Каталитическая функция. Большинство известных в настоящее время ферментов, называемых биологическими катализаторами, является белками. Транспортная функция
 Скачать 1.95 Mb. Скачать 1.95 Mb.
|

|
Метаболизм или обмен веществ — набор химических реакций, которые возникают в живом организме для поддержания жизни. Эти процессы позволяют организмам расти и размножаться, сохранять свои структуры и отвечать на воздействия окружающей среды. Метаболизм обычно делят на две стадии: в ходе катаболизма сложные органические вещества деградируют до более простых; в процессах анаболизма с затратами энергии синтезируются такие вещества, как белки, сахара, липиды и нуклеиновые кислоты. Обмен веществ происходит между клетками организма и межклеточной жидкостью, постоянство состава которой поддерживается кровообращением: за время прохождения крови в капиллярах через проницаемые стенки капилляров плазма крови 40 раз полностью обновляется с интерстициальной жидкостью. Серии химических реакций обмена веществ называют метаболическими путями, в них при участии ферментов одни биологически значимые молекулы последовательно превращаются в другие. Основные метаболические пути и их компоненты одинаковы для многих видов. Например, некоторые карбоновые кислоты, являющиеся интермедиатами цикла трикарбоновых кислот присутствуют во всех организмах Катаболические, анаболические пути. Ферментативное расщепление основных питательных веществ, а именно углеводов, жиров и белков, происходит в клетке через ряд последовательных ферментативных реакций. Катаболизм основных питательных веществ включает три главные стадии. На первой стадии крупные пищевые молекулы расщепляются на составляющие их основные строительные блоки. Полисахариды, например, расщепляются до гексоз или пентоз, липиды — до жирных кислот, глицерина и других компонентов, белки — до аминокислот, которых имеется 20 видов. На второй стадии большое число продуктов, образовавшихся на первой стадии, превращается в более простые молекулы, число типов которых сравнительно невелико. Так, гексозы, пентозы и глицерин, разрушаясь, превращаются сначала в трехуглеродный фосфорилированный сахар — глицеральдегид-3-фосфат, а затем расщепляются далее до единственной двууглеродной формы — ацетильной группы, входящей в состав ацетил- КоА. Двадцать различных аминокислот также дают при расщеплении лишь несколько конечных продуктов, а именно ацетил-КоА, α-кетоглутарат, сукцинат, фумарат и оксалоацетат. Продукты, образовавшиеся на второй стадии, вступают в третью стадию, которая для них всех является общей и на которой они в конечном счете окисляются до CO2 и воды. Процесс анаболизма также включает три стадии. Исходными веществами, или строительными блоками служат для него соединения, поставляемые третьей стадией катаболизма. Таким образом, третья стадия катаболизма является в то же время первой, исходной, стадией анаболизма. Синтез белка, например, начинается на этой стадии с α - кетокислот, являющихся предшественниками α -аминокислот. На второй стадии анаболизма α -кетокислоты аминируются аминогруппой доноров с образованием α -аминокислот, а на третьей, заключительной, стадии аминокислоты объединяются в пептидные цепи. Продукты метаболизма выводятся с мочой, фекалиями, выдыхаемым воздухом и потом. Специфические вещества задерживаются или удаляются из организма в той степени, в какой это необходимо для поддержания гомеостаза, при этом вместе со шлаковыми продуктами распада выводятся потенциально полезные вещества. Небольшие количества этих веществ выводятся из организма в виде кишечных газов, волос, ногтей, десквамированного эпителия кожи, кожного сала, ушной серы, слизи из полости носа и влагалища, слюны, слез, семенной жидкости и менструальных выделений. Уровни потерь этих веществ опубликованы в работах. 43. Митохондриальное окисление. Дегидрирование субстратов и окисление водорода как источник энергии для синтеза АТФ. Структура и функции переноса электронов, окислительно-восстановительный потенциал структурных компонентов цепи. Нарушение функции цепи переноса электронов при гипоксиях и других патологических состояниях. Система митохондриального окисления - мультиферментная система, постепенно транспортирующая протоны и электроны на кислород с образованием молекулы воды. Все ферменты митохондриального окисления встроены во внутреннюю мембрану митохондрий. Только первый переносчик протонов и электронов - никотинамидная дегидрогеназа расположена в матриксе митохондрии. Этот фермент отнимает водород от субстрата и передает его следующему переносчику. Полный комплекс таких ферментов образует «дыхательную цепь», в пределах которого атомы водорода отнимаются от субстрата, затем передаются последовательно от одного переносчика к другому, и, наконец, передаются на кислород воздуха с образованием воды. Существует строгая последовательность работы каждого звена в цепочке переносчиков. Эта последовательность определяется величиной редокс-потенциала (окислительно-восстановительного потенциала, сокращенно - овп) каждого звена. ОВП - это химическая характеристика способности вещества принимать и удерживать электроны. Вещества с положительным ОВП окисляют водород (отнимают от него электроны), вещества с отрицательным ОВП окисляются самим водородом. Самый низкий ОВП имеет начальное звено цепи, самый высокий - у кислорода, расположенного в конце цепочки переносчиков. Таким образом, передача водорода идет от более низкого к более высокому ОВП. На одной из стадий происходит разделение атомов водорода на Н+ и электроны. Протоны остаются временно в окружающей среде, а электроны идут дальше по цепи и в ее конце используются для активации О2. Кислород является конечным акцептором электронов. O2 + 4e -----> 2O-2 (полное восстановление кислорода). Все реакции, происходящие в дыхательной цепи, сопряжены. Переносчики водорода и электронов расположены в строгом порядке, в соответствии с величиной их редокс-потенциала. В настоящее время различают три варианта дыхательных цепей: Главная дыхательная цепь - это три мультиферментных комплекса, встроенных во внутреннюю мембрану митохондрии. Обозначаются они латинскими цифрами – I, III и IV. Сокращенная (укороченная) дыхательная цепи. Известны два субстрата. Главным из них является сукцинат (янтарная кислота). Сукцинатдегидрогеназа – это комплекс II, который в укороченном варианте цепи является начальным звеном окисления. В составе комплекса – простетическая группа ФАД и FeSII. От ФАД.Н2 два атома водорода переносятся на KoQ. Значит, первое звено, которое имеется в полной цепи - исключается. Перепад окислительно-восстановительного потенциала между ФАД и KoQ невелик. Поэтому переноса H+ в межмембранное пространство в этой точке не происходит. + также создается, но меньший, чем в полной цепи. Значит, меньше и эффективность фосфорилирования - коэффициент Р/О=2. Аналогичным образом окисляется и второй субстрат – ацил-КоА (активная форма любой жирной кислоты): Максимально сокращенная (максимально укороченная) дыхательная цепь. Она представлена только цитохромной частью. Эксперименты показали, что здесь может быть окислен только один субстрат - аскорбиновая кислота, с участием фермента, восстановленные эквиваленты включаются в цепь на уровне цитохрома С цитохромоксидазы (цитохром аа3), но в реальных условиях такого окисления практически не происходит. Образуется вода и 1 молекула АТФ. Коэффициент Р/О=1. 44. Окислительное фосфорилирование, его сопряжение с дыхательной цепью. Химиоосмотическая теория Митчела. Коэффициент Р/О. Разобщение тканевого дыхания и окислительного фосфорилирования. Биологические разобщители как регуляторы окислительного фосфорилирования. Нарушения энергетического обмена: гипоксические состояния. Окислительное фосфорилирование и дыхательный контроль. Функция дыхательной цепи – утилизация восстановленных дыхательных переносчиков, образующихся в реакциях метаболического окисления субстратов (главным образом в цикле трикарбоновых кислот). Каждая окислительная реакция в соответствии с величиной высвобождаемой энергии ≪обслуживается≫ соответствующим дыхательным переносчиком: НАДФ, НАД или ФАД. Соответственно своим окислительно-восстановительным потенциалам эти соединения в восстановленной форме подключаются к дыхательной цепи. В дыхательной цепи происходит дискриминация протонов и электронов: в то время как протоны переносятся через мембрану, создавая ΔрН, электроны движутся по цепи переносчиков от убихинола к цитохромоксидазе, генерируя разность электрических потенциалов, необходимую для образования АТФ протонной АТФ-синтазой. Таким образом, тканевое дыхание ≪заряжает≫ митохондриальную мембрану, а окислительное фосфорилирование ≪разряжает≫ ее. Разность электрических потенциалов на митохондриальной мембране, создаваемая дыхательной цепью, которая выступает в качестве молекулярного проводника электронов, является движущей силой для образования АТФ и других видов полезной биологической энергии. Механизмы этих превращений описывает хемиосмотическая концепция превращения энергии в живых клетках. Она была выдвинута П. Митчеллом в 1960 г. для объяснения молекулярного механизма. Согласно хемиосмотической концепции, движение электронов по дыхательной цепи является источником энергии для транслокации протонов через митохондриальную мембрану. Возникающая при этом разность электрохимических потенциалов (ΔμH+) приводит в действие АТФ-синтазу, катализирующую реакцию АДФ + Рi = АТФ. В дыхательной цепи есть только 3 участка, где перенос электронов сопряжен с накоплением энергии, достаточным для образования АТФ, на других этапах возникающая разность потенциалов для этого процесса недостаточна. Максимальная величина коэффициента фосфорилирования, таким образом, составляет 3, если реакция окисления идет с участием НАД, и 2, если окисление субстрата протекает через флавиновые дегидрогеназы. Теоретически еще одну молекулу АТФ можно получить в трансгидрогеназной реакции (если процесс начинается с восстановленного НАДФ): Обычно в тканях восстановленный НАДФ используется в пластическом обмене, обеспечивая разнообразные синтетические процессы, так что равновесие трансгидрогеназной реакции сильно сдвинуто влево. Эффективность окислительного фосфорилирования в митохондриях определяется как отношение величины образовавшегося АТФ к поглощенному кислороду: АТФ/О или Р/О (коэффициент фосфорилирования). Экспериментально определяемые значения Р/О, как правило, оказываются меньше 3. Это свидетельствует о том, что процесс дыхания не полностью сопряжен с фосфорилированием. Действительно, окислительное фосфорилирование в отличие от субстратного не является процессом, в котором окисление жестко сопряжено с образованием макроэргов. Степень сопряжения зависит главным образом от целостности митохондриальной мембраны, сберегающей разность потенциалов, создаваемую транспортом электронов. По этой причине соединения, обеспечивающие протонную проводимость (как 2,4-динитрофенол), являются разобщителями. 45. Свободно-радикальное окисление. Токсичность кислорода: образование активных форм кислорода, их действие на липиды и другие вещества клетки. Механизм защиты: антиоксидантные системы Свободнорадикальное окисление – важный и многогранный биохимический процесс превращений кислорода, липидов, нуклеиновых кислот, белков и других соединений под действием свободных радикалов, а перекисное окисление липидов (ПОЛ) – одно из его последствий. Свободные радикалами (СР) представляют собой соединения, имеющие неспаренный электрон на наружной орбите и обладающие высокой реакционной способностью. К числу первичных СР относятся супероксидный анион-радикал, окись азота, а вторичными СР являются гидроксильный радикал, синклетный кислород, перекись водорода, пероксинитрит. Образование СР тесно связано, с одной стороны, с появлением свободных электронов при нарушениях процессов окисления в дыхательной цепи, превращении ксантина, синтезе лейкотриенов и простогландинов. Эти реакции зависят от активности ксантиноксидазы, дегидроротатдегидрогеназы, льдегидоксидазы, холестериноксидазы, ферментов цитохрома Р-450. Синтез супероксид — аниона инициируется ангиотензином II, который образуется из ангиотензина I под действием ангиотензинпревращающего фермента. Его активность в сосудах больных с выраженным атеросклерозом особенно высока. Причина этого явления пока неясна. Оксид азота образуется при окисленииL-аргинина под действием NO-синтетазы при участии кальмодулина. В литературе представлены данные о том, что синтез NO при атеросклерозе не нарушен, но его дефицит может возникать при увеличении его соединения с супероксид – анионом. Оно завершается синтезом пероксинитрита с большими патогенными потенциями, в том числе и образованием окисленных форм ЛПНП. С другой стороны, для синтеза вторичных СР используется водород НАДФ-Н, НАД-Н — его донаторов. Супероксид — анион может восстанавливать Fe3+ в Fe2+, при взаимодействии которого с перекисью водорода, перекисями липидов и гипохлоритом образуются высокотоксичные вторичные радикалы. Из всех СР наибольшей активностью обладают гидроксильный радикал и пероксинитрит. Активность СР ограничивается антиоксидантами, которые разрывают цепи молекул при реакциях СРО, разрушают молекулы перекисей. К числу ферментных антиоксидантов относятся супероксиддисмутаза (СОД), глютатионпероксидаза, каталаза, находящиеся в клеточных структурах. Неферментные антиоксиданты – витамины Е, К, С, убихиноны, триптофан, фенилаланин, церулоплазмин, трансферрин, гаптоглобин, глюкоза, каротиноиды – блокируют активность СР в крови. 46. Микросомальное окисление. Структурные компоненты и функции оксидаз, монооксигеназ и диоксигеназ. Значение микросомального окисления для организма: обезвреживание природных и искусственных экотоксикантов. Свободное окисление. Одна из задач свободного (несопряженного) окисления – превращения природных или неприродных субстратов, называемых в этом случае ксенобиотиками (ксено – несовместимый, биос – жизнь). Они осуществляются ферментами диоксигеназами и монооксигеназами. Окисление протекает при участии специализированных цитохромов, локализованных чаще всего в эндоплазматическом ретикулуме, поэтому иногда этот процесс называют микросомальным окислением. В реакциях свободного окисления участвуют также кислород и восстановленные дыхательные переносчики (чаще всего НАДФН). Акцептором электронов является цитохром Р-450 (иногда цитохром b5). Окисление субстрата протекает по следующей схеме: SH + O2 –> SOH. Механизм действия оксигеназ включает изменение валентности входящих в их состав ионов двухвалентных металлов (железа или меди). Диоксигеназы присоединяют к субстрату молекулярный кислород, активируя его за счет электрона атома железа в активном центре (железо при этом становится трехвалентным). Оксигенация протекает как атака субстрата образующимся супероксид-анионом кислорода. Одной из биологически важных реакций такого типа является превращение β-каротина в витамин А. Монооксигеназы требуют участия в реакции НАДФН, атомы водорода которого взаимодействуют с одним из атомов кислорода, поскольку только один электрон связывается с субстратом. К широко распространенным монооксигеназам относятся разнообразные гидроксилазы. Они принимают участие в окислении аминокислот, оксикислот, полиизопреноидов. В процессе свободного окисления вследствие особенностей используемых цепей передачи электронов не происходит образования АТФ; биологическая роль этих процессов заключается в метаболизме ряда природных и ксенобиотических субстратов. В последнем случае свободное окисление выполняет важную функцию модификации чужеродных соединений. 47. Основные углеводы пищи. Переваривание углеводов в ротовой полости и желудочно-кишечном тракте, амилолитические ферменты слюны, поджелудочной железы, гидролиз дисахаридов. Всасывание моносахаридов (механизм всасывания). По количеству углеводных остатков УВ разделяются на 3 основных класса: 1. моносахариды (глюкоза, фруктоза, галактоза, манноза и др.); 2. дисахариды (мальтоза, сахароза, лактоза); 3. полисахариды (гомополисахариды крахмал, гликоген, клетчатка). Крахмал – резервный гомополисахарид растений, построенный из остатков α-глюкозы. Крахмал представляет собой смесь двух гомополисахаридов: амилозы и амилопектина. В амилозе остатки глюкозы связаны α-1,4-гликозидными связями, в точках ветвления амилопектина - α-1,6-гликозидными связями. Молекулярная масса крахмала – 106-107. Гликоген – резервный гомополисахарид высших животных и человека, построенный из остатков α–Д-глюкозы. Гликоген содержится практически во всех органах и тканях человека и животных; наибольшее его количество находится в печени и мышцах. Молекулярная масса гликогена 107 – 109 и выше. Гликоген по своему строению близок к амилопектину. Остатки глюкозы связаны α-1,4-гликозидными и α-1,6-гликозидными связями (в точках ветвления). В молекуле гликогена имеется большее число α-1,6-гликозидных связей по сравнению с крахмалом. Клетчатка – это единственный гомополисахарид, который не переваривается в ЖКТ человека, т.к. пищеварительные железы человека не вырабатывают β–глюкозидазу. Однако клетчатка выполняет ряд важных функций: 1. способствует формированию кала; 2. усиливает перистальтику кишечника; 3. является адсорбентом, с которым из кишечника выводятся излишки холестерина, соли тяжелых металлов. Роль углеводов: 1. Энергетическая (глюкоза – основной источник энергии для организма. При сгорании 1 г УВ выделяется 4 ккал энергии). 2. Структурно-функциональная (УВ – обязательный компонент гликопротеинов и протеогликанов, которые выполняют разнообразные функции: гормональную, рецепторную, защитную, ферментативную и др.). 3. Метаболическая (пентозы участвуют в синтезе нуклеиновых кислот и нуклеотидных коферментов). Суточная потребность в углеводах составляет 400-500 г. Переваривание углеводов - это совокупность процессов поэтапного ферментативного гидролиза полисахаридов до моносахаридов, которые всасываются в кишечнике, разносятся током крови к печени и другим тканям организма, где подвергаются различным метаболическим превращениям. Переваривание УВ начинается в ротовой полости под действием фермента амилазы слюны (оптимум рН=6,8-7,2), которая гидролизует в крахмале α -1,4-гликозидные связи с образованием декстринов. Всасывание УВ из просвета кишечника в энтероцит и из энтероцита в кровь происходит: 1) облегченной диффузией с участием переносчиков; 2) вторично активным транспортом (симпортом с ионами натрия) с использованием энергии K, Na-АТФ-азы. Быстрее всех всасываются глюкоза и галактоза. От кишечника всосавшиеся моносахариды транспортируются в печень, где происходит до 90% превращений моносахаров. Поступление глюкозы в клетки сердечной, скелетных мышц и жировой ткани регулируется инсулином. 48. Потребность ребенка в углеводах. Своеобразие переваривания и всасывания углеводов в желудочно-кишечном тракте у ребенка.  49. Обмен галактозы, фруктозы, маннозы. Взаимопревращение моносахаров в организме. Наследственные нарушения обмена моносахаридов: галактоземия, непереносимость фруктозы. 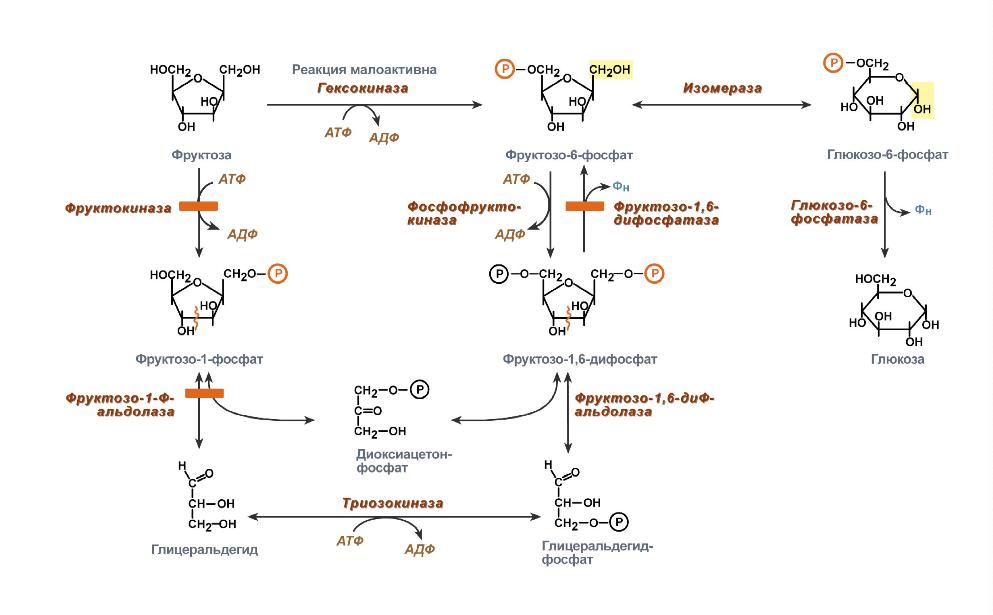 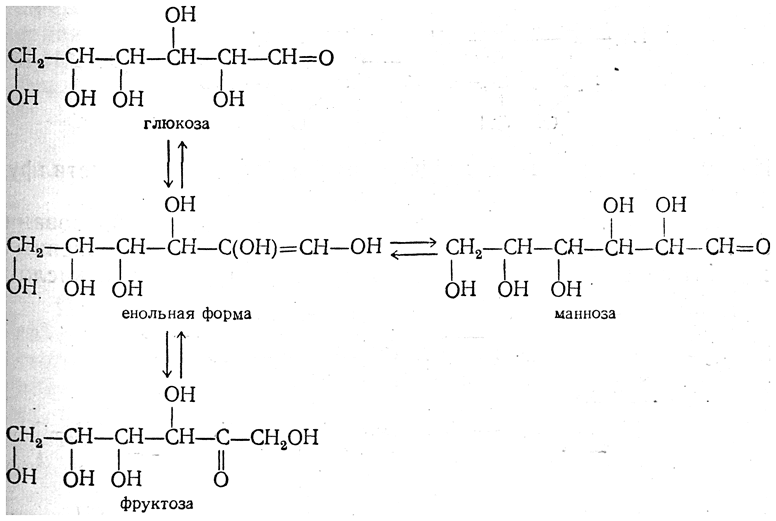 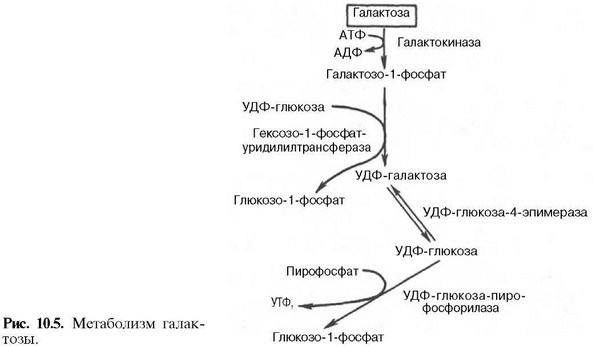 Нарушения, связанные с недостаточностью ферментов, которые принимают участие в переваривании и всасывании УВ. 1. Непереносимость лактозы: а) врожденная. Дефект по лактазе в просвете тонкого кишечника. Большой осмотический эффект невсосавшейся лактозы вызывает приток жидкости в тонкий кишечник, поэтому клиническими симптомами являются вздутие живота, тошнота, судороги, боль и водная диарея. Наследуется по аутосомно-рецессивному типу. б) приобретенная (временная). Может быть, и у взрослых вследствие инфекционных заболеваний или интенсивного лечения антибактериальными препаратами. 2. Нарушения обмена фруктозы: а) Фруктоземия (наследственная непереносимость фруктозы). Недостаточность фермента кетозо-1- фосфат-альдолазы приводит к резкому увеличению концентрации фруктозо-1-фосфата в клетках печени, вследствие чего развиваются хроническая недостаточность функций печени и почек, гипогликемия, диарея, рвота, боли в животе. б) Эссенциальная фруктозурия. Причина: недостаток фруктокиназы. Следствие: нарушение фосфорилирования фруктозы приводит к повышению содержания фруктозы в крови и аномальному выведению фруктозы с мочой. Это нарушение не вызывает патологических симптомов. 3. Нарушения обмена галактозы: а) Галактоземия. Причина: недостаток фермента гексозо-1-фосфатуридилтрансфераза печени. Больные дети плохо растут, прием молока вызывает рвоту и понос. Происходит увеличение печени и желтуха. При этом заболевании увеличивается концентрация галактозы и галактозо-1-фосфата. Галактоземия сопровождается галактозурией. У детей галактоземия приводит к умственной отсталости и катаракте хрусталика. Определяющим диагностическим критерием служит отсутствие в эритроцитах гексозо-1-фосфатуридилтрансферазы. б) Недостаток галактокиназы. Происходит накопление галактозы и ее превращение в галактитол. Следствие: раннее развитие катаракты. 50. Анаэробный распад глюкозы. Этапы этого процесса. Гликолитическая оксиредукция, субстратное фосфорилирование. Энергетическая ценность анаэробного распада глюкозы. Регуляторные механизмы, участвующие в этом процессе. 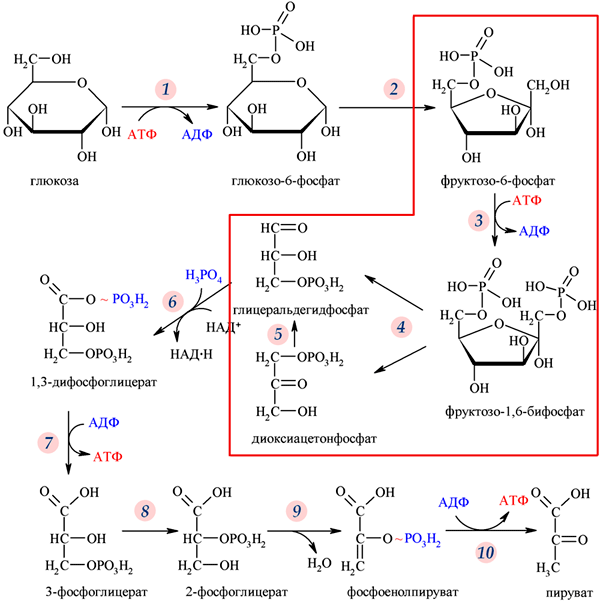 Гликолиз – синоним молочнокислого брожения – сложный ферментативный процесс превращения глюкозы до двух молекул молочной кислоты, протекающий в тканях человека и животных без потребления кислорода. Гликолиз включает 11 ферментативных реакций, протекающих в цитоплазме клетки. Реакции гликолиза проходят в 2 стадии. В ходе первой стадии – энергопотребляющей – используются 2 АТФ в 1-ой и 3-ей реакциях. В процессе 7-ой и 10-ой реакций второй стадии – энергодающей – образуются 4 АТФ. Из 11 реакций - 3 необратимые (1-ая, 3-я и 10-ая). 51. Анаэробный гликолиз, повышенная активность анаэробного распада глюкозы у ребенка раннего возраста. Биохимические критерии гликолиза. Связь с возникновением ацидоза. |