Колебания и волны. Звук. Ультразвук. Колебания. Гармонические колебания. Характеристики колебаний амплитуда, период, частота, циклическая частота, фаза
 Скачать 1.35 Mb. Скачать 1.35 Mb.
|

|
63. Механика мышечного сокращения. Саркомеры. Взаимодействие актиновых и миозиновых нитей. Строение мышечных волокон. Сократимость – способность мышцы укорачиваться при возбуждении, в результате чего возникает сила тяги.Скелетная мышца состоит из мышечных волокон (миоцитов). Мышечные волокна представляют собой гигантские многоядерные клетки. Волокно покрыто эластичной оболочкой — сарколеммой и состоит из саркоплазмы, содержащей митохондрии, рибосомы, трубочки и пузырьки саркоплазматической сети и так называемая Т-система. Т-система также имеет прямое отношение к мышечному сокращению, так как по ней передаётся изменение электрического потенциала поверхностной мембраны элементам ретикулума, что приводит к освобождению ионов кальция, поступающих к миофибриллам и запускающих процесс мышечного сокращения. Сократительные элементы – 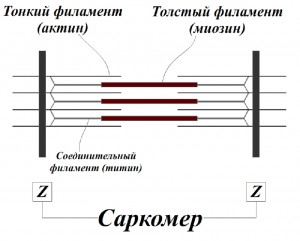 миофибриллы. миофибриллы. Они состоят из многочисленных параллельно расположенных нитей – филаментов. Перегородки, называемые Z – пластинками, разделяют их на отдельные участки, называемые саркомерами. Филаменты бывают двух типов: толстые (из белка миозина) и тонкие (из актина). В поперечном сечении толстые и тонкие нити располагаются правильными шестиугольниками так, что каждая толстая нить окружена шестью тонкими, а каждая тонкая нить может вступать в контакт с тремя толстыми. Мышца сокращается в результате укорочения множества последовательно соединенных саркомеров в миофибриллах, при этом тонкие актиновые нити скользят вдоль толстых миозиновых, двигаясь между ними к середине саркомера. Единственным прямым источником энергии для мышечного сокращения служит АТФ. При активации мышцы повышение внутриклеточной концентрации ионов кальция приводит к сокращению и усиленному расщеплению АТФ, интенсивность метаболизма повышается в 100-1000 раз. АТФ гидролитически расщепляется с помощью миозин-АТФ-фазы до АДФ и неорганического фосфата. Расщепление одного моля АТФ обеспечивает около 48 кДж энергии.40-50% этой энергии преобразуется в механическую работу, а 50-60% превращается в тепло. 64. КПД мышечных сокращений. 40% если считать АТФ готовым продуктом, 20% если учесть затраты энергии на синтез АТФ. Нерв.импульс--высвобождение ионов Са—наползание нитей КПД мышечной клетки около 50 %, мышцы в целом не более 20%. Максимальная сила мышц не достигается в реальных условиях; не все клетки мышцы используются одновременно и сокращаются с максимальной силой, иначе при сокращении многих скелетных мышц будут повреждены сухожилия или кости (что иногда и наблюдается при сильных судорогах). КПД мышцы также зависит от внешних условий; например, на холоде он значительно снижается, так как для организма важнее сохранить температуру тела. 65. Изотонический режим работы мышц. Уравнение Хилла. Изометрический режим. Статическая работа мышц. Различают два вида работы мышц: статическая (звенья ОДА фиксированы, движение отсутствует); динамическая (звенья ОДА перемещаются относительно друг друга). Различают три режима мышечного сокращения: изометрический – режим мышечного сокращения, при котором момент силы мышцы равен моменту внешней силы (длина мышцы не изменяется). Изометрический режим соответствует статической работе. преодолевающий (концентрический) – режим мышечного сокращения, при котороммомент силы мышцы больше момента внешней силы (длина мышцы уменьшается). уступающий (эксцентрический) – режим мышечного сокращения, при котором момент силы мышцы меньше момента внешней силы (длина мышцы увеличивается). v – скорость укорочения;P – сила (нагрузка);P0 – максимальное изометрическое напряжение, которое может развивать мышца;b – константа, имеющая размерность скорости; a – константа, имеющая размерность силы Механическая работа (А), совершаемая мышцей, измеряется произведением поднимаемого веса (Р) на расстояние (h): А = Р * h Чем больше величина груза, тем меньше высота, на которую его поднимает мышца. Изотонический режим – сокращение мышцы при неизменном напряжении, выражающееся в уменьшении её длины и увеличении поперечного сечения. В организме изотоническое мышечное сокращение в чистом виде не наблюдается. К чисто изотоническому мышечному сокращению приближается движение ненагруженной конечности. Изометрический режим (режим постоянной длины мышцы) характеризуется напряжением мышцы в условиях, когда она закреплена с обоих концов или когда мышца не может поднять слишком большой груз. При этом h = 0 и, соответственно, механическая работа тоже равна нулю (А = 0). Этот режим наблюдается при сохранении заданной позы и при выполнении статической работы . В этом случае в мышечном волокне все равно происходят процессы возникновения и разрушения мостиков между актином и миозином, т. е. тратится энергия на эти процессы, но отсутствует механическая реакция перемещения нитей актина вдоль миозина. 66. Второй закон механики Ньютона. Его применение для анализа травматизма. Способы увеличения продолжительности удара. Изменение количества движения пропорционально приложенной движущей силе и происходит по направлению той прямой, по которой эта сила действует. a=F/m При травмаопасных ситуациях тело испытывает большое отрицательное ускорение. По второму закону Ньютона, ускорение – результат действия силы: a=F/m => ∆V/∆t=F/m => F∆t=m∆V 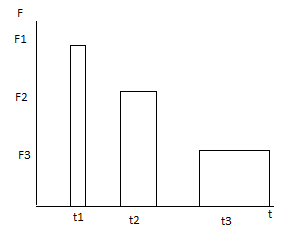 Одно и то же значение m∆V можно обеспечить разными способами. Способы уменьшения силы удара – это способы увеличения его продолжительности:
67. Строение и модели клеточных мембран. Биологические мембраны образуют наружную оболочку всех животных клеток и формируют многочисленные внутриклеточные органеллы. Толщина мембраны несколько нанометров. Мембраны в основном состоят из липидов и белков, количество которых неодинаково у разных типов клеток. Сложность изучения молекулярных механизмов функционирования клеточных мембран обусловлена тем, что при выделении и очистке клеточных мембран нарушается их нормальное функционирование. В настоящее существует несколько моделей клеточной мембраны, среди которых наибольшее распространение получила жидкостно-мозаичная модель: мембрана представлена бислоем фосфолипидных молекул, ориентированных таким образом, что гидрофобные концы молекул находятся внутри бислоя, а гидрофильные головки направлены в водную фазу. Такая структура идеально подходит для образования раздела двух фаз: вне- и внутриклеточной. Эта модель демонстрирует структуру размещения протеинов внутри или на биполярном слое липидов. Размещаются протеины в хаотичном порядке, при этом получается мозаика протеинов. За счет протеинов осуществляется проницаемость, активный транспорт через мембрану, генерация электрического потенциала. Мембраны выполняют две функции: матричную – являются основой для удержания белков и барьерную – защищают клетку от проникновения чужеродных частиц. 68. Физические свойства биологических мембран. С одной стороны мембрана ведет себя как жидкость, с другой стороны, мембрана – это упорядоченная структура. Следовательно, фосфолипиды в мембране находятся в жидкокристаллическом состоянии. Вязкость липидного слоя больше вязкости воды, примерно равна вязкости масла. Поверхностное натяжение меньше, чем у воды. При измнении температуры в мембране происходят фазовые переходы: плавление липидов при нагревании и кристаллизация при охлаждении. Жидкокристаллическое состояние имеет меньшую вязкость и большую растворимость, чем твердое состояние. Толщина жидкокристаллического слоя меньше, чем твердого. Структура молекул в жидком и твердом состоянии различна. В жидкой фазе молекулы фосфофлипидов могут образовывать полоски, в которые способны внедряться молекулы диффундирующего вещества. Двойной фосфолипидный слой уподобляет мембрану конденсатору, электроемкость 1 мм2 состовляет 5-13 нФ. 69. Функции клеточной или плазматической мембраны Мембрана обеспечивает: 1) Избирательное проникновение в клетку и из нее молекул и ионов, необходимых для выполнения специфических функций клеток; 2) Избирательный транспорт ионов через мембрану, поддерживая трансмембранную разницу электрического потенциала; 3) Специфику межклеточных контактов. Благодаря наличию в мембране многочисленных рецепторов, воспринимающих химические сигналы — гормоны, медиаторы и другие биологически активные вещества, она способна изменять метаболическую активность клетки. Мембраны обеспечивают специфику иммунных проявлений, благодаря наличию на них антигенов — структур, вызывающих образование антител, способных специфически связываться с этими антигенами. Ядро и органеллы клетки также отделены от цитоплазмы мембранами, которые предупреждают свободное движение воды и растворенных в ней веществ из цитоплазмы в них и наоборот. Это создает условия для разделения биохимических процессов, протекающих в различных отсеках (компартментах) внутри клетки. 70. Ионный состав цитоплазмы и межклеточной жидкости. Проницаемость клеточной мембраны для различных ионов. Разность потенциалов на мембране клетки. Поскольку плазма и межклеточная жидкость разделены высокопроницаемой мембраной капилляров, ионный состав этих жидкостей почти одинаков. Однако, проницаемость клеточной мембраны для различных ионов различна. Концентрация положительно заряженных ионов (катионов) в плазме немного выше (примерно на 2%) по сравнению с межклеточной жидкостью. Белки плазмы в сумме заряжены отрицательно, следовательно, они способствуют присоединению катионов, таких как ионы Na+ и К+, удерживая их в плазме в несколько большем количестве. Напротив, в межклеточной жидкости отрицательно заряженные ионы (анионы) присутствуют в слегка повышенной по сравнению с плазмой концентрации, т.к. отрицательный заряд белков плазмы отталкивает анионы. Внеклеточная жидкость содержит большое количество ионов Na+ и Cl- и достаточно высокое число бикарбонатов; содержание же калия, кальция, магния, фосфатов и ионов органических кислот в ней незначительно. Внутриклеточная жидкость, в отличие от внеклеточной, содержит лишь небольшое количество ионов натрия и хлора, а ионы кальция в ней практически отсутствуют. Внутри клетки, напротив, содержится очень большое количество ионов калия, а также умеренное число ионов магния и сульфатов; концентрация всех перечисленных веществ вне клетки низка. Кроме того, в клетках содержится большое количество белка, в 4 раза превышающее его содержание в плазме. Мембранный потенциал — разность электрических потенциалов между наружной и внутренней поверхностями биологической мембраны, обусловленная неодинаковой концентрацией ионов. Внутренняя поверхность заряжена отрицательно по отношению к наружной. Чем больше разность потенциалов между сторонами мембраны, тем больше ток. В состоянии покоя мембрана клетки мало проницаема для ионов Na°. При возбуждении проницаемость мембраны увеличивается, и она пропускает положительно заряженные ионы натрия внутрь клетки, что приводит к понижению мембранной разности потенциалов (деполяризации мембраны) и даже к появлению разности потенциалов противоположного знака. 71. Потенциал покоя клетки. Уравнение Гольдмана-Ходжкина-Катца. Потенциал покоя — мембранный потенциал возбудимой клетки в невозбужденном состоянии. Он представляет собой разность электрических потенциалов, имеющихся на внутренней и наружной сторонах мембраны и составляет у теплокровных от -60 до -100 мВ. -60мВ – порог возбудимости. Условия возникновения потенциала покоя: Наличие градиентов концентрации ионов на цитомембране. Различная проницаемость мембраны для различных ионов. Мембрана возбудимой клетки способна быстро менять свою проницаемость для определённого вида катионов, вызывая переход положительного заряда с внешней стороны на внутреннюю. Уравнение Гольдмана-Ходжина-Катца. ЭДС=(RT/Fz)*ln*(pK +[Ko]+pNa+[Nao]+…+pA - [Ai-])/(pK+[Ki]+pNa+[Nai]+…+pA – [Ao-]) R - универсальная газовая постоянная, Т - абсолютная температура растворов, F - число Фарадея, p - проницаемость мембраны, [] – концентрация иона, i – внутри клетки, о – снаружи, А – анион. Итак, мембранный потенциал покоя - это дефицит положительных электрических зарядов внутри клетки, возникающий за счёт утечки из неё положительных ионов калия и электрогенного действия натрий-калиевого насоса. 72. Возбудимость клеток и тканей. Методы возбуждения. Закон «всё или ничего». Нервные и мышечные ткани обладают возбудимостью, т.е. способны в ответ на внешнее раздражение резко изменять свое состояние. Не всякое по интенсивности раздражение приводит к возбуждению. Минимальная сила раздражения, вызывающая возбуждение, носит название порога раздражения. Чем больше минимальная сила раздражения, требуемая для вызова реакций, чем выше порог раздражения, тем ниже возбудимость. По отношению к разным раздражителям порог раздражения может быть различен.Обязательным признаком возбуждения является изменение электрического состояния поверхностной клеточной мембраны. В возбудимой клетке постоянно имеется разность электрических потенциалов между ее цитоплазмой и внешней средой, Внутренняя поверхность заряжена отрицательно по отношению к наружной. Эту разность потенциалов называют мембранным потенциалом. Потенциал мембраны может изменяться под действием различных стимулов. Искусственным стимулом может служить электрический ток, подаваемый на внешнюю или внутреннюю сторону мембраны через электрод. В естественных условиях стимулом часто служит химический сигнал от соседних клеток, поступающий через синапс или путём диффузной передачи через межклеточную среду. Закон «все или ничего». Если раздражение ниже пороговой силы, то клетка никак не отреагирует. Если больше, то реакция клетки будет стандартной. В результате надпорогового возбуждения на мембране возбудимой клетки генерируется электрический импульс, который заканчивается восстановлением прежнего состояния. 73. Потенциал действия: графический вид и характеристики, механизмы возникновения и развития. Потенциал действия — волна возбуждения, перемещающаяся по мембране живой клетки. Представляет собой электрический разряд — быстрое кратковременное изменение потенциала на небольшом участке мембраны возбудимой клетки (нейрона, мышечного волокна или железистой клетки), в результате которого наружная поверхность этого участка становится отрицательно заряженной по отношению к соседним участкам мембраны, тогда как его внутренняя поверхность становится положительно заряженной по отношению к соседним участкам мембраны. Потенциал действия является физической основой нервного или мышечного импульса. В основе любого потенциала действия лежат следующие явления:1. Мембрана живой клетки поляризована 2. Мембрана обладает избирательной проницаемостью 3. Мембрана возбудимой клетки способна быстро менять свою проницаемость для определённого вида катионов, вызывая переход положительного заряда с внешней стороны на внутреннюю. Потенциал мембраны может изменяться под действием различных стимулов. Искусственным стимулом может служить электрический ток, подаваемый на внешнюю или внутреннюю сторону мембраны через электрод. В естественных условиях стимулом часто служит химический сигнал от соседних клеток, поступающий через синапс или путём диффузной передачи через межклеточную среду. График потенциала действия 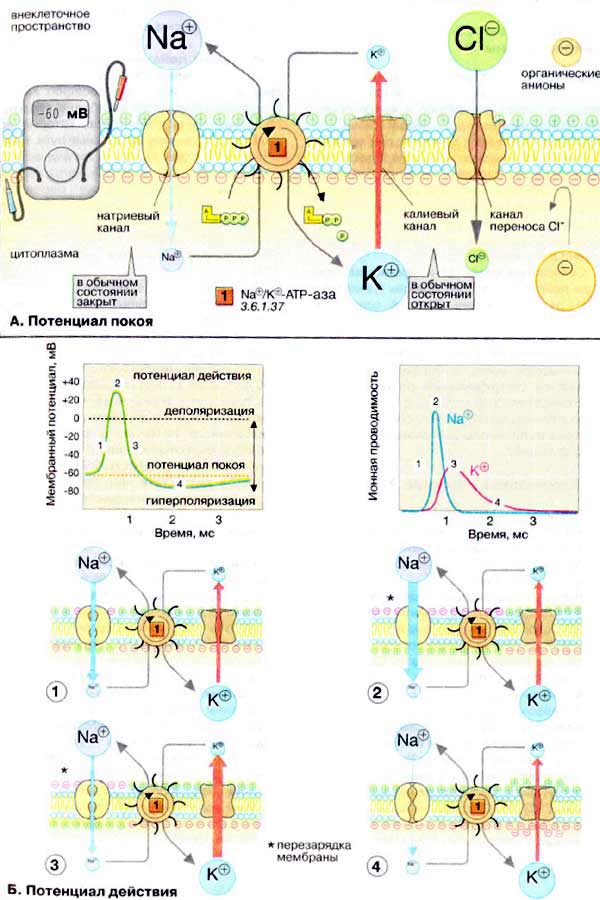 Потенциал действия возникает в ткани под влиянием порогового и сверхпорогового раздражителей и является импульсивным возбуждением. Под влиянием раздражителей изменяется проницаемость клеточной мембраны - повышается для всех потенциалобразующих ионов, но больше всего для ионов Nа+ (в 500 раз). В результате движение ионов Na происходит внутрь клетки, ионы К двигаются из клетки, что приводит к перезарядке клеточной мембраны. Наружная поверхность мембраны несет отрицательный заряд, внутренняя – положительный. Ионы Na путем простой диффузии поступают в клетку без затрат энергии. Достигнув пороговой силы, мембранный потенциал снижается до критического уровня деполяризации (примерно 50 мВ). Критический уровень деполяризации – это то количество милливольт, на которое должен снизиться мембранный потенциал, чтобы возник лавинообразный ход ионов Na в клетку. Пик потенциала действия является постоянным компонентом потенциала действия. Он состоит из двух фаз: фазы деполяризации; фазы реполяризации. Лавинообразное поступление ионов Na в клетку приводит к изменению потенциала на клеточной мембране. Чем больше ионов Na войдет в клетку, тем в большей степени деполяризуется мембрана, тем больше откроется активационных ворот. Возникновение заряда с противоположным знаком называется инверсией потенциала мембраны. Движение ионов Na внутрь клетки продолжается до момента электрохимического равновесия по иону Na. Амплитуда потенциала действия не зависит от силы раздражителя, она зависит от концентрации ионов Na и от степени проницаемости мембраны к ионам Na. Нисходящая фаза (фаза реполяризации) возвращает заряд мембраны к исходному знаку. При достижении электрохимического равновесия по ионам Na происходит инактивация активационных ворот, снижается проницаемость к ионам Na и возрастает проницаемость к ионам K. Полного восстановления мембранного потенциала не происходит. В процессе восстановительных реакций на клеточной мембране регистрируются следовые потенциалы – положительный и отрицательный. |