|
|
Каталитическая функция. Большинство известных в настоящее время ферментов, называемых биологическими катализаторами, является белками. Транспортная функция
Значение анаэробного гликолиза: 1. каждая молекула глюкозы в анаэробных условиях снабжает клетку 2-мя молекулами АТФ; 2. промежуточные метаболиты гликолиза (фосфоглицериновый альдегид, фосфодиоксиацетон, фосфоглицерат, пируват) могут использоваться клеткой в обмене липидов и белков; 3. пируват и НАДН используются в реакциях аэробного окисления глюкозы; 4. восемь реакций гликолиза из 11 обратимые, поэтому возможен синтез глюкозы из лактата (глюконеогенез). Эффект Пастера. Подавление анаэробного гликолиза аэробным окислением глюкозы. Факультативные анаэробы потребляют больше глюкозы, т.к. энергоотдача гликолиза – всего 2 АТФ. При добавлении О2 в среду потребление глюкозы резко уменьшается, т.к. возрастает количество АТФ, которое блокирует активность гексокиназы и фосфофруктокиназы. Спиртовое брожение. В процессе спиртового брожения молекула глюкозы в анаэробных условиях превращается в пируват, который затем декарбоксилируется и восстанавливается при участии НАДН с образование этанола. Особенности гликолиза и его нарушения у детей. У новорожденного аэробные процессы преобладают над аэробными. Это продолжается в течение первого месяца, пока фетальный Hb F не заменится на взрослый HbА. У детей высокая скорость аэробных процессов, в ходе которых вырабатывается АТФ, необходимая для синтетических процессов и роста. Детям требуется больше УВ в пище в качестве источника энергии. В детском организме анаэробные механизмы распада глюкозы и получения энергии подключаются при стрессах быстрее, чем у взрослых. Это свидетельствует о лучшей адаптации организма к экстремальным ситуациям. В подростковом возрасте еще не совершенны процессы гормональной регуляции (действие инсулина, глюкагона, адреналина). Распад гликогена может идти быстро, расход глюкозы большой. Но если не восполняется количество глюкозы в крови, легко наступает гипогликемия.
52. Аэробный распад глюкозы: этапы этого процесса и его энергетика. Физиологическое значение аэробного распада глюкозы.
Аэробный путь окисления глюкозы и его энергетика. Аэробный путь окисления глюкозы происходит в 3 этапа: 1. окисление молекулы глюкозы до 2 молекул пирувата; 2. окислительное декарбоксилирование пирувата (с образованием 2 молекул ацетил-КоА); 3. реакции цикла Кребса, где каждая молекула ацетил-КоА сгорает с образованием 12 АТФ. 1-ый этап. Окисление молекулы глюкозы до 2 молекул пирувата - это первые 10 реакций гликолиза. Суммарная реакция 1 этапа:
Глюкоза + 2 НАД++ 2 АДФ → 2 пирувата + 2 НАДН + Н+ + 2 АТФ
2-ой этап. Окислительное декарбоксилирование пирувата (с образованием 2 молекул ацетилКоА) происходит в матриксе митохондрий и осуществляется мультиферментным комплексом пируватдегидрогеназой. В его состав входят 3 фермента и 5 коферментов: 1. Пируватдекарбоксилаза – кофермент тиаминпирофосфат (активная форма витамина В1). 2. Дегидролипоилацетилтрансфераза – кофермента: в активном центре – липоевая кислота (витаминоподобное вещество), из матрикса митохондрий используется Коэнзим А (КоА
SH). 3. Дигидролипоилдегидрогеназа – коферменты: в активном центре – ФАД, из матрикса митохондрий используется – НАД. Суммарная реакция:
2 Пируват + 2 НАД+ + КоА → 2 ацетил-КоА + 2 НАДН + Н+
Каждый восстановленный НАД в митохондриях отдает протоны и электроны в полную ц.п.э., где образуется 2 х 3 АТФ = 6 АТФ. Таким образом, ценность второго этапа – 6 АТФ. Эти реакции происходят только в аэробных условиях! 3-ий этап. Реакции цикла Кребса, где каждая молекула ацетил-КоА сгорает с образованием 12 АТФ. Следовательно, энергетический выход третьего этапа (в расчете на молекулу глюкозы) – 2 х 12 АТФ = 24 АТФ . Энергетическая ценность аэробного окисления глюкозы: 1 этап – 8 (6) АТФ 2 этап – 6 АТФ 3 этап – 24 АТФ Итого: 38 (36) АТФ (разница в 2 АТФ зависит от челнока, который переносит протоны и электроны из цитозоля от НАДН, образованного в 6-ой реакции гликолиза в митохондрии в ц.п.э.). Аэробное окисление глюкозы является главным источником энергии для большинства клеток организма.
53. Интенсификация процесса прямого окисления глюкозы у детей раннего возраста. Причины этого явления.
54. Пентозофосфатный путь превращения глюкозы. Этапы и стадии этого процесса. Окислительная стадия пентозного цикла. Значение пентозофосфатного пути превращения глюкозы для организма.
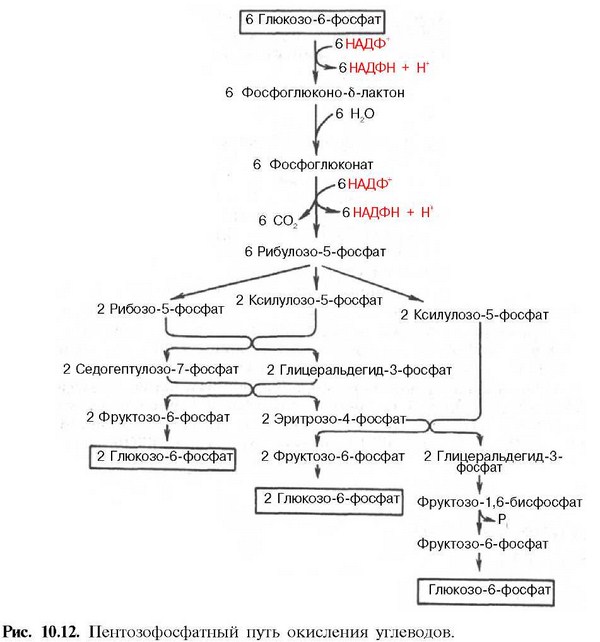
55. Биосинтез глюкозы (глюконеогенез) из неуглеводных продуктов. Основные этапы, три обходных реакции гликолиза. Значение процесса. Гормональная регуляция глюконеогенеза. Взаимосвязь гликолиза в мышцах и глюконеогенеза в печени (цикл Кори).
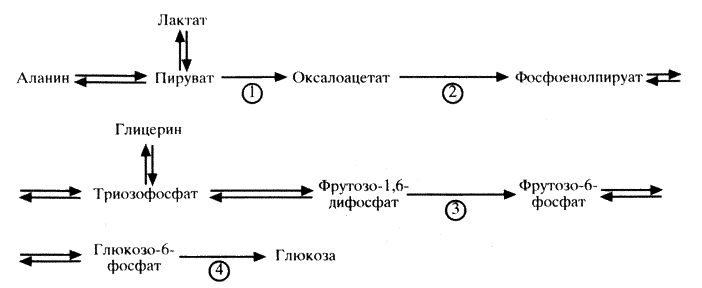
Биосинтез глюкозы (глюконеогенез) из молочной кислоты. Глюконеогенез – обращение реакций гликолиза. Из 11 реакций гликолиза 3 необратимы (1-ая, 3-я и 10-я):
1. глюкоза → глюкозо-6 фосфат
2. фруктозо-6-фосфат → фруктозо-1,6-дифосфат
3. фосфоенолпируват → пируват
Для этих реакций существуют обходные пути. Первый обходной путь начинается в матриксе митохондрий. Пируват проникает сквозь мембрану митохондрий в матрикс митохондрии, затем пируват подвергается карбоксилированию под действием фермента пируваткарбоксилаза (с использованием биотина и энергии АТФ). В результате этой реакции образуется оксалоацетат. Затем оксалоацетат превращается в малат, чтобы выйти из митохондрий в цитозоль клетки для дальнейшего превращения. В цитозоле малат обратимо превращается в оксалоацетат (под действием цитозольной малатдегидрогеназы). Образовавшийся оксалоацетат под действием фермента фосфоенолпируваткарбоксикиназы (при участии биотина и энергии ГТФ) превращается в фосфоенолпируват. В дальнейшем реакции глюконеогенеза идут до образования фруктозо-1,6-дифосфата (обратимые реакции гликолиза). Второй обходной путь: Фруктозо-1,6-дифосфат + Н2О → фруктозо-6-фосфат + Фн (реакцию катализирует фруктозо-1,6-бифосфатаза) Третий обходной путь: Глюкозо-6-фосфат + Н2О → глюкоза + Фн (реакцию катализирует глюкозо-6-фосфатаза) В процессе глюконеогенеза 7-я реакция гликолиза (1,3-дифосфоглицерат + АДФ ↔ 3- фосфоглицерат + АТФ) расходуется 2 АТФ. В ходе синтеза глюкозы используются макроэргические соединения: 4 АТФ и 2 ГТФ. Регуляция глюконеогенеза происходит с участием гормонов и с участием метаболитов.
Гормональная регуляция. Глюкокортикоиды проникают в клетку, усиливают биосинтез ферментов глюконеогенеза, способствуют повышению уровня сахара в крови.
Метаболическая регуляция. Ацетил-КоА стимулирует пируваткарбоксилазу, осуществляющую реакцию превращения пирувата в ЩУК и далее в глюкозу. АТФ участвует в процессах карбоксилирования, стимулирует глюкозо-6-фосфатазу и фруктозо-1,6-бисфосфатазу, усиливает процессы глюконеогенеза. Витамин Н (биотин) – кофермент пируваткарбоксилазы. Фруктозо-2,6- дифосфат ингибирует фруктозо-1,6-бисфосфатазу.
Цикл Кори. В процессе анаэробного гликолиза в скелетных мышцах образуется лактат, который выходит из клеток в кровь, доставляется в печень, где включается в реакции глюконеогенеза с образованием глюкозы. Глюкоза выходит в кровь, доставляется к тканям, где используется как источник энергии. Цикл Кори выполняет две важные функции: 1) обеспечивает утилизацию лактата в реакциях глюконеогеназа; 2) предотвращает накопление лактата и развитие лактоацидоза (смещение рН в кислую сторону)
56. Глюконеогенез у детей. Лабильность этого процесса у ребенка.
57. Свойства и распространение гликогена как резервного полисахарида. Биосинтез гликогена, мобилизация гликогена. Регуляция обмена гликогена гормонами и аллостерическая регуляция. Особенности распада гликогена в печени и мышцах. Гликогенозы и агликогенозы
Гликоген – высокомолекулярный полимер, построенный из остатков глюкозы, связанных α-1,4- и α-1,6-гликозидными связями. В большом количестве содержится в печени и мышцах. Гликоген упакован в гранулы, размер которых 100 – 400 А. В гранулах находятся ферменты синтеза и распада гликогена. Распад гликогена происходит в интервалах между приемами пищи; этот процесс ускоряется при физических нагрузках. Гликогенолиз происходит за счет отщепления остатков глюкозы в виде глюкозо-1-фосфата. Фермент гликогенфосфорилаза расщепляет α-1,4-гликозидные связи в гликогене в результате остаются 4 остатка глюкозы до места ветвления (α-1,6-гликозидная связь). Олигосахаридтрансфераза переносит фрагмент гликогена (состоящий из трех остатков глюкозы) на неразветвленный участок цепи, остается один остаток глюкозы, связанный α-1,6-гликозидной связью. Фермент α-1,6-глюкозидаза отщепляют мономерный остаток глюкозы. Суммарная реакция: Глюкозо-1-фосфат изомеризуется в глюкозо-6-фосфат под действием фосфоглюкомутазы. Образующийся глюкозо-6-фосфат вступает в реакции гликолиза (или пентозного цикла) клеток мышц и мозга, а в клетках печени, почек и кишечника происходит дефосфорилирование глюкозо-6-фосфата с образованием глюкозы и фосфорной кислоты (под действием фермента глюкозо-6-фосфатазы). Далее глюкоза поступает в кровь и разносится с кровью к тканям. Гормональная регуляция обмена гликогена осуществляется адреналином, глюкагоном и инсулином. Адреналин и глюкагон активируют распад гликогена и тормозят его синтез. Гормональная регуляция опосредуется общими механизмами через ц-АМФ, который активирует цитозольные ферменты протеинкиназу, киназу фосфорилазы и киназу гликогенсинтазы. Киназы при участии АТФ фосфорилируют ферменты гликогенфосфорилазу и гликогенсинтазу. Фосфорилаза активна в фосфорилированном состоянии, поэтому под действием адреналина и глюкагона активируется распад гликогена, а гликогенсинтаза, напротив, в фосфорилированном состоянии становится неактивной, поэтому синтез гликогена под действием адреналина и глюкагона тормозится. Под действием инсулина активируются ферменты фосфодиэстераза и фосфопротеинфосфатазы. Фосфодиэстераза разрушает ц-АМФ; фосфопротеинфосфатазы катализируют дефосфорилирование ферментов гликогенфосфорилазы и гликогенсинтазы. Фосфорилаза в дефосфорилированном состоянии становится неактивной и процесс распада гликогена прекращается, а гликогенсинтаза, напротив, активируется, поэтому под действием инсулина активируется синтез гликогена. Заболевания, связанные с обменом гликогена: гликогенозы и агликогенозы. 1. Болезнь Гирке (недостаток глюкозо-6-фосфатазы печени). Клиническая картина: увеличение печени уже в период новорожденности, гипогликемия между приемами пищи, гипогликемия, сохраняющаяся при действии адреналина и глюкагона, нарушение жирового обмена (усиливается отложение жира в подкожной клетчатке и внутренних органах). 2. Болезнь Форбса-Кори (дефицит фермента амило-1,6-глюкозидазы печени): неполное расщепление гликогена, гепатомегалия, мышечная слабость, гипогликемия. Эта болезнь не угрожает жизни ребенка, хотя может привести к задержке физического развития и полового созревания. У некоторых больных в подростковом возрасте отмечается умеренная миопатия, т.к. понижена мышечная утилизация гликогена. 3. Болезнь Андерсена (недостаток активности «ветвящего» фермента в печени): гепатоспленомегалия, цирроз печени, прогрессирующая печеночная недостаточность - длинные неразветвленные цепи плохо разрушаются; развивается цирроз печени. Эта болезнь встречается крайне редко и всегда приводит к ранней смерти. 4. Болезнь Херса (недостаток активности печеночной фосфорилазы): гепатомегалия, умеренная гипергликемия, ацидоз. Данная патология встречается крайне редко и только у мальчиков. 5. Болезнь Мак-Ардля (недостаток мышечной фосфорилазы): спазмы мышц при нагрузке, преходящая миоглобинурия. Агликогенозы – наследственное заболевание, вызванное дефектом гликогенсинтазы. В печени почти или полностью отсутствует гликоген, выражена гипогликемия (0,39 – 0,67 ммоль/л). Характерный симптом – у детей судороги обычно по утрам. Их можно предупредить частым кормлением ночью. Гипогликемия новорожденных. Наблюдается у всех новорожденных из-за прекращения поступления крови через плаценту (преходящая гипогликемия). Коррекция происходит быстро и самостоятельно. У новорожденных, матери которых страдают сахарным диабетом, может быть тяжелая гипогликемия – проявление функционального гиперинсулинизма, устраняемое только введением глюкозы
58. Своеобразие метаболизма гликогена в печени у ребенка. Патология обмена гликогена (гликогенозы).
Биосинтез гликогена. При голодании гликоген распадается до небольшого фрагмента, содержащего 4-6 остатков глюкозы, связанных α-1,4-гликозидными связями. Этот фрагмент гликогена называется «затравкой» (или праймером); на нем начинается процесс биосинтеза гликогена. Глюкоза вступает в синтез гликогена в «активной» форме УДФ-глюкозы. В период пищеварения глюкоза поступает в клетку и фосфорилируется с образованием глюкозо-6-фосфата под действием фермента гексокиназы (или глюкокиназы в печени), затем изомеризуется в глюкозо-1-фосфат под действием фосфоглюкомутазы. Глюкозо-1-фосфат превращается в УДФ-глюкозу под действием фермента УДФ-глюкуронилтрансферазы. Реакция, катализируемая гликогенсинтазой, возможна лишь в том случае, когда в цепи «затравки» гликогена содержится более 4-6 остатков глюкозы:
УДФ-глюкоза + гликоген (n) → гликоген (n+1) + УДФ, где n ≥ 4-6.
УДФ-глюкоза взаимодействует с нередуцированным остатком цепи. Ветвление гликогена осуществляется ферментом амило-1,4-1,6-глюкозилтрансферазой («гликогенветвящий» фермент). Этот фермент присоединяет полисахаридную цепочку (n ≥7) с помощью α-1,6-гликозидной связи (точка ветвления). В месте ветвления находится 10-12 остатков глюкозы, от ближайшего места ветвления не менее 4 остатков глюкозы.
Значение ветвления: увеличивается растворимость, ускоряется рост полисахаридной цепи.
59. Регуляция углеводного обмена: саморегуляция, нейрогуморальная регуляция. Механизм действия глюкозы, адреналина, инсулина и глюкокортикоидов на обмен углеводов. Уровень глюкозы крови в норме. Гипергликемия и гипогликемия. Нарушения обмена углеводов при сахарном диабете. Биохимическая диагностика сахарного диабета
В регуляции постоянства концентрации сахара в крови главную роль выполняет печень. При избыточном поступлении углеводов в организм в печени происходит накопление гликогена, а при недостаточном поступлении, наоборот, гликоген, в ней распадается до глюкозы. Таким образом поддерживается нормальное количество сахара. Постоянство содержания глюкозы в крови, гликогена в печени регулируется нервной системой. На обмен углеводов оказывает влияние кора больших полушарий головного мозга. Доказательством этого является повышение сахара в моче у студентов после трудного экзамена. Центр углеводного обмена находится в гипотоламусе и продолговатом мозге. Влияние гипоталамуса и коры больших полушарий на углеводный обмен осуществляется преимущественно посредством симпатической нервной системы, которая вызывает усиленную секрецию адреналина надпочечниками. Большое значение в углеводном обмене имеют железы внутренней секреции — поджелудочная, щитовидная, надпочечники, гипофиз и др., которые под действием ЦНС регулируют ассимиляцию и диссимиляцию углеводов. Гормон поджелудочной железы инсулин переводит глюкозу в гликоген и тем самым уменьшает количество сахара в крови. Адреналин и гликогон увеличивают расщепление гликогена в печени, в мышцах, вследствие чего увеличивается содержание сахара в крови. Следовательно, инсулин — это сахаропонижающий гормон, гликогон — сахароповышающий. При снижении концентрации сахара в крови возбуждается центр углеводного обмена в гипоталамусе, который дает импульсы поджелудочной железе, и она увеличивает выработку глюкагона до тех пор, пока содержание глюкозы за счет распада гликогена не увеличится до нормального уровня.
60. Лабильность уровня глюкозы в крови у детей раннего возраста. Наклонность к гипогликемии. Своеобразие учета результатов нагрузной пробой сахара у детей.
Нарушение углеводного обмена. Сахарный диабет. Важным показателем гомеостаза является концентрация глюкозы в крови. В норме концентрация глюкозы в крови 3,3 – 5,5 ммоль/л. Больше 5,5 ммоль/л – гипергликемия. В норме сахар в моче отсутствует. Почечный порог для глюкозы – 10 ммоль/л или 180 мг% в крови. Для уточнения диагноза сахарный диабет проводят тест на толерантность к глюкозе. Тест на толерантность к глюкозе (метод сахарной нагрузки). Перед проведением теста в течение 3-х дней рекомендуется низкоуглеводная диета. В день анализа пациенту натощак дают выпить раствор 100 г глюкозы/100 мл воды. Концентрацию глюкозы определяют в трех пробах крови: натощак, через 60 и через 120 минут после «Сахарной нагрузки». Результат на фоне здоровья: натощак менее 5,55 ммоль/л, через 60 мин. – менее 8,88 ммоль/л, через 120 мин. – менее 6,66 ммоль/л. На основе полученных данных строят «сахарные кривые»: Особенности сахарной нагрузки у детей. До 6 лет дают 1,5 г/кг веса, от 6 до 12 – 2 г/кг веса, после 13 лет по 2,5 г/кг веса. Наличие гипергликемии через 2 часа после начала исследования, глюкозурия и обнаружение кетоновых тел в моче – триада симптомов для постановки диагноза сахарный диабет. Содержание глюкозы в моче определяют пробой Бенедикта.
61. Пищевые жиры, их гидролиз. Всасывание продуктов переваривания. Роль желчных пигментов в переваривании и всасывании липидов. Транспорт ресинтезированных в кишечнике жиров. Использование жиров, включенных в транспортные липопротеины: липопротеинлипазы. Гиперлипопротеинемия.
Жиры, или триглицериды — природные органические соединения, полные сложные эфиры глицерина и одноосновных жирных кислот; входят в класс липидов. В живых организмах выполняют структурную, энергетическую и др. функции. Наряду с углеводами и белками, жиры — один из главных компонентов питания. Жидкие жиры растительного происхождения обычно называют маслами. Гидролиз жиров. Расщепление жиров на глицерин и жирные кислоты проводится обработкой их щёлочью — (едким натром), перегретым паром, иногда — минеральными кислотами. Этот процесс называется омылением. Липопротеинлипаза — фермент, относящийся к классу липаз. ЛПЛ расщепляет триглицериды самых крупных по размеру и богатых липидами липопротеинов плазмы крови — хиломикронов и липопротеинов очень низкой плотности (ЛПОНП или ЛОНП)). ЛПЛ регулирует уровень липидов в крови, что определяет её важное значение в атеросклерозе. Гиперлипидемия (гиперлипопротеинемия) — аномально повышенный уровень липидов и/или липопротеинов в крови человека. Нарушение обмена липидов и липопротеинов встречается довольно часто в общей популяции. Гиперлипидемия является важным фактором риска развития сердечно-сосудистых заболеваний в основном в связи со значительным влиянием холестерина на развитие атеросклероза. Кроме этого, некоторые гиперлипидемии влияют на развитие острого панкреатита.
62. Особенности переваривания и всасывания жиров в организме ребенка.
|
|
|
 Скачать 1.95 Mb.
Скачать 1.95 Mb.