Лекция 1. Клиническая анатомия и физиология уха
 Скачать 1.14 Mb. Скачать 1.14 Mb.
|

|
Рис. 5.22. Инерционный (а) и компрессионный (б) механизмы костного проведения Рассмотрим роль отдельных элементов органа слуха в проведении звуковых волн. Ушная раковина играет роль своеобразного коллектора, направляющего высокочастотные звуковые колебания во вход в наружный слуховой проход. Ушные раковины имеют также определенное значение в вертикальной ототопике. При изменении положения ушных раковин вертикальная ототопика искажается, а при выключении их путем введения в наружные слуховые проходы полых трубочек полностью исчезает. Однако при этом не нарушается способность локализовать источники звука по горизонтали.
Наружный слуховой проход является проводником звуковых волн к барабанной перепонке. Ширина и форма наружного слухового прохода не играют особой роли при звукопроведении. Однако полное заращение просвета наружного слухового прохода или его обтурация препятствуют распространению звуковых волн и приводят к заметному ухудшению слуха. В слуховом проходе вблизи барабанной перепонки поддерживается постоянный уровень температуры и влажности независимо от колебаний температуры и влажности во внешней среде, и это обеспечивает стабильность упругих свойств барабанной перепонки. Кроме того, в наружном слуховом проходе происходит избирательное усиление на 10-12 дБ звуковых волн частотой около 3 кГц. С физической точки зрения это объясняется резонансными свойствами слухового прохода, имеющего длину около 2,7 см, что составляет 1/4 длины волн резонансной частоты. ПОЛОСТЬ СРЕДНЕГО УХА И СЛУХОВАЯ ТРУБА Для нормального функционирования системы звукопроведения необходимо, чтобы по обе стороны барабанной перепонки было одинаковое давление. При несоответствии давления в полостях среднего уха и в наружном слуховом проходе натяжение барабанной перепонки меняется, акустическое (звуковое) сопротивление возрастает и слух понижается. Выравнивание давления обеспечивается вентиляционной функцией слуховой трубы. При глотании или зевании слуховая труба открывается и становится проходимой для воздуха. Учитывая, что слизистая оболочка среднего уха постепенно поглощает воздух, нарушение вентиляционной функции слуховой трубы ведет к повышению наружного давления над давлением в среднем ухе, что вызывает втяжение барабанной перепонки внутрь. Это при- водит к нарушению звукопроведения и вызывает патологические изменения в среднем ухе.
Помимо вентиляционной, слуховая труба выполняет также защитную и дренажную функции. Защитная функция слуховой трубы обеспечивается слизистой оболочкой, которая в хрящевом отделе особенно богата слизистыми железами. Секрет этих желез содержит лизоцим, лактоферин, иммуноглобулины - все эти факторы препятствуют проникновению возбудителей в барабанную полость. Дренажную функцию слуховая труба выполняет благодаря наличию мерцательного эпителия, движения ресничек которого направлены в сторону глоточного устья трубы. Барабанная перепонка и слуховые косточки. По законам физики, передача звуковых волн из воздуха в жидкие среды внутреннего уха сопровождается потерей до 99,9% звуковой энергии. Это связано с различным акустическим сопротивлением указанных сред. Структуры среднего уха - барабанная перепонка и рычажная система слуховых косточек - являются тем механизмом, который компенсирует потерю акустической (звуковой) энергии при переходе из воздушной среды в жидкую. Благодаря тому, что площадь основания стремени (3,2 мм2) в окне преддверия значительно меньше рабочей 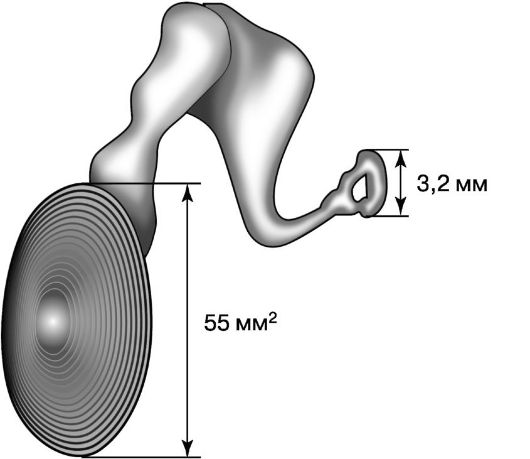 Рис. 5.23. Влияние соотношения площадей барабанной перепонки и основания стремени на увеличение силы звука Рис. 5.23. Влияние соотношения площадей барабанной перепонки и основания стремени на увеличение силы звукаплощади барабанной перепонки (55 мм2), увеличивается сила звуковых колебаний за счет уменьшения амплитуды волн (рис. 5.23). Увеличение силы звука происходит также в результате рычажного способа сочленения слуховых косточек. В целом давление на поверхности окна преддверия оказывается примерно в 19 раз больше, чем на барабанной перепонке. Благодаря барабанной перепонке и слуховым косточкам воздушные колебания большой амплитуды и малой силы трансформируются в колебания перилимфы с относительно малой амплитудой, но большим давлением. Слуховые мышцы. В барабанной полости расположены две самые миниатюрные мышцы человеческого тела: напрягающая барабанную перепонку и стременная. Первая из них иннервируется тройничным нервом, вторая - лицевым, и это определяет различие в раздражителях, вызывающих сокращение той и другой мышцы, и их неодинаковую роль. Обеспечивая оптимальное натяжение отдельных элементов звукопроводящего аппарата, эти мышцы регулируют передачу звуков разной частоты и интенсивности, и тем самым выполняют аккомодационную функцию. Защитная функция внутриушных мышц обеспечивается тем, что при воздействии звуков большой мощности мышцы рефлекторно резко сокращаются. Это в конечном счете приводит к уменьшению звукового давления, передаваемого перилимфе.
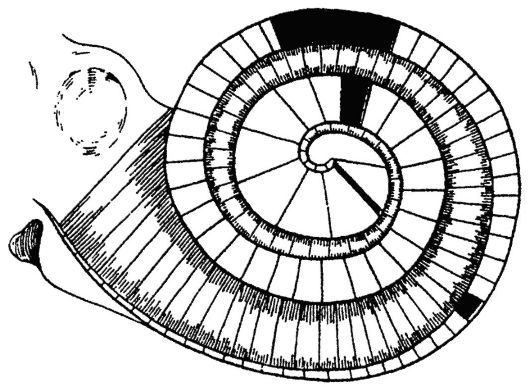 Рис. 5.24. Схема резонансной теории слуха Гельмгольца Рис. 5.24. Схема резонансной теории слуха ГельмгольцаЭтим рецепторы внутреннего уха предохраняются от сильных звуков. Звуковосприятие представляет сложный нейрофизиологический процесс трансформации энергии звуковых колебаний в нервный импульс, его проведение до центров в коре головного мозга, анализ и осмысливание звуков. Звуковая волна, дошедшая через окно преддверия до перилимфы, вовлекает ее в колебательные движения. Эти колебания восходят по завиткам улитки, по лестнице преддверия к ее вершине, где через геликотрему переходят на барабанную лестницу, по которой возвращаются к основанию улитки, вызывая прогиб вторичной барабанной перепонки. В колебания вовлекается базилярная мембрана и находящийся на ней спиральный орган, чувствительные волосковые клетки которого при этих колебаниях подвергаются сдавлению или натяжению покровной (текториальной) мембраной. Упругая деформация волосковых клеток лежит в основе их раздражения, что означает трансформацию механических звуковых колебаний в электрические нервные импульсы. Для объяснения происходящих во внутреннем ухе процессов рецепции звуков предложены различные теории слуха. Пространственная (или резонансная) теория была предложена Гельмгольцем еще в 1863 году и основана на представлениях о периферическом анализе звука на уровне улитки. Теория допускает, что базилярная мембрана состоит из серии сегментов, каждый из которых резонирует в ответ на воздействие определенной частоты звукового сигнала. Входящий стимул, таким образом, приводит к вибрации тех участков базилярной мембраны, собственные частотные характеристики которых соответствуют компонентам стимула. По аналогии со струнными инструментами звуки высокой частоты приводят в колебательное движение (резонируют) участок базилярной мембраны с короткими волокнами у основания улитки, а звуки низкой частоты вызывают колебания участка мембраны с длинными волокнами у верхушки улитки (рис. 5.24).
Согласно резонансной теории, любой чистый тон имеет свой ограниченный участок восприятия на базилярной мембране. При подаче и восприятии сложных звуков одновременно начинает колебаться несколько участков мембраны. Теория Гельмгольцавпервыепозволилаобъяснитьосновные свойства уха - способность определения высоты, громкости и тембра. В свое время эта теория нашла много сторонников и до сих пор считается классической. Вывод Гельмгольца о том, что в улитке происходит первичный анализ звуков, нашел подтверждение в работах Л.А. Андреева. Согласно его данным, при разрушении верхушки улитки у собак наблюдается выпадение условных рефлексов на низкие звуки, при разрушении ее основного завитка - на высокие звуки. Резонансная теория Гельмгольца получила подтверждение и в клинике. Гистологическое исследование улиток умерших людей, страдавших понижением слуха, позволило обнаружить изменения спирального органа в участках, соответствующих утраченной части слуха. Вместе с тем современные знания не подтверждают возможность резонирования отдельных «струн» базилярной мембраны. Вслед за теорией Гельмгольца появилось множество других пространственных теорий. Особый интерес представляет теория «бегущей волны» лауреата Нобелевской премии Бекеши (1960). Прямое изучение механических свойств базилярной мембраны показало, что ей не свойственна высокая механическая избирательность. Звуковые волны различных частот вызывают колебания мембраны на довольно больших ее участках. Звуки определенной высоты вызывают на базилярной мембране «бегущую волну», гребню которой соот- 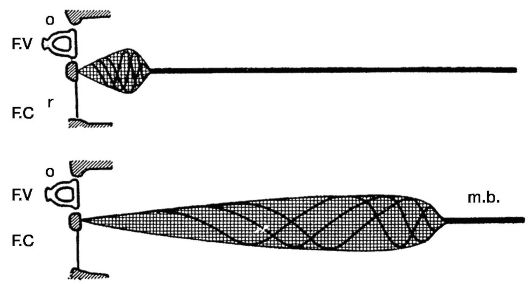 Рис. 5.25. Схема теории «бегущей волны» Бекеши: Рис. 5.25. Схема теории «бегущей волны» Бекеши:а - при высоких звуках; б - при низких звуках; FV - окно преддверия; FC - окно улитки; m.b. - базилярная пластинка
ветствует наибольшее смещение мембраны на одном из ее участков. Локализация этого участка зависит от частоты звуковых колебаний. Наиболее низкие звуки вызывают прогибание мембраны у верхушки улитки, звуки высокой частоты - в области основного завитка улитки (рис. 5.25). Базилярная мембрана больше всего смещается на гребне «бегущей волны» и, колеблясь, вызывает деформацию сдвига волосковых клеток спирального органа над этим участком мембраны. Отрицательным моментом этой теории является то, что с механической точки зрения невозможно объяснить способность различать ухом огромное множество разных частот. По мнению П.П. Лазарева, при механическом раздражении волосковых клеток в них возникает химическая реакция, сила которой зависит от количества разлагающегося вещества (слухового пурпура); при этом освобождаются ионы, которые и вызывают процесс нервного возбуждения. Гуморальная регуляция функции кортиева органа в определенной степени обеспечивается особыми клетками сосудистой полоски - апудоцитами, являющимися элементами системы эндокринной клеточной регуляции. Апудоциты продуцируют биогенные амины - серотонин, мелатонин и пептидные гормоны - адреналин, норадреналин. Функция подкорковых слуховых центров изучена недостаточно. Через них осуществляется безусловная рефлекторная связь с двигательными реакциями в ответ на воздействие звука: повороты головы, глаз, кохлео-пальпебральный рефлекс Бехтерева, кохлео-пупиллярный рефлекс Шурыгина и т.п. Роль корковых отделов слухового анализатора заключается в осуществлении высшего анализа звуковых сигналов и синтеза их в слитный звуковой образ. Корковый отдел не только принимает и анализирует информацию, поступающую от кохлеарных рецепторов, но и имеет эфферентную связь с улиткой, через посредство которой кора регулирует, настраивает функциональную активность рецепторного аппарата. С деятельностью центральных отделов в коре височной доли связаны такие свойства слухового анализатора, как ототопика, адаптация, маскировка и др.
ФУНКЦИЯ ВЕСТИБУЛЯРНОГО АНАЛИЗАТОРА Вестибулярная функция зависит от деятельности вестибулярных рецепторов, расположенных в ампулах полукружных каналов и мешочках преддверия. Это интерорецепторы, воспринимающие информацию о положении тела или головы в пространстве, изме- нении скорости и направления движения. Полный и тонкий анализ полученной от вестибулярных рецепторов информации осуществляется, как и в отношении звуковых сигналов, при участии всего анализатора, включая его центральные отделы. Трансформационным механизмом, преобразующим механическую энергию в нервный импульс, является смещение волосков нейроэпителиальных клеток с помощью инерционных структур: в мешочках преддверия - отолитовой мембраны, в полукружных каналах - эндолимфы и купулы. Под влиянием смещения этих инерционных структур происходит упругая деформация пространственно поляризованного волоскового аппарата рецепторных клеток ампулярного и отолитового отделов. Как известно, в волокнах вестибулярного нерва в состоянии покоя регистрируется постоянная биоэлектрическая активность. При воздействии на вестибулярные рецепторы адекватного раздражителя с положительным или отрицательным значением происходит возрастание или уменьшение импульсации по сравнению с исходным, в состоянии покоя, уровнем. Объяснением этому может быть тот факт, что сгибание чувствительных волосков под влиянием смещения эндолимфы (в ампуле) или отолитовой мембраны (в мешочках преддверия) приводит к изменению взаимной ориентации киноцилии и стереоцилий, расстояние между которыми либо уменьшается, либо увеличивается. Это, в свою очередь, сопровождается гиперили гипополяризацией клеток и в конечном счете - торможением или возбуждением рецепторных клеток (рис. 5.26). Адекватным раздражителем для ампулярных рецепторов является угловое ускорение с положительным или отрицательным знаком. Система полукружных каналов осуществляет анализ кругового ускоренного движения и в физиологических пределах наиболее приспособлена к реагированию на повороты головы. Отолитовые рецепторы реагируют на действие прямолинейного ускорения и постоянно регистрируют направление земного притяжения по отношению к голове. Отолитовый аппарат наиболее приспособлен к реагированию в физиологических условиях на наклоны головы, запрокидывание головы, начало и конец ходьбы, спуск и подъем.
В соответствии с рассмотренными ранее ассоциативными связями вестибулярного анализатора различают вестибулярные реакции, 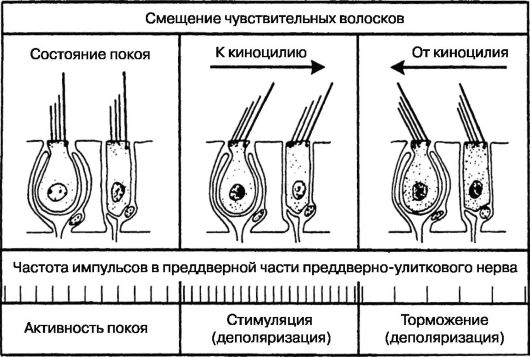 Рис. 5.26. Изменение спонтанной активности вестибулярных рецепторов в зависимости от направления смещения киноцилия Рис. 5.26. Изменение спонтанной активности вестибулярных рецепторов в зависимости от направления смещения киноцилиякоторые по природе своей могут быть сенсорные, вегетативные или соматические. Все вестибулогенные реакции являются системными реакциями организма и могут быть физиологическими или патологическими. *Вестибулосенсорные реакции обусловлены наличием вестибулокортикальных связей и проявляются осознанием положения и изменения положения головы в пространстве. Патологической спонтанной вестибулосенсорной реакцией является головокружение. *Вестибуловегетативные реакции связаны с тесным взаимодействием ядерного вестибулярного комплекса и ретикулярной фармации. Вестибулярные влияния на висцеральные органы опосредованы через симпатические и парасимпатические отделы нервной системы. Они имеют адаптационный характер и могут проявляться изменением самых разнообразных жизненных функций: возрастанием артериального давления, учащением сердцебиения, изменением дыхательного ритма, возникновением тошноты и даже рвоты при воздействии вестибулярного раздражения. *Вестибулосоматические (анимальные) реакции обусловлены связями вестибулярных структур с мозжечком, поперечно-полосатой мускулатурой конечностей, туловища и шеи, а также с глазодвигательной мускулатурой. Соответственно различают вестибуломозжечковые, вестибулоспинальные и вестибулоглазодвигательные реакции. *Вестибуломозжечковые реакции направлены на поддержание положения тела в пространстве посредством перераспределения мышечного тонуса в динамическом состоянии организма, т.е. в момент совершения активных движений на фоне воздействия ускорений.
*Вестибулоспинальные реакции связаны с влиянием вестибулярной импульсации на мышечный тонус шеи, туловища и конечностей. При этом возрастание импульсации от вестибулярных рецепторов одного из лабиринтов приводит к повышению тонуса поперечно-полосатой мускулатуры противоположной стороны, одновременно ослабляется тонус мышц на стороне возбужденного лабиринта. *Вестибулоглазодвигательные (окуломоторные) реакции обусловлены связями вестибулярной системы с ядрами глазодвигательных нервов. Эти связи делают возможными рефлекторные сочетанные отклонения глаз, в результате которых направление взгляда не меняется при перемене положения головы. Они же определяют возникновение нистагма. Способность человека сохранять вертикальное положение тела в покое и при движении, обозначаемая как функция равновесия, может быть реализована лишь при содружественном функционировании ряда систем, среди которых важную роль играет вестибулярный анализатор. Наряду с другими сенсорными системами, зрительной и проприоцептивной, вестибулярный аппарат участвует в информационном обеспечении и реализации функции равновесия. Информация о положении тела в пространстве от различных сенсорных входов поступает в центральные отделы вестибулярного анализатора, экстрапирамидной системы, мозжечок, ретикулярную фармацию и кору головного мозга. Здесь осуществляется интеграция поступающей информации и переработка поступающих сигналов для воздействия на эффекторные органы (рис. 5.27).  |