ЭКЗАМЕН по Биологии вопросы 29-42. Исторический очерк
 Скачать 0.62 Mb. Скачать 0.62 Mb.
|

|
Моногибридное скрещивание. Некоторые закономерности наследования были впервые установлены Г. Менделем. Он достиг успеха в своих экспериментах благодаря использованию гибридологического метода —скрещивания организмов, различающихся по каким-либо признакам, и анализа всех последующих поколений с целью установления закономерностей наследования этих признаков. Гибридологический метод и до настоящего времени остается одним из основных в генетических исследованиях. Г. Мендель усовершенствовал данный метод, и в отличие от своих предшественников, анализировал наследование ограниченного количества признаков (одного, двух, трех). При этом он выбирал признак с альтернативным(контрастирующим) проявлением его у скрещиваемых организмов. Так, он скрещивал сорта гороха с окрашенными и белыми цветками, гладкими и морщинистыми семенами и т. п. Кроме того, Мендель проверял перед скрещиванием, насколько устойчиво наследуются выбранные им признаки в ряду поколений при самоопылении. В процессе эксперимента им проводился также точный количественный учет всех гибридных растений во всех поколениях. Моногибридное скрещивание. I и II законы Г. Менделя. Моногибридным называется такое скрещивание, при котором родительские пары различаются по одному признаку. В своих опытах Мендель использовал горох: отцовское растение с красными цветками, а материнское — с белыми или наоборот. Родительские организмы, взятые для скрещивания, обозначают латинской буквой Р (рис. 1 и 2). 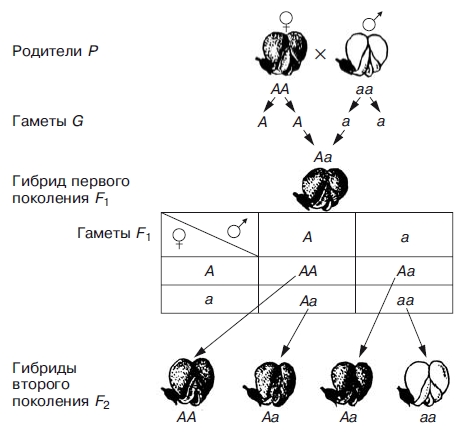 Рис. 1.Схема моногибридного скрещивания. Наследование пурпурной и белой окраски цветков у гороха: À — фактор пурпурной; à — фактор белой окраски цветка  Рис. 2. Схема, иллюстрирующая поведение пары гомологичных хромосом при моногибридном скрещивании: À — фактор пурпурной окраски цветка; à — фактор белой окраски Полученные в результате скрещивания гибриды первого поколения F1 обладали только красными цветками. Следовательно, признак второго родителя (белые цветы) не проявился. Преобладание у гибридов первого поколения признака одного из родителей (красные цветки) Мендель назвал доминированием, а сам этот признак —доминантным («преобладающим»). «Подавляемый» признак (белые цветки) получил название рецессивного. Феномен преобладания одного из признаков у всех гибридов первого поколения Мендель определил как закон единообразия гибридов первого поколения (I закон Менделя). Он формулируется следующим образом: при скрещивании гомозиготных особей, анализируемых по одной паре альтернативных признаков, наблюдается единообразие гибридов первого поколения как по фенотипу, так и по генотипу. При скрещивании однородных гибридов первого поколения между собой во втором поколении F2Мендель наблюдал появление растений как с доминантными (красные цветки), так и с рецессивными (белые цветки) признаками. Эта закономерность носит название расщепления. И оно оказывалось не случайным, а строго закономерным: 3/4 от общего числа гибридов второго поколения F2имеют красные цветки, а 1/4 - белые. Иными словами, соотношение числа растений с доминантными и рецессивными признаками составляет 3 : 1. Из этого следует, что рецессивный признак у гибридов F 1 не исчез, а был подавлен и проявился во втором поколении. Расщепление во втором поколении гибридов было названо Менделем законом расщепления гибридов второго поколения (II закон Менделя). Формулируется следующим образом: при скрещивании гетерозиготных особей, анализируемых по одной паре альтернативных признаков, наблюдается расщепление в соотношении 3:1 по фенотипу и 1:2:1 по генотипу. Пытаясь дать объяснение выявленным закономерностям, автор теории высказал ряд предположений о механизмах наследования признаков: > поскольку у гибридов F1проявляется лишь один признак (доминантный), а второй (рецессивный) отсутствует, но вновь проявляется у гибридов F2, то, следовательно, наследуются не сами признаки, а наследственные факторы (какие-то материальные частицы), их определяющие; > эти факторы являются постоянными, присутствуют в организме попарно и передаются из поколения в поколение через гаметы, причем в половую клетку попадает лишь один наследственный фактор из пары; > при слиянии половых клеток в новом организме вновь оказывается пара наследственных факторов (по одному от отцовского и материнского организмов); > наследственные факторы неравноценны по своей «силе», более «сильный» доминантный подавляет более «слабый» рецессивный (чем и объясняется единообразие гибридов первого поколения F1); в ходе оплодотворения могут сливаться гаметы, несущие либо одинаковые факторы (только доминантные или только рецессивные), либо разные (одна гамета содержит доминантный, другая - рецессивный). В первом случае у нового организма будет присутствовать пара одинаковых факторов. Мендель назвал такие организмыгомозиготными (либо АА, либо аа). Во втором случае организмы содержат два разных фактора — онигетерозиготные (Аа); > сочетание доминантных и рецессивных факторов в строго определенных комбинациях обусловливает расщепление признаков в соотношении 3 : 1 у гибридов второго поколения F2. Теперь вместо слова «фактор» используется «ген». Все предположения, высказанные Менделем о механизме наследования признаков у организмов, получили в ходе развития науки полное подтверждение. Менделем была предложена и система записи результатов скрещивания с использованием буквенной символики, которой пользуются в генетике до сих пор. Парные наследственные факторы (т. е. аллельные гены) обозначаются одной буквой, при этом доминантный ген — прописной (например, А), а рецессивный — строчной (а). Для установления генотипа особи с доминантным признаком при полном доминировании применяютанализирующее скрещивание. Для этого данный организм скрещивают с рецессивным гомозиготным по данной аллели. Возможны два варианта результатов скрещивания:  Если в результате скрещивания получаетсяединообразие гибридов первого поколения, то анализируемая особь является гомозиготной, а если в F1, произойдет расщепление признаков 1:1, то — гетерозиготной. Рассмотрим результаты опытов по моногибридному скрещиванию в виде схемы на основе системы записи, предложенной Менделем (см. рис. 1). Уже при жизни ученого в работах исследователей указывалось на то, что закономерности наследования признаков при моногибридном скрещивании подчас отличаются от установленных им. Например, при скрещивании растений «ночной красавицы» с красными и белыми цветками все гибриды F1 имеют розовые цветки. А во втором поколении гибридов F2 наблюдается расщепление признака в соотношении 1 : 2 : 1 (растения с красными, розовыми и белыми цветками) (рис. 3). 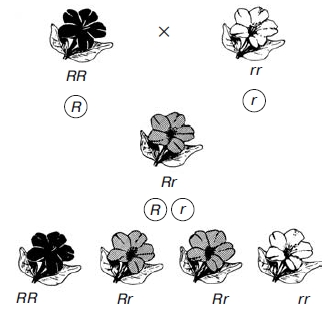 Рис. 3. Схема неполного доминирования В этом случае наблюдается промежуточный характер наследования, т. е. у гетерозиготных гибридов (Rr) не проявляется ни доминантный признак (красные цветки), ни рецессивный (белые цветки). Эта закономерность наследования получила название неполного доминирования. Кроме данного феномена были выявлены и другие закономерности наследования, отличные от законов Менделя. Следовательно, они не являются абсолютными, а имеют ограниченный характер. В современной генетике существуют понятия менделирующие признаки (наследующиеся по законам Менделя) инеменделирующие (наследующиеся по иным законам). Менделирующих признаков у всех организмов большое число. Немало их и у человека (табл. 8 и рис. 5). Таблица 8 Некоторые менделирующие признаки у человека
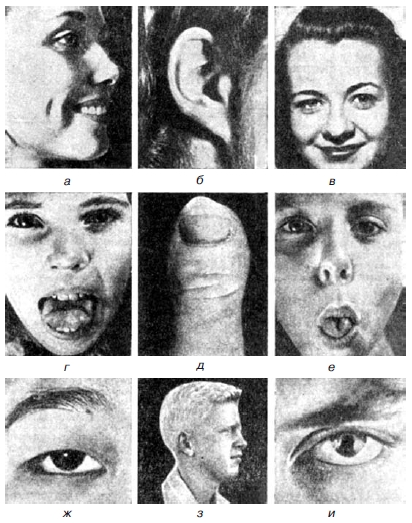 Рис. 4. Некоторые наследственные признаки человека: а — ямочки на щеках (доминантный признак); б— приросшая мочка уха (рецессивный признак); в — рост волос по средней линии лба(доминантный признак); г— способность загибать язык назад (доминантный признак); л — расплющенный большой палец (доминантный признак); е— способность свертывать язык трубочкой (доминантный признак);ж —монголоидный разрез глаз (доминантный признак); з — альбинизм (рецессивный признак); и — зрачок, заходящий на радужную оболочку (сцепленный с полом рецессивный признак) Применяемые Менделем приемы легли в основу нового метода изучения наследования — гибридологического. Гибридологический анализ — это постановка системы скрещиваний, позволяющих выявить закономерности наследования признаков. Условия проведения гибридологического анализа: 1) родительские особи должны быть одного вида и размножаться половым способом (иначе скрещивание просто невозможно); 2) родительские особи должны быть гомозиготными по изучаемым признакам; 3) родительские особи должны различаться по изучаемым признакам; 4) родительские особи скрещивают между собой один раз для получения гибридов первого поколения F1, которые затем скрещивают между собой для получения гибридов второго поколения F2; 5) необходимо проведение строгого учета числа особей первого и второго поколения, имеющих изучаемый признак. Менделирующими признаками называются те, наследование которых про исходит по закономерностям, установленным Г. Менделем. Менделирующие признаки определяются одним геном моногенно (от греч.monos-один) то есть когда проявление признака определяется взаимодействием аллельных генов, один из которых доминирует (подавляет) другой. Менделевские законы справедливы для аутосомных генов с полной пенетрантностью (от лат.penetrans-проникающий, достигающий) и постоянной экспрессивностью (степенью выраженности признака). Если гены локализованы в половых хромосомах (за исключением гомологичного участка в Х- и У-хромосомах), или в одной хромосоме сцеплено, или в ДНК органоидов, то результаты скрещивания не будут следовать законам Менделя. Общие законы наследственности одинаковы для всех эукариот. У человека также имеются менделирующие признаки, и для него характерны все типы их наследования: аутосомно-доминантный, аутосомно-рецессивный, сцепленный с половыми хромосомами (с гомологичным участком Х- и У-хромосом). Типы наследования менделирующих признаков: I. Аутосомно-доминантный тип наследования. По аутосомно-доминантному типу наследуются некоторые нормальные и патологические признаки: 1) белый локон над лбом; 2) волосы жесткие, прямые (ежик); 3) шерстистые волосы - короткие, легко секущиеся, курчавые, пышные; 4) кожа толстая; 5) способность свертывать язык в трубочку; 6) габсбургская губа - нижняя челюсть узкая, выступающая вперед, нижняя губа отвислая и полуоткрытый рот; 7) полидактилия (от греч.polus – многочисленный, daktylos- палец) – многопалость, когда имеется от шести и более пальцев; 8) синдактилия (от греч. syn - вместе)-сращение мягких или костных тканей фаланг двух или более пальцев; 9) брахидактилия (короткопалость) – недоразвитие дистальных фаланг пальцев; 10) арахнодактилия (от греч. агаhna – паук ) – сильно удлиненные «паучьи» пальцы II. Аутосомно-рецессивный тип наследования. Если рецессивные гены локализованы в аутосомах, то проявиться они могут при браке двух гетерозигот или гомозигот по рецессивному аллелю. По аутосомно-рецессивному типу наследуются следующие признаки: 1)волосы мягкие, прямые; 2)кожа тонкая; 3)группа крови Rh-; 4)неощущение горечи вкуса фенилкарбамида; 5)неумение складывать язык в трубочку; 6)фенилкетонурия – блокируется превращение фенилаланина в тирозин, который превращается в фенилпировиноградную кислоту, являющуюся нейротропным ядом (признаки – судорожные синдромы, отставание в психическом развитии, импульсивность, возбудимость, агрессия); 7)галактоземия - накопление в крови галактозы, которая тормозит всасывание глюкозы и оказывает токсическое действие на функцию печени, мозга, хрусталика глаза; 8)альбинизм. Частота рецессивных наследственных болезней особенно повышается в изолятах и среди населения с высоким процентом кровнородственных браков. III. Менделирующие признаки, сцепленные с полом (неполно). Х и У-хромосомы имеют общие гомологичные участки. В них локализованы гены, детерминирующие признаки, наследующиеся одинаково как у мужчин, так и у женщин (подобно признакам, сцепленным с аутосомами). Гены, локализованные в гомологичных участках Х- и У-хромосом, обусловливают развитие некоторых болезней. 1)пигментная ксеродерма - заболевание, при котором под влиянием ультрафиолетовых лучей на открытых частях тела появляются пигментированные пятна. Вначале они в виде веснушек, затем в виде более крупных папиллом различной величины и, наконец, опухолей. Для большинства больных пигментная ксеродерма заканчивается летально 2)болезнь Огучи – в слое палочек и колбочек, пигментом эпителии наблюдаются дегенеративные изменения (болезнь чаще встречается в Японии). 3)спастическая параплегия – спастика и слабость нижних конечностей, возникающая в результате дегенерации пирамидных путей в области грудного и поясничного отделов спинного мозга, изредка в стволе головного мозга, изредка в стволе головного мозга. 4)эпидермолиз буллезный – образование пузырей после механических травм кожи. 5)полная (общая) цветовая слепота – полное отсутствие цветового зрения. ложенных на Наследование групп крови. Большое значение для медицинской практики имеет изучение групп крови, которые зависят от антигенов, распоповерхности эритроцитов. Антигены – это высокомолекулярные вещества, в ответ на введение которых в организме вырабатываются антитела (гамма-глобулины – одна из фракций белков крови, которая синтезируется лимфоцитами). Следует отметить, что на собственные антигены организм с нормальной иммунной системой антител не вырабатывает. В настоящее время хорошо изучены группы крови систем: АВ0, Rh, МN, Р, Даффи, Льюис, Лютеран, Келл, Кидд и др. В систему входят группы крови, которые детерминируются (определяются) аллелями одного гена. Множественные аллели - количество аллелей в природе больше двух. Одним из примеров множественных аллелей у человека являются группы крови системы АВ0. В зависимости от антигенов, которые находятся на поверхности эритроцитов, все люди земного шара делятся на четыре группы. У одних людей на поверхности эритроцитов нет антигенов А и В - это 0 (I) группа, у других есть антиген А -А(П) группа, у третьих есть антиген В - В (III) группа, а у четвертых есть антигены А и В - АВ (IV) группа. В процессе длительной эволюции живые организмы приспособились к сохранению постоянства своего антигенного состава и не допускают вмешательства других антигенов. Поэтому у людей 0(1) группы крови, не имеющей на поверхности антигенов А и В, есть антитела α и β против антигенов А и В; у людей А (II) группы крови есть антитела β против антигена В; у людей В(Ш) группы есть антитела α против антигена А; у АВ (IV) группы нет антител против антигенов А и В. Четыре группы крови (системы АВ0) определяются аллельными генами, которые располагаются в девятой паре хромосом человека. Обозначаются аллельные гены разными буквами алфавита (IА, IВ, I°), как исключение из правил генетики. 0(I), А(П) и В(Ш) группы наследуются как менделирующие признаки. Гены IА и IВ по отношению к гену I° ведут себя доминантно. Аллельные гены IА и IВ у лиц IV группы ведут себя независимо друг от друга: ген IА детерминирует антиген А, а ген IВ - антиген В. Такое взаимодействие аллельных генов называется кодоминированием (каждый аллель детерминирует свой признак). Наследование АВ(IV) группы крови не следует закономерностям, установленным Менделем. Группы крови А(II) и В(Ш) системы АВ0 наследуются по аутосомно-до-минантному типу, а 0(I) группа - по аутосомно-рецессивному типу. Рассмотрим, как наследуются группы крови системы АВ0. 1). Если гомозиготная женщина А(П) группы крови выйдет замуж за мужчину с 0(I) группой, то все дети будут А(П) группы крови. P -♀ IА IА × I° I° ♂ G - IА I° F - IА I° - 100 % А(II) группы 2). Женщина А(П) группы крови гетерозиготная вышла замуж за мужчину с 0(I) группой крови. Вероятность рождения детей будет: 50 % с 0(I) группой и 50 % с А(П) группой крови. P -♀ IА I° × I° I° ♂ G - IА I° I° F - IА I° I° I° 50% А(II) 50% 0(I) 3) Если женщина В(Ш) группы крови гомозиготная, а мужчина 0(I) группы, то все дети будут В(Ш) группы гетерозиготные. P -♀ IВ IВ × I° I° ♂ G - IВ I° F - IВ I° - 100 % В(Ш) группа 4) Женщина В(Ш) группы крови гетерозиготная, а мужчина 0(I) группы крови. Вероятность рождения детей от этого брака составит: 50 % В(Ш) группы крови гетерозиготных на 50% 0(I) группы крови. P -♀ IВ I° × I° I° ♂ G - IВ I° I° F - IВ I° I° I° 50 % В(Ш) 50% 0(I) 5)Если женщина А(II) группы крови вышла замуж за мужчину В(Ш) группы крови (оба гомозиготные), то от этого брака все дети будут АВ (IV) группы крови. P -♀ IА IА × IВ IВ♂ G - IА × IВ F - IА IВ- 100 % АВ (IV) группа крови 6)Женщина А(II) группы крови вышла замуж за мужчину В(Ш) группы (оба гетерозиготные). От этого брака равновероятно рождение детей 0(I), А(II), В(Ш), АВ (IV) группы крови, так как происходит случайная встреча гамет родителей свободная комбинация генов. P -♀ IА I° × IВ I° ♂ G - IА I° IВ I° F - IА IВ , IВ I° , IА I° , I° I°. АВ (IV) В(Ш) А(II) 0 Кроме антигенов А, В, 0 на поверхности эритроцитов у людей расположены антигены групп системы резус. Если на эритроцитах находится антиген Rh, то такие люди относятся к группе Rh+ (их около 85 %), а если отсутствует данный антиген, то они относятся к группе Rh¯ (их около 15 %). Группы крови Rh+ и Rh¯ системы резус детерминируются генами, которые локализованы в первой паре хромосом человека. Группа крови Rh+ может быть гомозиготная (DD) и гетерозиготная (Dd), группа Rh¯ - только гомозиготная (dd). Группы крови резус-системы наследуются как менделирующие признаки. Проследим, какие могут быть последствия для детей, если мать имеет резус-отрицательную группу крови. Женщина с группой крови Rh¯ вышла замуж за мужчину, у которого группа крови Rh+ гомозиготная. Р - ♀ dd × DD♂ G - d D F – Dd – 100% От этого брака все дети будут резус-положительные гетерозиготные, так как ген D полностью доминирует над геном d и F - - единообразно. Во время беременности Rh+ эритроциты плода могут попасть в кровь матери, и материнский организм начнет выработку антител против этих эритроцитов. С каждой последующей беременностью увеличивается риск иммунизации и возрастает вероятность гемолитической болезни новорожденных и ее тяжести. Если женщина с группой Rh¯ вступает в брак с мужчиной гетерозиготным (что чаще встречается), то вероятность рождения детей от этого брака будет равна 50 % Rh+ гетерозиготных и 50 % Rh¯. Р - ♀ dd × Dd♂ G - d D d F – Dd,dd Вопрос 33( Сцепление генов. Кроссинговер, Сцепление генов, совместная передача двух или более генов от родителей потомкам. Объясняется тем, что эти гены лежат в одной хромосоме, то есть принадлежат одной группе сцепления и поэтому не могут случайно перекомбинироваться в мейозе, как это бывает при наследовании генов, лежащих в разных хромосомах. С. г. было открыто в 1906 английскими генетиками У. Бэтсоном и Р. Пеннетом, обнаружившими в опытах по скрещиванию растений у некоторых генов тенденцию передаваться совместно и тем самым нарушать закон независимого комбинирования признаков (см. Менделя законы, Менделизм). Правильное объяснение этому дали Т.Моргани сотрудники, обнаружившие аналогичное явление при изучении наследования признаков у дрозофилы. Мерой С. г. служит частота образования гетерозиготой по этим генам кроссоверных гамет или спор, в которых гены находятся не в исходных, а в новых сочетаниях благодаря обмену частями несущих их гомологичных хромосом путём кроссинговера. У некоторых бактерий др. мерой С. г. служит частота совместной передачи по наследству разных генов при конъюгации, генетической трансформации и трансдукции. Сила С. г. может быть различной у разных полов (обычно она больше у гетерогаметного пола, см. Половые хромосомы) или даже С. г. может быть полным (отсутствие кроссинговера) у одного из полов (например, у самцов дрозофилы или у самок тутового шелкопряда). Кроме того, сила С. г. может варьировать в зависимости от возраста родителей, температуры, наличия хромосомных перестроек и др. факторов, а также от присутствия особых мутантных генов, специфически влияющих на силу С. г. См. также Генетические карты хромосом. Кроссинговер Кроссинговер (от англ. crossing–over – перекрёст) – это обмен гомологичными участками гомологичных хромосом (хроматид). Само явление перекрёста хромосом открыл Ф. Янссенс (1909, Бельгия), обнаруживший хиазмы в профазе I мейоза у саламандры. Однако теоретическая возможность кроссинговера была предсказана раньше. Во-первых, американский цитолог У. Сэттон (1903) предположил, что в одной хромосоме может находиться несколько генов. В этом случае должно наблюдаться сцепленное наследование признаков, т.е. несколько разных признаков могут наследоваться так, как будто они контролируются одним геном. Тогда совокупность генов в одной хромосоме образует группу сцепления. Во-вторых, в 1906 г. У. Бэтсон и Р. Пеннет обнаружили сцепленное наследование у душистого горошка. Они изучали совместное наследование: окраски цветков (пурпурная или красная) и формы пыльцевых зерен (удлиненная или округлая). При скрещивании дигетерозигот в их потомстве наблюдалось расщепление 11,1:0,9:0,9:3,1 вместо ожидаемого 9:3:3:1. Таким образом, два признака (окраска цветков и морфология пыльцы) оказываются сцепленными. Соответственно, сцеплены и гены, контролирующие эти признаки («эффект взаимного притяжения»). Однако это сцепление не абсолютно: гены, контролирующие сцепленные признаки, могут приобретать независимость, и тогда появляются новые комбинации признаков. Вскоре (1911) были получены данные по сцепленному наследованию и нарушению сцепления некоторых признаков у дрозофилы. Т. Морган (1910) и его ученик К. Бриджес (1916) доказали, что число групп сцепления соответствует гаплоидному числу хромосом. Опираясь на работы Янссенса и результаты гибридологического анализа, они связали явление перекрёста хромосом с феноменом нарушения сцепления генов. При перекрёсте хромосом происходит обмен между ними генетическим материалом (аллелями), и тогда происходит рекомбинация – появление новых сочетаний аллелей, например, AB + ab → Ab + aB. Таким образом, термин «кроссинговер» употребляется и в цитологическом, и в генетическом значении. В дальнейшем связь между перекрестом хромосом и появлением новых сочетаний признаков была подтверждена работами К. Дарлингтона, К. Штерна, Б. Мак-Клинток и др. | ||||||||||||||||||||||||||||