ЭКЗАМЕН по Биологии вопросы 29-42. Исторический очерк
 Скачать 0.62 Mb. Скачать 0.62 Mb.
|

Полигенное наследование признаковКумулятивная полимерия. Значительная часть признаков у эукариот, наследуемых по-лигенно, находится под контролем не двух-трех, а большего числа генов (их количество пока еще трудно определить). При моногенном типе наследования в моногибридном скрещивании один ген проявляется в двух альтернативных состояниях без переходных форм. Такие признаки относятся к качественным, при их анализе, как правило, не проводится никаких измерений. При неаллельном взаимодействии двух несцепленных генов даже при сохранении менделевского отношения 9:3:3:1 фенотип первого поколения гибридов зависит от действия обоих генов. Однако наследование качественных признаков может определяться взаимодействием трех и более генов. При этом каждый из этих генов имеет свою долю влияния на развитие признака. Примером может служить наследование красной и белой окраски зерен пшеницы в опытах шведского генетика Нильсона-Эле. Результаты этих опытов были опубликованы в 1909 г. При скрещивании сорта пшеницы, зерна которой имели темно-красную окраску, с сортом, имеющим белые зерна, гибриды первого поколения имели красную окраску более светлых тонов. Во втором поколении получилось такое соотношение по фенотипу: на 63 окрашенных зерна с различными оттенками красного цвета приходилась 1 белое зерно (неокрашенное). Эти результаты были объяснены Нильсоном-Эле следующим образом. Темно-красная окраска зерен пшеницы обусловлена действием трех пар доминантных генов, а белая - трех пар рецессивных, при этом по мере увеличения числа доминантных генов окраска становится более интенсивной. Обозначим доминантные аллели трех генов, локализованных в разных хромосомах, прописными буквами А1 А2 А3 а рецессивные - строчными а1 а1 а3, тогда генотипы исходных форм будут: А1А1 А2А2 А3А3 x а1я1 а2а2 a33a. Окраска зерен у гибридов первого поколения A1a1 A2a2 A3a3 при наличии трех доминантных аллелей будет промежуточной светло-красной. При скрещивании гибридов первого поколения A1a1 A2a2 A3a3 x A1a1 A2a2 A3a3 у каждого из гибридов образуется по 8 типов гамет, поэтому во втором поколении ожидается расщепление в 64-х долях (8 х 8). Среди 63/64 растений с окрашенными зернами интенсивность окраски усиливается по мере увеличения числа доминантных аллелей различных генов в генотипе. Видимо, каждый доминантный ген способствует увеличению количества синтезированного пигмента, и в этом смысле такой признак можно отнести к количественным. Тип аддитивного действия генов, каждый из которых оказывает свою, часто небольшую, долю влияния на признак, называется кумулятивной полимерией. Используя решетку Пеннета, можно подсчитать частоты доминантных генов среди генотипов второго поколения. Для этого в каждой из 64 клеток вместо генотипа записывается число присутствующих в нем доминантных аллелей. Определив частоты доминантных аллелей, можно убедиться, что генотипы с числом доминантных генов 6,5,4,3, 2, 1,0 встречаются 1,6,15,20,15,6,1 раз соответственно. Эти данные представлены в виде графика на рисунке. На горизонтальной оси указано число доминантных генов в генотипе, а на вертикальной - частоты их встречаемости. С увеличением числа генов, определяющих один признак, этот график приближается к идеальному нормальному распределению. 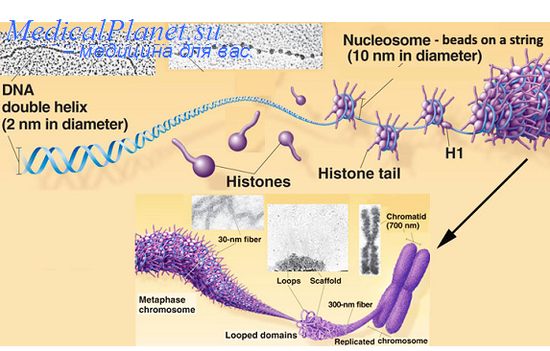 Такого типа графики характерны для количественных признаков, таких как рост, вес, длительность жизни, яйценоскость и других признаков, показатели которых можно измерить. К количественным относятся признаки, варьирующие более или менее непрерывно от одной особи к другой, что позволяет распределить особей по классам в соответствии со степенью выраженности признака. На рисунке приведен пример распределения по росту у мужчин. Эта выборка разделена на 7 классов с 5 см-интервалом. Мужчины со средним ростом (171-175 см) составляют большую часть выборки. С наименьшей частотой встречаются мужчины, которые включены в класс с ростом 156—160 см и 186—190 см. С увеличением выборки и с уменьшением классового интервала график может приблизиться к нормальному распределению по росту. Фенотипическая изменчивость без разрывов в проявлении, представленная на графике нормального распределения признака, называется непрерывной. Непрерывная изменчивость количественных признаков зависит от двух причин: 1) от генетического расщепления по большому числу генов, 2) от влияния среды, как причины модификационной изменчивости. Впервые датский генетик Иогансен показал, что непрерывная изменчивость такого количественного признака как масса бобов фасоли Phaseolus vulgaris зависит как от генетических, так и средовых факторов. Путем инбридинга в течение ряда поколений он вывел несколько чистых (гомозиготных) линий, различающихся по средней массе бобов. Например, средняя масса бобов в линии 1 была 642 мг, в линии 13 —454 мг, в линии 19 — 351 мг. Далее Иоган-сен вел отбор крупных и мелких бобов в каждой линии с 1902 по 1907 г. Вне зависимости от массы родительских семян средняя масса бобов после 6 лет отбора была такой же, как и в исходной линии. Так влинии № 13 при массе родительских семян от 275 мгдо 575 мг средняя масса семян в потомстве сохранилась на том же уровне ±450 мг. При этом в каждой линии масса бобов варьировала от минимальных до максимальных значений, а наиболее многочисленным был класс со средней массой, что характерно для количественных признаков. Отбор в чистых линиях оказался невозможен . Еще один пример, в 1977 г. Д.С. Билева, Л.Н. Зимина, А.А. Малиновский изучали влияние генотипа и среды на продолжительность жизни двух инбредных линий Drosophila melanogaster. Путем инбридинга и отбора были выведены две линии № 5 и № 3, четко различающиеся по длительности жизни. Продолжительность жизни определялась на трех вариантах корма: полноценном (дрожжи, манная крупа, сахар, агар-агар), обедненном (манная крупа, сахар, агар-агар) и сахарном (сахар, агар-агар). Обеднение состава корма приводило к уменьшению длительности жизни. Продолжительность жизни самок 5-й линии на сахарном корме (в днях) снизилась с 58+2,1 до 27,2±1,8, а самцов с 63,7±2,9 до 34,8±1,5, т.е. оказалась примерно в 2 раза меньше, чем на полноценном корме. Такая же закономерность была характерна и для самок и самцов 3-й линии. Длительность жизни самок этой линии снизилась с 50,7±],9 до 24,3±1,2, а самцов с 32,9+2,9 до 21,6±1,5 дня. При этом гистограмма, отражающая изменчивость по данному признаку на полноценном корме, близка к гистофамме представленной на рисунке, я, а на обедненном и сахарном наблюдается ассиметричное распределение со сдвигом средней величины в сторону уменьшения длительности жизни. Некумулятивная полимерия. Наряду с кумулятивной (аддитивной) полимерией известны случаи наследования по типу некумулятивной (неаддитивной) полимерии, когда характер проявления признака не меняется в зависимости от числа доминантных полимерных генов. Так у кур оперенность ног определяется доминантными аллелями двух генов A1 и А2: Р А1А1 А2А2 х а1а1a2a2 оперенная неоперенная оперенные F2 9 А1_А2_; 3 А1_ а2а2:; 3 a1a1 A2_; 1 а1а1 a2a2 оперенные (15) неоперенные (1) В F2 среди 15/16 гибридов с оперенными ногами есть такие, которые имеют четыре доминантных аллеля (А1А1 А2А2), три (А1А'1 А2а2), два (А1а1 А2а2) или всего один (А1а1 а2а2), характер оперенности ног в этих случаях один и тот же. Главные гены в системе полигенов. Среди генов, влияющих на количественный признак, может оказаться «сильный» или главный ген, и более «слабые» гены. Действие главного гена иногда настолько существеннее действия других генов, что признак, кодируемый им, наследуется по мекделевским законам. Изменчивость одного и того же признака может находиться под контролем как одного главного гена, так и полигенов. Например, карликовость у человека в случае ахондроплазии обусловлена специфическим главным геном, в то время как изменчивость по росту в нормальной популяции индивидов является примером полигенной изменчивости. Гены, действие которых заметно сильнее действия других генов на этот признак, можно изучать по отдельности от действия других генов. С другой стороны, один и тот же ген вследствие плейотропного действия, может оказывать сильное влияние на один признак и менее значительное на другой признак. К тому же к главным генам могут быть отнесены те, которые определяют признаки, наследуемые по законам Менделя, без их отношения к системе полигенов. Подразделение генов на главные и неглавные не всегда обосновано, хотя бесспорно, что их роль в определении признака может быть различна. Широко распространенные болезни человека, например, артериальная гипертензия, ишемическая болезнь сердца, бронхиальная астма, язвенная болезнь желудка, наследуются полигенно. При этом тяжесть заболевания зависит не только от совокупного действия множества генов, но и от провоцирующих средовых факторов Вопрос 38 (Наследования группы крови и резус-фактора) 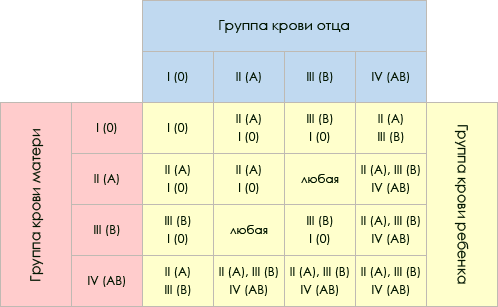 Группа крови — врожденное свойство человека и неизменна в течение всей его жизни (онтогенеза). К настоящему времени известно несколько систем группы крови. Каждая из этих систем наследственно обусловлена. Невозможно найти двух людей (кроме однояйцевых близнецов), которые имели бы одинаковые группы крови по всем системам. Это явление используется в судебной медицине. В клинической медицине для переливания крови необходимо знание группы крови системы АВО (I—IV группы крови) и резус-фактора. Система групп крови АВО открыта в начале XX века австралийским ученым К. Ландштейнером при изучении поведения эритроцитов (красных кровяных телец) в сыворотке (жидкой части) крови разных людей. Ученый обратил внимание на тот факт, что эритроциты в сыворотке крови одних людей распределяются равномерно, а других — склеиваются. Используя разные комбинации эритроцитов и сывороток, он обнаружил три группы крови (I—III), а существование IV группы (более редкой) было установлено позднее. Частота встречаемости групп крови системы АВ0 в разных популяциях человека различна. Обладание одной из четырех групп крови определяется парой генов, пришедших по одному от каждого из родителей. Каждый ген может быть в одной из трех аллелей (функциональных состояний) — А, В, О. Аллели А и В доминируют над О, но, оказавшись вместе в одном организме, А и В проявляют совместное действие (кодоминирование) и обусловливают наличие IV группы крови. Многие считают, что у родителей и детей группа крови всегда одна и та же. Это заблуждение. Установлено, что совпадение здесь имеет место далеко не во всех случаях. Фенотипическими (то есть биохимическими, морфологическими или другими методами) можно определить четыре группы крови: 1(О), II(А), III (В) и IV(АВ). Фенотипы I и IV групп совпадают с их генотипами. Генотипы же ВВ и ВО (для III группы крови), АА и АО (для II группы) без знания групп крови родителей различать невозможно. Рассмотрим такой пример. Двое мужчин имеют II группу крови, а их жены — I. В этом случае у одной супружеской пары могут родиться двое детей со II (или с I) группой крови, а у другой пары возможен один ребенок с I, а другой — со II группой крови. I группа крови новорожденному гарантирована только в том случае, если оба родителя имеют эту группу. Если же у обоих родителей II или III группа крови, то их дети, кроме родительской, могут иметь и I группу крови. В случае, когда у родителей II и III группы крови (например, у матери — II, у отца —III),их дети могут обладать I—IV группами. У родителей с I и IV группами крови (например, у матери — I, у отца — IV) дети будут иметь II или III группу. Правда, недавно в специальной литературе появилось описание нескольких японских семей, у ко¬торых родители имели I и IV группы крови, а дети — IV. Такие семьи представляют большой научный интерес, а «неправильное» наследование IV группы крови тщательно исследуется. !Присутствие генов А и В обусловливает наличие в эритроцитах антигенов А и В, а их отсутствие приводит к появлению антител А и В в сыворотке крови. Если антиген А, содержащийся в эритроцитах, встречается с антигеном А, содержащимся в сыворотке, то происходит склеивание эритроцитов. В норме такого не происходит, так как в крови нет антител, способных склеивать собственные эритроциты (антигены). Но около 5% зигот (оплодотворенных яйцеклеток) погибает, а в среднем 1 % новорожденных имеют гемолитическую болезнь из-за несовместимости матери и ребенка по группам крови системы АВ0. Если, например, ребенок имеет генотип 1А1°, а мать 1В1°, то у ребенка есть антиген А, а у матери — антитело А. Последние проникают в кровь плода и склеивают, а затем и разрушают эритроциты, обусловливая этим гемолитическую болезнь. Ученые предполагают, что существует специальная генетическая система защиты плаценты, которая нейтрализует антитела матери до их попадания в кровь плода. В 1940 г. К. Ландштейнер и А. С. Винер обнаружили в эритроцитах человека совершенно новый антиген, названный ими резус-фактором (Rh). Резус-фактор присутствует в крови 85% людей, а 15% лиц этого фактора не содержат. Система антигенов резус-фактора представлена 6 основными антигенами. Образование резус-антигенов контролируется тремя парами ал-лельных генов: Dd, Cc, Ее, которые расположены на двух хромосомах. Каждая из хромосом способна нести только 3 гена из 6, причем лишь 1 ген из каждой пары - D или d, С или с, Е или е. Гены Dnd, Сие, Еие являются по отношению к друг другу аллельными. В последнее время было доказано, что аллельного гена d не существует. Наиболее активным из всех антигенов является Rh0(D). В зависимости от его наличия или отсутствия кровь людей делят на резус-положительную (Rh+) или резус-отрицательную (Rh-). Указанные 6 антигенов резус встречаются в эритроцитах в виде одного из 18 возможных сочетаний. Фенотипически каждый человек содержит 5, 4 или 3 антигена резус в зависимости от количества генов, по которым он гомозиготен. Однако генотипическая формула изображается шестью буквами, например cDE/CDe, обозначающими 3 гена резус, унаследованных с хромосомой одного из родителей, 3 - с хромосомой другого. Иначе подходят к оценке резус-принадлежности лиц, являющихся донорами. Требуется дополнительное исследование крови доноров по факторам rh и rh. Резус-отрицательными могут быть только доноры, в крови которых отсутствуют все три антигена (Rh0, rh, rh). Такой подход к оценке резус-принадлежности доноров позволяет исключить возможность сенсибилизации реципиента к любому из трех основных антигенов: Rh"(D), rh(C), rh(E). В повседневной практике переливания крови ограничиваются опреАнтигены резус относят к липопротеидам. Они являются очень активными и способны вызывать образование иммунных антител. Резус-антитела, являясь иммунными (неполными, моновалентными, блокирующими), характеризуются способностью фиксироваться к резус-положительным эритроцитам, не вызывая их склеивания. Они агглютинируют эритроциты только в присутствии коллоидных растворов, протеолитических ферментов или под действием специально приготовленной антиглобулиновой преципитирующей сыворотки. Неполные антитела относят к классу иммуноглобулинов IgG. Наличие резус-антигена выявляется у человеческого плода начиная с 5-8 недели и хорошо выражено у 3-4-месячного эмбриона. Существование антигенов системы резус в эритроцитах человека является физиологическим, антител к этим антигенам в организме нет. Образование иммунных антител происходит при поступлении в организм человека чужеродного ему изоантигена. У сенсибилизированных людей антитела анти-Rh содержатся не только в крови, но и в экссудате, транссудате, моче, слезе и других средах. Вопрос 39( Взаимодействие неаллельных генов. Комплементарность, эпистаз ,полимерия) Многие признаки формируются при участии нескольких неаллельных генов, взаимодействие между которыми отражается на формировании фенотипа. Описаны три основных типа взаимодействия неаллельных генов: комплементарность, эпистаз (для качественных признаков) и полимерия (для количественных признаков). Комплементарность. Впервые взаимодействие неаллельных генов было обнаружено в начале 20-го столетия при анализе наследования формы гребня у кур. Разные породы кур (леггорны, виандоты, европейские, малайские) имеют соответственно листовидный, розовидный, гороховидный и ореховидный гребни. В результате скрещивания кур, имеющих розовидный и гороховидный гребни, в потомстве первого поколения (Fj) возникает новая ореховидная форма гребня (новая форма гребня, возникает из-за взаимодействия генов А и В). Скрещивание гибридов Fi приводит к следующим результатам во втором поколении (Fi). Впоследствии такой тип взаимодействия неаллельных генов, при котором совместно присутствующие в генотипе организма неаллельные гены обусловливают развитие нового признака, был назван компле-ментарностью. К комплементарным относятся такие гены, которые при совместном действии в генотипе в гомо- (ААВВ) или гетерозиготном (А-В-) состоянии обусловливают развитие нового признака. Присутствие доминантных аллелей двух генов А и В у 9/16 кур второго поколения ведёт к образованию нового фенотипа - ореховидного гребня. Действие же каждого гена в отдельности (А-вв или ааВ-) ведёт к воспроизведению признака лишь одного из скрещиваемых родителей. У человека по типу комплементарности взаимодействуют доминантные неаллельные гены М и R. Образование чёрного пигмента меланина контролирует ген М, который представлен в популяциях людей тремя аллелями: М8, MBw, MBd. Доминантный аллель (R2) второго гена контролирует синтез красного пигмента. Сочетание аллелей названных генов обусловливает весь спектр пигментации волос у человека. Исключение составляют альбиносы, гомозиготные по гену альбинизма, который локализован в ином локусе и вызывает полное отсутствие синтеза пигмента в организме. Противоположным комплементарности типом взаимодействия неал-лельных генов является, по своей сути, эпистаз. Под эпистазом понимают подавление одним геном действия другого неаллельного гена. Различают рецессивный эпистаз (аа>ВВ) - когда эпистатируют рецессивные аллели, и доминантный эпистаз (АА>ВВ, или АА>вв). Явление эпистаза открыто при анализе наследования масти лошадей (рис. 65). Известно, что вороная окраска определяется доминантным аллелем В, рыжая - рецессивным аллелем Ъ, доминантный аллель С обусловливает из-за раннего поседения волоса серую масть. Аллель С контролирует нормальное развитие и пигментацию волос. Гомозиготы и гетерозиготы по аллелю С всегда будут серыми из-за седины, независимо от того, какие аллели гена В будет содержать генотип лошади: Это расщепление выводится из расщепления 9:3:3:1, поскольку 3/16 потомков, имеющих генотип ЪЪС-, по фенотипу будут также серые и в сумме с 9/16 дадут 12/16. Подавляющий ген называется эпистатическим (эпистазирующим) или геном-супрессором, а подавляемый ген - гипо-статическим геном. Эпистаз наиболее характерен для генов, участвующих в регуляции работы структурных генов в онтогенезе и контролирующих процессы в иммунной системе человека. Известно немало примеров эпистатического взаимодействия локусов у человека, приводящих к тому, что тот или иной доминантный аллель у некоторых индивидуумов не получает фенотипического выражения. По- добным примером может послужить полидактилия, контролируемая, как правило, доминантным аллелем. Иногда она встречается у детей «совершенно здоровых» родителей (у последних действие данного аллеля, вероятнее всего, подавлялось другими генами). Эпистаз и комплементарность характеризуют наследование альтернативных признаков, т.е. различающихся качественно. В 1908 году шведский генетик Г. Никольсон-Эле, скрещивая пшеницу с красными и белыми зернами, обнаружил в ряде случаев в потомстве F2 расщепление 15:1, т.е. 15 зёрен оказывались красными и одно неокрашенным. Последующий анализ в F3 показал, что дальнейшее расщепление отсутствовало только у растений с наиболее красной и с чисто белой окраской. В результате анализа же промежуточных форм выявилось, что красная окраска контролируется двумя доминантными неаллельными генами. Интенсивность окраски определяется числом доминантных аллелей, присутствующих в генотипе. Неаллелъные гены такого типа были названы полимерными. Поскольку эти гены влияют на один и тот же признак, было принято обозначать их одной латинской буквой с указанием индекса А}, А2, А3 и т.д. Следовательно, исходные родительские формы, давшие в опытах Г. Нильсона-Эле расщепление 15:1, имели генотипы А1А1А2А2 и а/Я/а^. Гибрид Fj обладал генотипом AjaiA2a2, а в потомстве F2 развились зерна с разным числом доминантных генов. Наличие всех четырёх доминантных аллелей генов окраски А1А1А2А2 у 1/16 растений определяли самую интенсивную окраску зерна, 4/36 всех зёрен F2 имели три доминантных аллеля (AiA^ai), 6/16 - два доминантных аллеля (A\aiA2a2), 4/16 - один аллель (Aja/a2a2). Эти генотипы определяли промежуточные типы окраски, переходные между интенсивно красной и белой. Гомозиготная по обоим рецессивным генам (aia^ai) 1/16 всех зёрен оказалась неокрашенной. По типу полимерных генов наследуется пигментация кожи у человека. Например, в потомстве представителей европеоидной и австрало-негроидной рас могут родиться мулаты с промежуточной интенсивностью пигментации кожи: Полимерия бывает двух типов: а) кумулятивная полимерия, когда число доминантных аллелей в генотипе организма влияет на степень выраженности данного признака (проиллюстрирована выше); б) некумулятивная полимерия, при которой не имеет значения количество доминантных аллелей в генотипе, а важно только их присутствие. Комплемента́рность (в химии, молекулярной биологии и генетике) — взаимное соответствие молекул биополимеров или их фрагментов, обеспечивающее образование связей между пространственно взаимодополняющими (комплементарными) фрагментами молекул или их структурных фрагментов вследствие супрамолекулярных взаимодействий (образование водородных связей, гидрофобных взаимодействий, электростатических взаимодействий заряженных функциональных групп и т. п.). Взаимодействие комплементарных фрагментов или биополимеров не сопровождается образованием ковалентной химической связи между комплементарными фрагментами, однако из-за пространственного взаимного соответствия комплементарных фрагментов приводит к образованию множества относительно слабых связей (водородных и ван-дер-ваальса) с достаточно большой суммарной энергией, что приводит к образованию устойчивых молекулярных комплексов. Вместе с тем, следует отметить, что механизм каталитичекой активности ферментов определяется комплементарностью фермента и переходного состояния либо промежуточного продукта катализируемой реакции — и в этом случае может происходить обратимое образование химической связи. Эпистаз |