ЭКЗАМЕН по Биологии вопросы 29-42. Исторический очерк
 Скачать 0.62 Mb. Скачать 0.62 Mb.
|

|
Ген — структурная и функциональная единица наследственности, контролирующая развитие определённого признака или свойства. Совокупность генов родители передают потомкам во время размножения. Однако перенос генов от родителей к потомкам не является единственным способом передачи генов. В 1959 году был описан случай горизонтального переноса генов. В отличие от вертикального переноса, в горизонтальном организм передаёт гены организму, который не является его потомком. Этот способ передачи широко распространён среди одноклеточных организмов и в меньшей степени среди многоклеточных. Гены эукариот Отметим вначале, что у эукариотических организмов ДНК присутствует не только в ядрах, но и в органеллах — митохондриях, которые есть у всех эукариот, и хлоропластах, имеющихся у зеленых растений. По многим признакам предполагается, что орга-неллы происходят от прокариот: митохондрии от а-пурпурных бактерий, а хлоропласты — от цианобактерий. Их роднят с прокариотами многие черты белок-синтезирующего аппарата. Учитывая направленность интересов генетической инженерии, ограничимся здесь рассмотрением только ядерных генов. Строение. Гены эукариот по строению и характеру транскрипции значительно отличаются от прокариотических генов. Их отличительной особенностью является прерывность, т. е. чередование в них последовательностей нуклеотидов, которые представлены (экзоны) или не представлены (интроны) в мРНК. Отсюда ясно, что интроны относятся к некодирующим последовательностям. Они могут располагаться не только в области, ограниченной инициирующим и терминирующим кодонами, но и вне их, в начале или в конце гена. Их длина может превышать 10 т.п.н. У низших эукариот прерывные гены составляют меньшинство всех генов (5 % у дрожжей), а у высших — большинство (94 % у млекопитающих). Отметим, что мозаичность генов найдена и в прокариотических клетках. Эволюционно связанные гены, обладающие высокой степенью физической гомологии, образуют семейства. Белки, кодируемые такими генами, действуя одновременно или на разных этапах развития организма, выполняют одинаковые функции. Например, состав белков в а- и р-цепях гемоглобина крови млекопитающих различен у эмбриона, плода и взрослого организма, что вызвано дифференциальной экспрессией генов, входящих в а- и р-семей-ства глобиновых генов. Наряду с функционирующими генами, в семействах обнаружены нефункционирующие. Такие гены получили название псевдогенов. Они не экспрессируются по различным причинам (изменение рамки считывания из-за делеции или вставки, отсутствие интрона и т. п.). Характерной чертой генов, входящих в семейство, является сходная картина локализации большинства интронов. Это сходство не ограничивается рамками определенного генома. Так, в случае глобиновых генов сходными по расположению интронов оказались гены у всех исследованных животных — у млекопитающих, птиц и лягушек. Однако длины и нуклеотидные последовательности интронов могут значительно варьировать, меняя тем самым и размеры самих генов. Транскрипция. Гены эукариот не группируются в опероны, поэтому каждый из них имеет собственные промотор и терминатор транскрипции. Транскрипцию ведут три различные РНК-полимеразы: I, II и III, которые синтезируют рРНК, мРНК и тРНК, соответственно. Как и в случае прокариот, рассмотрим только механизм экспрессии генов, кодирующих белки. Поэтому далее под эукариотической РНК-полимеразой подразумевается РНК-полимераза II. Она состоит из более десятка субъединиц, но все же связываться непосредственно с промотором не может. Ее посадке на промотор способствуют транскрипционные факторы белковой природы. Ряд из них распознают специфические последовательности (боксы) в промоторе. Длина типового промотора высших эукариот — около 100 п.н. В нем следует различать две части — базовую и дополнительную. Гены, имеющие только базовую часть промотора, функционируют в любых клетках организма и не подвержены ткане-специфичес-кому контролю. Эта часть служит для инициации транскрипции и точной ориентации РНК-полимеразы II относительно первого транскрибируемого нуклеотида. Дополнительная часть совместно с энхансерами используется для повышения эффективности транскрипции и регуляции активности гена. Прокариоты (лат. Procaryota, от др.-греч. προ «перед» и κάρυον «ядро»), или доядерные — одноклеточные живые организмы, не обладающие (в отличие от эукариот) оформленным клеточным ядром и другими внутренними мембранными органоидами (за исключением плоских цистерн у фотосинтезирующих видов, например, у цианобактерий). Единственная крупная кольцевая (у некоторых видов — линейная) двухцепочечная молекула ДНК, в которой содержится основная часть генетического материала клетки (так называемый нуклеоид) не образует комплекса с белками-гистонами (так называемого хроматина). К прокариотам относятся бактерии, в том числе цианобактерии (сине-зелёные водоросли), и археи. Потомками прокариотических клеток являютсяорганеллы эукариотических клеток — митохондрии и пластиды. Прокариоты разделяют на два таксона в ранге домена (надцарства): Бактерии (Bacteria) и Археи (Archaea).[1] Для клеток прокариот характерно отсутствие ядерной оболочки, ДНК упакована без участия гистонов. Тип питания осмотрофный. Генетический материал прокариот представлен одной молекулой ДНК, замкнутой в кольцо, имеется только один репликон. В клетках отсутствуют органоиды, имеющие мембранное строение. В геноме могут присутствовать мобильные генетические элементы, а у некоторых прокариот (например, вольбахия) их содержится необычно много. Изучение бактерий привело к открытиюгоризонтального переноса генов, который был описан в Японии в 1959 г. Это процесс широко распространен среди прокариот, а также у некоторых эукариот. Открытие горизонтального переноса генов у прокариот заставило по-другому взглянуть на эволюцию жизни. Ранее эволюционная теория базировалась на том, что виды не могут обмениваться наследственной информацией. Прокариоты могут обмениваться генами между собой непосредственно (конъюгация, трансформация) а также с помощью вирусов -бактериофагов (трансдукция). Уникальные гены — это гены, которые встречаются в клетке два или несколько раз (до 10-20). Большинство исследователей считает, что у многоклеточных общее число генов в среднем равно сто тысяч и подавляющее их число — это уникальные гены. Характерная черта генов эукариотов — мозаичное экзон-интронное строение. Интроны, не несущие генетической информации, вырезаются (сплайсинг). Число и размер интронов у разных видов варьируется. Присутствие их в гене приводит к значительному увеличению размеров гена. Интроны стабилизируют экзоны, однако существует представление, что интрон — это так называемая «эгоистическая» ДНК, не дающая организму никаких эволюционных преимуществ. Экзоны контролируют синтез белков: 1 экзон — 1 домен. К повторяющимся генам относятся прежде всего гены больших и малых рРНК и гистонов. Число их сильно варьирует и может достигать более 2000. Гены больших рРНК организованы в блоки, в которых последовательно идут гены 18S рРНК, 58S рРНК и 28S рРНК. Между ними имеются промежутки, различающиеся по длине у разных организмов. Межгенные участки имеют повторы разных типов, с необычной последовательностью, богатых парами ГЦ. Гены низкомолекулярных ядерных РНК блоков не образуют. Гены гистонов повторяются в геноме десятки (у млекопитающих), и сотни (у дрозофилы), и тысячи (у аксолотля) раз. Причем не удается уловить связи между этим показателем и положением организма на эволюционной лестнице. Перестраивающиеся, или рекомбинирующие, гены — это гены, кодирующие легкие и тяжелые цепи белков иммуноглобулинов, выполняющих функции антител. Гены этих белков состоят из двух типов генов для легких и пяти типов — для тяжелых цепей. Легкие цепи кодируются тремя отдельными генетическими элементами, тяжелые — четырьмя. Перестройки генома приводят к соединению разных участков и в итоге — к образованию иммуноглобулинов разных классов. Прыгающие гены, или транспозоны, — мобильные генетические элементы. Являясь нормальным компонентом генома, они составляют его значительную часть (у дрозофилы- 7% генома), могут быть представлены многими копиями, рассеянными по геному, и имеют варьирующую локализацию. Структура разных классов мигрирующих элементов (МЭ) варьирует, но для всех их характерно наличие на концах обращенных повторов. В середине МЭ могут иметь уникальные последовательности. МЭ проявляют высокую локусную специфичность, так как могут встраиваться в определенную последовательность на хромосоме. Повторы на днк Повторяющаяся ДНК - атрибут любой генетической системы. Еще на самых ранних этапах эволюции, когда возник почти полный репертуар полипептидов - носителей основных молекулярных функций (что-то вроде словаря генетического языка), им уже, по всей видимости была присуща внутренняя периодичность ( Ohno, 1981 , 1984 ; Go, 1983 ). Наличие олигомерных повторов в современных генах и белках объясняют тем, что вообще в любой информационной системе копирование текста - эффективное средство повышения помехоустойчивости при передаче сообщений . Применительно к генетическому языку существование внутригенных повторов резко снижает негативные эффекты таких ошибок, как сдвиги рамки трансляции, различные делеции и вставки ( Ohno, 1984 ). Действительно, белки, чьи гены содержат внутренние олигомерные повторы, также должны обладать периодичной первичной структурой. При этом длина полипептидного повтора будет зависеть от того, кратна ли трем длина соответствующего матричного повтора. Например, повторы из шести нуклеотидов будут порождать исключительно дипептидную периодичность, тогда как пентамерным повторам в гене должна отвечать периодичность той же размерности и в полипептидном продукте ( рис 1 ). Но, пожалуй, самая интересная особенность таких повторов состоит в том, что они обеспечивают совпадение обеих периодичностей, нуклеотидной и аминокислотной, во всех трех возможных рамках трансляции ( Ohno, 1984 , 1987 ). Поэтому С.Оно и полагает, что еще в самом начале, на заре жизни, когда биохимическая машина трансляции работала с частыми сбоями и когда рамка считывания вряд ли была однозначно фиксирована, как раз такие повторы (длиной , не кратной трем) должны были иметь ощутимое селективное преимущество. В изрядном количестве олигонуклеотидные мотивы именно такой конфигурации присутствуют практически во всех проанализированных генах самых разных видов про- и эукариот. В принципе нельзя исключить, что по крайней мере некоторые из них сохранились с тех давних времен, т.е. являются своеобразными молекулярными реликтами. Далее, спонтанно возникнув, даже сравнительно короткие повторенные участки существенно увеличивают вероятность дупликации (и автоматически - мультипликации) как их самих, так и тех сегментов генома, которые ими фланкированы ( Smith, 1976 ). Конкретные механизмы внутригеномного размножения повторов могут быть разными (неравный кроссинговер, скользящая гиперрепликация, обратная транскрипция и т.д.), но само размножение представляет собой типичный автокаталитический процесс ( Orgel, Crick, 1980 ; Doolitle et al., 1984 ; и др.). И хотя в целом ряде случаев количество повторов (например, МГЭ) регулируется по принципу обратной связи ( O'hare, Rubin, 1983 ; Simons, Kleckner, 1983 ; Snyder, Doolitle, 1988), хотя для любого вида организмов в норме имеются, по-видимому, и неспецифические селективные барьеры на пути "эгоистичного" распространения повторов по геному ( Бердников, Родин, Жарких, 1982 ; Rodin et al., 1985 ; Родин, 1985а,б ), для большинства современных организмов, в особенности эукариотических, характерна чрезвычайно высокая концентрация повторов практически на всех уровнях молекулярно-генетической организации. Структурные гены — уникальные компоненты генома, представляющие единственную последовательность, кодирующую определённый белок или некоторые виды РН…….. Вопрос 42 (Классификация генов: гены структурные ,регуляторы В зависимости от локализации генов в структурах клетки различают ядерные и митохондриальные гены (рис. IV. 14). По своему функциональному назначению гены могут быть разделены на две группы. Первая группа представлена генами, кодирующими белки; вторая группа — генами, контролирующими синтез РНК. Среди генов, кодирующих белки, различают: — гены «домашнего хозяйства», продукты которых необходимы для обеспечения функции любого типа клеток; — гены терминальной дифференцировки, т. е. гены, обеспечивающие специализированные функции клеток; 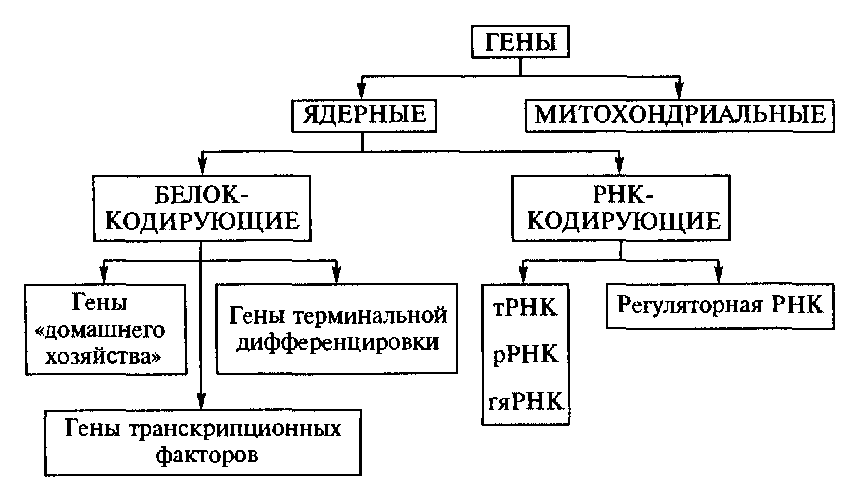 Рис. IV. 14. Классификация генов — гены траскрипционных факторов, контролирующие особые ядерные белки, способные соединяться с регуляторными областями многих структурных генов, вызывая либо активацию, либо подавление транскрипции. РНК-кодирующие гены определяют синтез различных видов РНК, необходимых для синтеза рибосом, обеспечения процессов трансляции, сплайсинга, а также для синтеза молекул РНК, влияющих на функционирование других генов (регуляторное действие). Гены человека, как правило, представляют собой функционально прерывистую последовательность нуклеотидов (рис. IV. 15). Относительно короткие кодирующие последовательности оснований чередуются в них с длинными некодирующими последовательностями. Последовательности гена, представленные в молекуле зрелой иРНК, получили название экзонов. Именно экзоны являются кодирующими участками гена, контролирующими аминокислотную последовательность белков. Экзоны разделены некодирующими участками — нитронами, которые вырезаются (сплайсинг) в процессе созревания иРНК и не участвуют в процессе трансляции. В настоящее время в понятие «ген» включаются не только транскрибируемые области (экзоны и интроны), но и фланкирующие ген последовательности. Фланкирующие области гена, как правило, высоко консервативны, т. е. характеризуются постоянством нук-леотидной последовательности, наблюдаемым даже при сравнении представителей различных видов. Фланкирующие области гена содержат последовательности, необходимые для его правильной работы: например, промоторная область в начале 5'-области или хвостовая нетранслируемая область поли-А, расположенная на З'-конце гена. Так, ТАГА — бокс (последовательность чередования Гены структурные регуляторы…. Генетическая информация о структуре белков и нуклеиновых кислот у всех организмов заключена в молекулах ДНК или РНК в виде последовательностей нуклеотидов, называемыхгенами . Совокупность генов организма, наряду с другими последовательностями ДНК составляет геном . Координированная работа (экспрессия) большого числа генов возможна благодаря наличию регуляторных механизмов, определяющих место, время и уровень экспрессии конкретного гена или группы генов. Чтобы экспрессия гена была регулируемой, он должен содержать индивидуальную (регуляторную) метку, по которой регуляторные компоненты генетической системы клетки или организма могли бы безошибочно оказать на него необходимое воздействие. В соответствии с этим любой ген состоит из двух основных функциональных частей (последовательностей нуклеотидов) - регуляторной и структурной. Регуляторная часть обеспечивает первые этапы реализации генетической информации, заключенной в структурной части гена. Размер гена складывается из размеров его структурной и регуляторной частей. Определить протяженность гена не так просто, особенно в случае генов эукариот. Отдельные элементы регуляторной области генов, например, энхансеры , могут располагаться на значительном (>60 т.п.о.) расстоянии от структурной части гена как перед ней, так и позади нее или даже в ней самой. В структурной части большинства эукариотических генов кодирующие последовательности нуклеотидов ( экзоны ) перемежаются протяженными некодирующими последовательностями (интронами ). Суммарный размер интронов, как правило, многократно превышает суммарный размер экзонов конкретных генов. Таким образом, геном эукариотического организма содержит не только последовательность нуклеотидов с генетической информацией о белках и нуклеиновых кислотах, но и большое количество последовательностей нуклеотидов, не несущих такой информации. Помимо интронов в геноме эукариот имеется большое количество других некодирующих последовательностей нуклеотидов, поэтому общая длина некодирующих последовательностей нуклеотидов в геноме эукариот в десятки раз превышает длину кодирующих последовательностей. Не вполне определенные и очень большие размеры генов эукариот, к тому же расположенных в геноме среди многочисленных некодирующих последовательностей нуклеотидов, создают значительные трудности для изучения их структуры и функционирования in vivo Свойство генов Дискретность (от лат. discretus — разделённый, прерывистый) — свойство, противопоставляемое непрерывности, прерывность. Под дискретностью понимают:
Стабильность — способность системы функционировать, не изменяя собственную структуру, и находиться в равновесии. Это определение должно быть неизменным во времени. Лабильность (от лат. labilis — скользящий, неустойчивый) в физиологии — функциональная подвижность, скорость протекания элементарных циклов возбуждения в нервной и мышечной тканях. Понятие «лабильность» введено русским физиологом Н. Е. Введенским (1886), который считал мерой лабильности наибольшую частоту раздражения ткани, воспроизводимую ею без преобразования ритма. Лабильность отражает время, в течение которого ткань восстанавливает работоспособность после очередного цикла возбуждения. Наибольшей лабильностью отличаются отростки нервных клеток — аксоны, способные воспроизводить до 500—1000 импульсов в 1 с; менее лабильны центральные и периферические места контакта — синапсы (например, двигательное нервное окончание может передать на скелетную мышцу не более 100—150 возбуждений в 1 с). Угнетение жизнедеятельности тканей и клеток (например, холодом, наркотиками) уменьшает лабильность, так как при этом замедляются процессы восстановления и удлиняется рефрактерный период. Лабильность — величина непостоянная. Так, в сердце под влиянием частых раздражений рефракторный период укорачивается, а следовательно, возрастает лабильность. Это явление лежит в основе т. н. усвоения ритма. Учение о лабильности важно для понимания механизмов нервной деятельности, работы нервных центров и анализаторов как в норме, так и при различных болезненных отклонениях. Множественный аллелизм — это существование в популяции более двух аллелей данного гена. В популяции оказываются не два аллельных гена, а несколько. Возникают в результате разных мутаций одного локуса. Гены множественных аллелей взаимодействуют между собой различным образом. В популяциях как гаплоидных, так и диплоидных организмов обычно существует множество аллелей, для каждого гена. Это следует из сложной структуры гена — замена любого из нуклеотидов или иные мутации приводят к появлению новых аллелей. Видимо, лишь в очень редких случаях любая мутация столь сильно влияет на работу гена, а ген оказывается столь важным, что все его мутации приводят к гибели носителей. Так, для хорошо изученных у человека глобиновых генов известно несколько сотен аллелей, лишь около десятка из них приводит к серьёзным патологиям. Плейотропи́я (от греч. πλείων — «больше» и греч. τρέπειν — «поворачивать, превращать») — явление множественного действия гена. Выражается в способности одного гена влиять на несколько фенотипических признаков. Таким образом, новая мутация в гене может оказать влияние на некоторые или все связанные с этим геном признаки. Этот эффект может вызвать проблемы при селективном отборе, когда при отборе по одному из признаков лидирует один из аллелей гена, а при отборе по другим признакам — другой аллель этого же гена. Плейотропия — это действие одного гена на несколько фенотипических признаков. Продукт фактически каждого гена участвует как правило в нескольких, а иногда и в очень многих процессах, образующих метаболическую сеть организма. Особенно характерна плейотропия для генов, кодирующих сигнальные белки….. специфичность — каждый ген кодирует свой признак |