. Каждая особь в популяции может иметь любые два из таких аллелей, но никак не больше, а каждая гамета, разумеется, содержит только один из них. Однако в популяции в целом соответствующий локус может быть представлен тремя или большим числом аллелей. По типу множественных аллелей наследуются группы крови О, А, В и АВ у человека. Ген IA содержит код для синтеза в эритроцитах специфического белка — агглютиногена А; ген IB вызывает синтез другого белка — агглютиногена В, ген i не продуцирует никакого агглютиногена. Ген i рецессивен по отношению к двум другим, но ни ген IA, ни ген IB не доминируют друг над другом. (Символы IA, IB и i указывают на то, что все эти три гена — аллели одного и того же локуса.) Таким образом, генотипы IA IA и IA i обусловливают группу крови А, генотипы IB IB и IB i — группу В, а генотип ii — группу О. Если же у человека имеются оба нерецессивных гена IA и IB, то у него образуются оба агглютиногена и он имеет группу крови АВ. Поскольку группы крови обусловлены генетически и не изменяются в течение всей жизни, их определение может помочь установить истину в случаях спорного отцовства. Однако на основании группы крови нельзя доказать, что данный человек действительно является отцом данного ребенка; можно лишь установить, что он мог бы быть его отцом, или же, исключить такую возможность. Подумайте: может ли мужчина с группой крови АВ быть отцом ребенка с группой О? Может ли ребенок с группой АВ быть сыном мужчины с группой О? Может ли ребенок с группой В иметь мать с группой А и отца с группой А или О? Наряду с системой АВО существует еще с десяток других систем групп крови, в том числе система MN и серия аллелей Rh; они определяются другими генами и наследуются независимо от групп АВО. Определение всех этих групп дозволяет иногда выяснять родственные отношения, которые нельзя было бы достоверно установить на основании одних лишь групп системы АВО. |