клеточный иммунный ответ. Клеточный иммунный ответ. Клеточный иммунный ответ
 Скачать 0.74 Mb. Скачать 0.74 Mb.
|

|
Рис. 8. 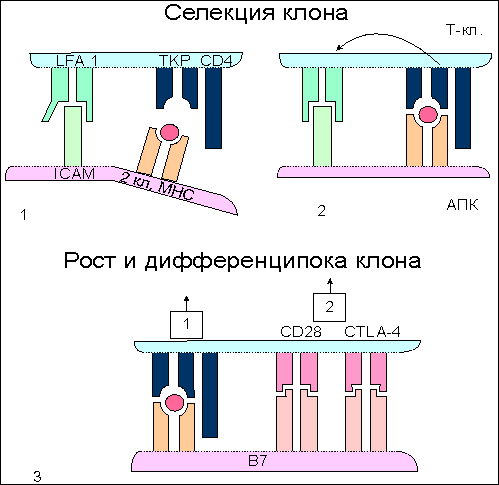 Участие адгезинов и костимуляторов в отборе и дифференцировке наивных Т-клеток. Участие адгезинов и костимуляторов в отборе и дифференцировке наивных Т-клеток. 1. Проникшие в кортикольную зону наивные Т-лимфоциты взаимодействуют неспецифически с антиигенпрезентирующими клетками (АПК) с поиощью адгезинов: LFA-1 - на Т-клетках и ICAM - на АПК. Взаимодействие LFA-1 - ICAM на этом этапе характеризуется низкой аффинностью. Если Т-клеточный антигенраспознающий рецептор (ТКР) "не находит" на АПК соответствующего по специфичности комплекса пептид:молекула МНС (пример генерации Т-хелперов), то клетка покидает АПК и вступает в процесс рециркуляции. 2. При соответствии по специфичности ТКР комплексу пептид:молекула МНС повышается аффинность взаимодействия LFA-1 с ICAM и контактное взаимодействие Т-клетки с АПК становится достаточно прочным. 3. Взаимодействие ТКР с антигенным комплексом формирует при участии ко-рецептора (на рис. ко-рецептор Т-хелперов CD4) первый сигнал к дифференцировке провзаимодействовавших клеток. Второй сигнал формируется ко-стимулятором В7, представленным на поверхности АПК, и соответствующими лигандами на Т-клетках - CD28 и CTLA-4. Только при совместном действии первого и второго сигналов происходит рост и дифференцировка наивных Т-клеток до зрелых эффекторов. Подавляющее большинство наивных Т-клеток не выдерживает отбора на специфичность и покидает лимфатический узел через эфферентный лимфатический сосуд с тем, чтобы вновь вступить в процесс рециркуляции в поисках соответствующих по специфичности антигенов. Связывание антигенспецифического рецептора с комплексом антигенный пептид:молекулы I или II класса МНС и включение в комплексообразование корецепторов CD8 или CD4 обеспечивает лишь одно из условий развития наивных Т-клеток - формирование первого сигнала к пролиферации и дифференцировке этих клеток . Чтобы специфически подготовленная клетка вышла, наконец, в процесс дальнейшего развития, необходим второй сигнал от клеточной поверхности к геному. Костимулятором в данном случае выступает молекула В7 , экспрессирующаяся на мембране антигенпрезентирующей клетки . Рецептором для В7 на поверхности наивной Т-клетки является белок CD28 , относящийся к суперсемейству иммуноглобулинов . Взаимодействие CD28:В7 обеспечивает формирование второго сигнала. Помимо CD28 активированные наивные Т-клетки синтезируют и экспрессируют еще один белок с костимулирующей активностью - СTLA-4 , обладающей большей аффинностью по отношению к В7. Между CD28 и CTLA-4 высокая гомология по последовательности аминокислотных остатков, а кодирующие их гены близко сцеплены в хромосоме. Ясно, что совместное костимулирующее действие этих молекул оказывает больший эффект на развитие Т-клеток. Тот факт, что одна и та же антигенпрезентирующая клетка выполняет двойную функцию - представление антигена в иммуногенной форме и экспрессию лиганда для костимуляции, т.е. определяет формирование специфического и неспецифического сигналов для дифференцировки наивных Т-клеток в активные эффекторы, имеет большой биологический смысл. Отрицательная селекция в тимусе , очевидно, не является абсолютно безошибочным процессом. Определенные "запрещенные" клоны могут выйти в циркуляцию и стать потенциальной причиной аутоиммунного поражения. Однако, как правило, этой аутоагрессии не наблюдается, поскольку сам факт распознавания антигена, в том числе и собственного, не является единственным условием запуска дифференцировки наивных Т-клеток. Необходимо включение костимулятора, которое происходит только на антигенпрезентирующих клетках. Антигенпрезентирующие клетки (АПК) в инициации иммунитета. Макрофаги: роль в инициации клеточного иммунитета. Макрофаги помимо участия в реакциях неспецифического иммунитета проявляют себя и в реакциях специфической иммунной защиты от инфекции в качестве антигенпрезентирующих клеток . В процессе активации T-лимфоцитов , клетки, представляющие антиген в иммуногенной форме на своей поверхности (антигенпрезентирующие клетки), должны обладать, по крайней мере, двумя основными свойствами: - способностью образовывать комплекс антигенного пептида с молекулами I или II классов МНС , что является первым сигналом к пролиферации и дифференцировке наивных T-клеток , и - экспрессировать костимуляторы, обеспечивающие прохождение второго сигнала активации Т-клеток . Макрофаги в состоянии покоя обладают очень незначительным количеством молекул MHC II класса и полностью лишены костимулятора В7 на своей поверхности. Выраженное представительство этих молекул на мембране макрофага начинается после захвата и внутриклеточного переваривания микроорганизмов. Один из способов поглощения бактерий связан с рецепторами к маннозе , которые способны взаимодействовать с углеводами бактериальной стенки. Захваченные микроорганизмы деградируют в фаголизосомах , образуя отдельные пептиды, которые выносятся на клеточную поверхность в комплексе с молекулами MHC. Именно в процессе внутриклеточного переваривания корпускулярого антигена происходит индукция синтеза и экспрессии на клеточной поверхности молекул MHC класса II и костимулятора В7. Факторами индукции, возможно, являются рецепторы клеточной поверхности, взаимодействующие с микроорганизмами, поскольку синтез В7 можно индуцировать простой инкубацией макрофагов с отдельными компонентами (углеводами, липополисахаридами) бактериальной стенки. Индукция костимулирующей активности к общим микробным компонентам позволяет иммунной системе отличать бактериальные антигены от собственных антигенов организма или безвредных, хотя и чужеродных белков. Из практической работы известно, что получение иммунного ответа к некоторым белкам возможно только с использованием адъювантов , включающих убитые микроорганизмы или продукты их бактериальной стенки. Схема возможных отношений в данном случае выглядит следующим образом. Если белковые антигены захватываются и презентируются макрофагами в отсутствие бактериальных компонентов, которые инициируют синтез В7 , то Т-клетка специфически распознает антиген, однако остается рефрактерной, так как отсутствует действие второго сигнала для запуска пролиферации и дифференцировки. Внесение в систему бактериальных компонентов - индукторов костимулятора В7 - обеспечивает полноценное включение в иммунный ответ Т-клеток. В условиях эксперимента аутоиммунное заболевание легко индуцируется смесью собственных тканевых антигенов с компонентами бактериальной стенки, иллюстрируя тем самым значение костимуляции в процессе разграничения "своего" от "чужого". Понимание того факта, что запуск Т-клеточного ответа связан с двухсигнальной системой активации, внесло ясность в работу макрофагов в качестве "мусорщиков". Купферовские клетки печени и макрофаги селезенки постоянно захватывают и разрушают отжившие клетки этих органов. При этом в отсутствие бактериальных стимуляторов экспрессируемые на поверхности фагоцитирующих клеток собственные антигены как результат деградации захваченных отживших клеток не в состоянии развить аутоиммунный ответ. В представленных примерах иммуногенность связана не со структурными особенностями антигена, а с реактивностью организма, с потенциальными возможностями его иммунокомпетентных клеток. Дендритные клетки: роль в инициации иммунитета. Не все патогены индуцируют синтез и экспрессию костимуляторов на поверхности макрофагов . К этой категории возбудителей инфекционнного заболевания относятся, в частности, вирусы, которые в течение эволюции адаптировались к использованию биосинтетического аппарата клетки для своего воспроизведения. Кажется, что подобная эксплуатация клеток противоречит принципам "всеобщей" защищенности организма от чужеродных антигенов. Подобное несоответствие разрешается включением в противовирусную защиту дендритных клеток . Этот тип антигенпрезентирующих клеток обильно представлен в лимфоидной ткани и обладает выраженной и, что очень важно, постоянной экспрессией костимулятора В7 , молекул I и II классов МНС , а также адгезинов ICAM-1 , ICAM-3 , LFA-3 . Иначе, для дендритных клеток не требуется инициации поверхностных структур, принимающих участие в формировании Т-клеточного ответа . Подобная заданность иммунологически значимых молекул определяет защитный потенциал дендритных клеток. При этом они не обладают способностью к фагоцитозу , но легко усваивают белки и вирусные частицы посредством пиноцитоза . Экспонирование антигена B-клеткам происходит на фолликулярных дендритных клетках (ФДК). Моноциты , активированные in vitro гранулоцитарно-макрофагальным колониестимулирующим фактором и интерлейкином-4 , теряют способность к фагоцитозу и превращаются в АПК, приобретая морфологию дендритных клеток, и начинают экспрессировать белки MHC класса II. Постоянная экспрессия молекул I и II классов MHC у этого типа клеток приводит к быстрому формированию иммуногенных комплексов вирусных пептидов с соответствующими молекулами. В результате создаются условия для включения в ответ наивных CD8 Т-клеток и наивных CD4 Т-клеток , дифференцирующихся в ТН1-клетки воспаления и хелперные ТН2-клетки . Большинство вирусных белков локализуется в цитозоле клетки, где они разрушаются до отдельных пептидов в протеасомах ( рис. 9 ). Особенность локализации вирусных белков обеспечивает формирование комплекса c молекулами I класса MHC в эндоплазматическом ретикулуме и его последующий прямой транспорт к плазматической мембране. Образование комплекса с некоторыми поверхностными вирусными белками имеет иной характер. Такие белки, оказавшись в эндоплазматических вакуолях , которые содержат молекулы II класса MHC , транспортируются к клеточной поверхности в составе этих вакуолей ( рис. 10 ). Комплекс вирусный пептид:молекулы II класса будет фактором включения в ответ Т-хелперов. Таким образом, в целом, дендритные клетки играют основную роль в формировании и клеточного, и гуморального антивирусного иммунитета. Рис. 9. 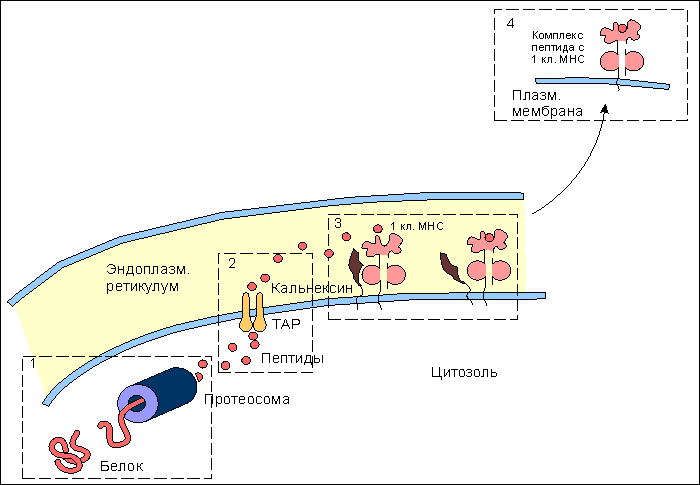 Этапы подготовки вирусных белков к взаимодействию с молекулами I класса главного комплекса гистосовместимости. I этап - разрушение вирусных белков, находящихся в цитозоле, с помощью протеазного комплекса - протеосомы. II этап - транспорт образовавшихся пептидов во внутреннее пространство эндоплазматического ретикулума с помощью ТАР-1 и ТАР-2, образующих гетеродимер на эндоплазматической мембране. III этап - встреча транспортируемых пептидов с молекулами I класса МНС. Молекулы МНС стабилизированы специальным белком - калнексином. Взаимодействие пептида с молекулой I класса приводит к отсоединению калнексина. Образовавшийся комплекс пептид:молекула I класса готов к дальнейшему транспорту к плазматической мембране (ПМ). IV этап - завершающий. Комплекс через аппарат Гольджи транспортируется к клеточной поверхности. Таким образом, вирусный пептид в комплексе с молекулой I класса МНС становится доступным (иммуногенным) для его распознавания Т-клеточными антигенраспознающими рецепторами. Рис. 10. 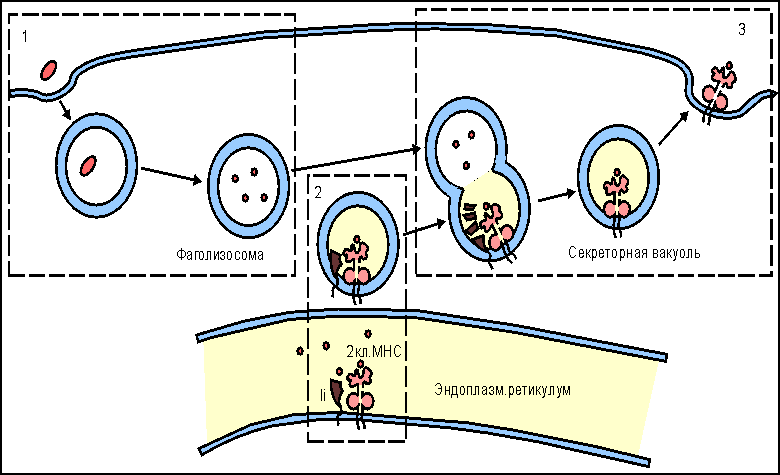 Этапы подготовки антигенов бактерий и их токсинов к взаимодействию с молекулами II класса главного комплекса гистосовместимости. I этап - поглощение бактерий или их токсинов фагоцитирующей, способной к презентации антигена клеткой и разрушение захваченного материала до отдельных пептидов в фаголизосомах. II этап - во внутреннем пространстве эндоплазматического ретикулума происходит сборка молекул II класса, которые до встречи с пептидом комплексированы со специальным белком, получившим название инвариантной цепи (Ii). Этот белок защищает молекулу II класса от случайной встречи с бактериальными пептидами в эндоплазматическом ретикулуме. Комплекс молекулы II класса с Ii покидает эндоплазматический ретикулум в составе вакуоли. III этап - вакуоль, содержащая комплекс молекулы II класса с Ii, сливается с фаголизосомой. Кислые протеазы фаголизосом разрушают Ii белок и таким образом снимают запрет на взаимодействие молекул II класса с бактериальными пептидами. Образовавшийся новый комплекс пептид:молекула II класса в составе секреторной вакуоли перемещается к мембране клетки. Результатом всех этих процессов является экспрессия чужеродного пептида в комплексе с молекулой II класса на клеточной поверхности, что и обеспечивает доступность бактериального пептида для антигенраспознающих рецепторов Т-клеток. Клетки Лангерганса: роль в инициации иммунитета Особое место в формировании иммунного ответа принадлежит клеткам Лангерганса . До недавнего времени эти клетки относили к тканевым макрофагам кожи. В настоящее время считается, что этот тип клеток функционально и морфологически следует относить к предшественникам дендритных клеток . При проникновении патогена через поврежденный участок кожи среди прочих клеточных форм в реакцию изоляции микроорганизма вступают клетки Лангерганса, способные к фагоцитозу , но не экспрессирующие костимулятор В7 . Мигрируя по лимфатическим сосудам в ближайший лимфатический узел клетки оседают и трансформируются в типичные дендритные клетки с поверхностным корецептором В7, что создает условия для включения в иммунный ответ CD8 T-клеток и CD4 T-клеток . Таким образом, функция этих клеток состоит в доставке антигена в регионарную лимфоидную ткань, последующей его обработке до иммуногенной формы и инициации как клеточного, так и гуморального иммунного ответа. B-лимфоциты: роль в инициации клеточного иммунитета. Помимо макрофагов и дендритных клеток еще одним типом клеток, способным представлять антиген в иммуногенной форме для наивных T-клеток , являются В-лимфоциты . Если макрофаги поглощают в основном бактерии, а дендритные клетки - различные вирусы, то объектом активности В-клеток являются белковые антигены , включая бактериальные токсины. Потенциальную способность В-лимфоцитов выступать в качестве антигенпрезентирующих клеток в T-клеточном иммунном ответе определяют два основных свойства B-клеток: - наличие поверхностных специфических иммуноглобулиновых рецепторов (sIg) и - выраженная экспрессия молекул II класса МНС . При этом у покоящихся В-клеток отсутствует третий обязательный компонент клеточной мембраны антигенпрезентирующих клеток - костимулятор В7 , однако он начинает экспрессироваться под влиянием компонентов бактериальных стенок, таких, например, как полисахариды. Активация В-клеток начинается после взаимодействия поверхностных иммуноглобулинов с белковым антигеном (11 ). Рис. 11. 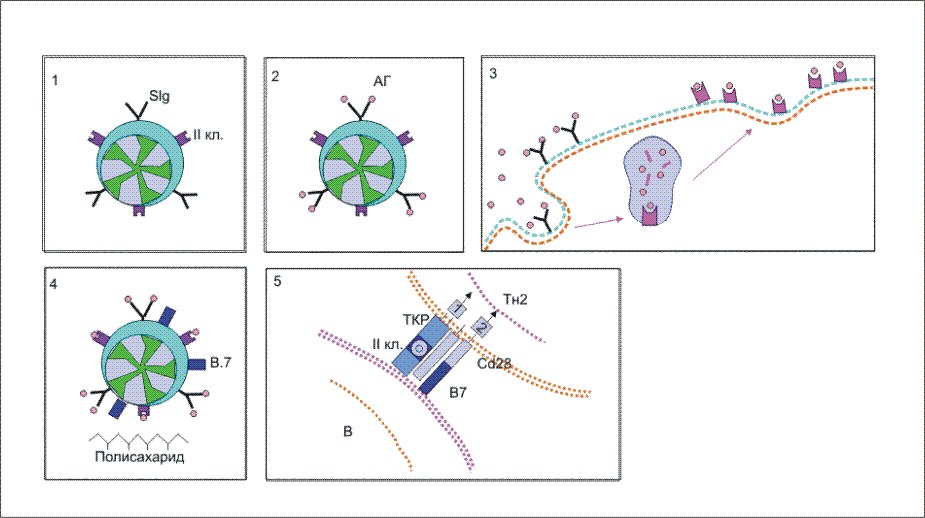 В-лимфоцит как антигенпрезентирующая клетка. 1. Выраженная экспрессия поверхностного рецепторного иммуноглобулина (sIg) и молекул II класса МНС у В-клеток. 2. Взаимодействие sIg с антигеном (АГ). Обычными для В-клеток антигенами являются токсины бактерий. 3. Эндоцитоз В-клеткой комплекса антиген:sIg и презентация антигенного пептида с молекулами II класса МНС на клеточной поверхности. 4. Экспрессия ко-стимулятора В7 на поверхности В-клетки под влиянием бактериальных полисахаридов. 5. Заключительный этап процесса - формирование двухсигнальной системы активации Т-хелперов (ТH2) при их взаимодействии с В-клетками. Первый сигнал (1) образуется от взаимодействия ТКР ТН с комплексом антигенный пептид:молекула II класса МНС. Второй сигнал (2) развивается при взаимодействии индуцированного ко-стимулятора В7 с CD28 на ТН2. В результате эндоцитоза образовавшегося комплекса и его деградации в лизосомальных вакуолях начинается экспрессия пептидных фрагментов белка в ассоциации с молекулами II класса МНС. Активный синтез этих молекул В-клетками обеспечивает выраженное представительство комплекса пептид:молекулы II класса на поверхности клетки. При этом в работу не включаются хелперные T-клетки , поскольку отсутствует экспрессия костимулятора В7 . В условиях реального ответа к инфекционным агентам стимулятором синтеза В7 выступают компоненты клеточной стенки бактерий. Как только начинается экспрессия В7, специфически провзаимодействовавшие наивные Т-клетки вступают в процесс пролиферации и дифференцировки, образуя активные хелперные T-клетки (ТН2) . Включение в работу зрелых Т-хелперов создает условия для полноценного развития гуморального иммунного ответа. Как и в случае с макрофагами, необходимость двойного сигнала для активации хелперных T-клеток является условием, контролирующим ответ В-клеток к собственным антигенам. При отсутствии инфекции специфическое взаимодействие наивной Т-клетки с В-лимфоцитом, экспрессирующим аутоантиген, приводит к анергии или гибели соответствующего клона Т-клеток. АПК в инициации иммунитета: заключительный обзор Т-клеточный ответ примируется тремя типами антигенпрезентирующих клеток (АПК) ( табл. 3 ), каждый из которых адаптирован к обработке определенного класса антигенов : - макрофагами , захватывающими и перерабатывающими до иммуногенной формы в основном бактерии и другие корпускулярные антигены; - дендритными клетками , которые поглощают самые разнообразные вирусные частицы, и - В-клетками , активность которых направлена на белковые антигены, включая бактериальные токсины. Табл. 3 Основные характеристики антигенпрезентирующих клеток
|