биология. EKZAMEN_BIO (Восстановлен). Краткий обзор 1) единство химического состава, 2) обмен веществ, 3) самовоспроизведение (репродукция), 4) наследственность
 Скачать 1.8 Mb. Скачать 1.8 Mb.
|

|
Вопрос 5. Назовите мембранные органоиды эукариотической клетки. Краткий ответ. Органоиды (от греч. органон - орудие, орган и идос - вид, подобие) - это надмолекулярные структуры цитоплазмы, выполняющие специфичные функции, без которых невозможна нормальная деятельность клетки. По своей структуре органоиды подразделяют на немембранные (не содержащие мембранных компонентов) и мембранные (имеющие мембраны) . Мембранные органоиды (эндоплазматическая сеть, комплекс Гольджи, лизосомы, пероксисомы, митохондрии и пластиды) характерны только для эукариотических клеток. Развернутый ответ. Эндоплазматическая сеть(ЭПС), или эндоплазматическая редикула (ЭПР) -это сеть каналов, пронизывающая цитоплазму. Стенки этих каналов представляют собой мембраны, контактирующие со всеми органами клетки.ЭПС и органоиды вместе составляют единую внутриклеточную систему, которая осуществляет обмен веществ и энергии в клетки обеспечивает внутриклеточный транспорт веществ. Различают гладкую и гранулярную ЭПС. Гранулярная ЭПС состоит из мембранных мешочков (цистерн), покрытых рибосомами, благодаря чему она кажется шероховатой (шероховатая ЭПС). ЭПС может быть и лишена рибосом (гладкая ЭПС); ее строение ближе к трубчатому типу. На рибосомах гранулярной сети синтезируются белки, которые затем поступают внутрь каналов ЭПС, где и приобретают третичную структуру. На мембранах гладкой ЭПС синтезируются липиды и углеводы, которые также поступают внутрь каналов ЭПС. ЭПС выполняет следующие функции: участвует : участвует в синтезе органических веществ, транспортирует синтезированные вещества в аппарат Гольджи, разделяет клетку на отсеки. Кроме того, в клетках печени ЭПС участвует в обезжиривание ядовитых веществ, а в мышечных клетках играет роль депо кальция, необходимого для мышечного сокращения. Аппарат Гольджи (Golgi Apparatus)-совокупность мембранных цистерн, пузырьков, трубочек, мешочков в клетке, обычно соединенная с эндоплазматическим ретикулумом. В нем накапливаются, упаковываются и в дальнейшем транспортируются белки, производимые в эндоплазматическом ретикулуме. (Синтезированные продукты выводятся из клетки с помощью элементов комплекса Гольджи. Кроме того, в них происходит синтез полисахаридов и образование белково-углеводных комплексов - ред.) Аппарат Гольджи очень хорошо развит в клетках, секретирующих различные вещества (например, в клетках поджелудочной железы, которые вырабатывают пищеварительные ферменты) Лизосомы (гр. лизо - растворять, сома - тело)- это небольшие пузырьки диаметром порядка 1 мкм, ограниченные мембраной и содержащие комплекс ферментов, который обеспечивает расщепление жиров, углеводов и белков. Они участвуют в переваривании частиц, попавших в клетку в результате эндоцитоза и в удалении отмирающих органов (например хвоста у головастиков), клеток и органоидов.При голодании лизосомы растворяют некоторые органоиды, не убивая при этом клетку. Образование лизосом идет в комплексе Гольджи. Митохондрии(гр. митос - нить и хондрион - гранула) - внутриклеточные органоиды, оболочка которых состоит из двух мембран. Наружная мембрана - гладкая, внутренняя образует выросты, называемые кристами. Внутри митохондрии находится полуждиткий матрикс, который сдержит РНК, ДНК, белки, липиды, углеводы, ферменты, АТФ и тр. вещества; в матриксе имеются также рибосомы. Размер митохондрий от 0,2- 0,4 до 1-7 мкм. Количество зависит от вида клетки ( например в клетке печени может быть 1000-2500 митохондрий). Митохондрии могут быть спиральными, округлыми, вытянутыми, чашевидными и т.д.; митохондрии могут менять форму.Функции митохондрий связаны с тем, что на внутренней мембране находятся дыхательные ферменты и ферменты синтеза АТФ. Благодаря этому митохондрии обеспечивают клеточное дыхание и синтез АТФ. Митохондрии могут сами синтезировать белки, т.к. в них есть собственные ДНК, РНК и рибосом. По своему строению митохондрии напоминают клетки прокариот; в связи с этим предполагают, что они произошли от внутриклеточных аэробных симбиотов. Митохондрии имеются в цитоплазме клеток болшенства растений и животных. Пластиды — органоиды эукариотических растений, прокариотов и некоторых фотосинтезирующих простейших. Покрыты двойной мембраной и имеют в своём составе множество копий кольцевой ДНК. Совокупность пластид клетки образует пластидом. По окраске и выполняемой функции выделяют три основных типа пластид(совокупность П. всех типов носит название пластом или пластидом. ): Лейкопласты — неокрашенные пластиды, как правило выполняют запасающую функцию. В лейкопластах клубней картофеля накапливается крахмал. Лейкопласты высших растений могут превращаться в хлоропласты или хромопласты. Хромопласты — пластиды, окрашенные в жёлтый, красный, зеленый или оранжевый цвет. Окраска хромопластов связана с накоплением в них каротиноидов. Хромопласты определяют окраску осенних листьев, лепестков цветов, корнеплодов, созревших плодов. Хлоропласты — пластиды, несущие фотосинтезирующие пигменты — хлорофиллы. Имеют зелёную окраску у высших растений, харовых и зелёных водорослей. Набор пигменто в, участвующих в фотосинтезе (и, соответственно, определяющих окраску хлоропласта) различен у представителей разных таксономических отделов. Хлоропласты имеют сложную внутреннюю структуру. Внутриклеточные органеллы цитоплазмы автотрофных растений, содержащие пигменты и осуществляющие синтез органических веществ. Вопрос 6. Назовите немембранные органоиды эукариотической клетки Краткий обзор: Рибосомы, клеточный центр и органоиды движения (жгутики и реснички). Полный ответ: РИБОСОМА, органоид, синтезирующий белки. Представляет собой сферическую частицу диаметром около 20 нм, состоящую из двух субчастиц, которые могут разъединяться и вновь объединяться. Структурный каркас рибосомы образован молекулами рибосомальной РНК (р-РНК) и связанными с ними белками. В клетках эукариот рибосомы формируются в ядрышке, где на ДНК синтезируется р-РНК, к которой затем присоединяются белки. Субчастицы рибосомы выходят из ядра в цитоплазму, и здесь завершается формирование полноценных рибосом. В цитоплазме рибосомы свободно находятся в цитоплазматическом матриксе или прикрепляются к внешним мембранам ядра и эндоплазматической сети. Свободные рибосомы синтезируют белки для внутренних нужд клетки. Рибосомы на мембранах образуют комплексы – полирибосомы, которые синтезируют белки, поступающие через эндоплазматическую сеть в аппарат Гольджи и затем секретируемые клеткой. В хлоропластах и митохондриях есть свои собственные мелкие рибосомы, они обеспечивают этим органоидам автономный (независимый от ядра) биосинтез белков Клеточный центр — органоид большинства клеток животных, , расположенный (в интерфазе) в центре клетки вблизи ядра. Клеточный центр состоит из двух центриолей и центросферы. Каждая центриоль имеет вид цилиндра, стенки которого образованы девятью триплетами микротрубочек. Центриоли расположены перпендикулярно друг другу и окружены радиально расходящимися от них микротрубочками, образующими лучистую центросферу. Перед делением клетки центриоли удваиваются и попарно расходятся к противоположным полюсам клетки. Функцией их является образование веретена деления (белковых микротрубочек), обеспечивающего равноценное распределение генетического материала между дочерними клетками. Реснички и жгутики - это выступающие из клетки органеллы, имеющие диаметр около 0,25 мкм и содержащие в середине пучок параллельно расположенных микротрубочек. Главная функция этих органелл состоит в передвижении самих клеток или в продвижении вдоль клеток окружающей их жидкости и частиц. Жгутики отличаются от ресничек лишь длиной. Обычно реснички короче жгутиков более чем в 10 раз. На всю длину реснички или жгутика тянутся микротрубочки - полые белковые цилиндры с внешним диаметром 25 нм. Микротрубочки, как и микрофиламенты, полярны, они удлиняются с одного конца вследствие полимеризации глобулярного белка. В ресничках и жгутиках они располагаются по системе 9+2; девять двойных микротрубочек (дублетов) образуют стенку цилиндра, в центре которого находятся две одиночные микротрубочки. Дублеты способны скользить друг относительно друга, что заставляет ресничку или жгутик изгибаться. Вопрос 7. Шероховатая эндоплазматическая сеть. Краткий обзор: Эндоплазматическая сеть — это обязательная органелла эукариотической клетки. Она обнаружена в клетках растений, животных и человека. Функции этой составляющей части клетки разнообразны и связаны в основном с синтезом, модификацией и транспортом органических соединений. Основная часть: Впервые эндоплазматическая сеть была обнаружена в 1945 году. Американский ученый К. Портер разглядел ее с помощью одного из первых электрических микроскопов. С этого времени началось ее активное исследование. В клетке есть две разновидности этой органеллы: Гранулярная, или шероховатая эндоплазматическая сеть (покрыта множеством рибосом). Агранулярная, или гладкая эндоплазматическая сеть. СТРОЕНИЕ. Это сеть каналов, трубочек, замкнутых полостей, которые образованы мембраной. Шероховатая эндоплазматическая сеть — содержит на своей поверхности рибосомы, т.е. на ней идет синтез белка. Функции. синтез белков, предназначенных для выведения из клетки ("на экспорт"); отделение (сегрегация) синтезированного продукта от гиалоплазмы, то есть выведение продукта обмена веществ из жидкой части цитоплазмы. конденсация (переход вещества из газообразного состояния вещества в жидкое) и модификация(изменение строения их радикалов и структуры, направленные на улучшение функциональных свойств, при помощи химических или ферментативных реакций) синтезированного белка; транспорт синтезированных продуктов в цистерны пластинчатого комплекса(мембранные органеллы эукариотической клетки) или непосредственно из клетки; синтез билипидных (с двойным слоем липидов) мембран. Вопрос №8 Гладкая эндоплазматическая сеть. Краткий обзор: ЭПС- одномембранная органелла,представляет собой совокупность мембранных вакуолей, трубочек и плоских мешков (цистерн), распределённых тем или иным способом в цитоплазме. Основная часть: Гладкая (агранулярная) ЭПС - цистерн почти нет, с мембранами вакуолей и трубочек никакие видимые структуры не связаны. Функции: В мембраны гладкой ЭПС встроены ферменты гид- роксилирования (или микросомального окисления). Они необходимы - при синтезе многих липидов (например, стероидных гормонов) - и для обезвреживания токсических соединений. Гладкая ЭПС особенно выражена в клетках - надпочечников и гонад, синтезирующих стероидные гормоны, - и печени – в связи с синтезом холестерина. В 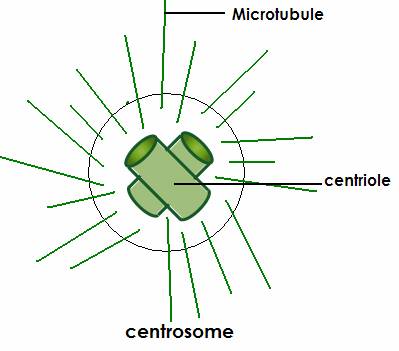 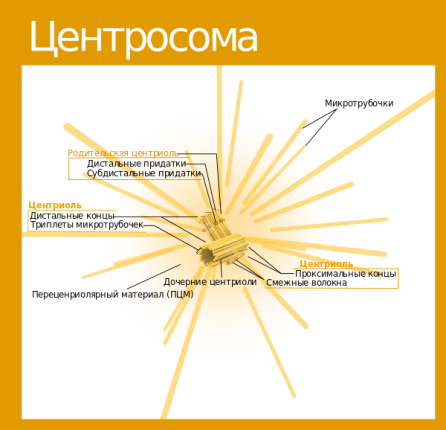 опрос 9. опрос 9.Центросома. Веретено деления. Молекулярное строение и функция. Краткий обзор: Клеточный центр (центросома)— немембранный органоид, главный центр организации микротрубочек и регулятор хода клеточного цикла в клетках эукариот. Веретено́ деле́ния — динамичная структура, которая образуется в митозе и мейозе для обеспечения сегрегации хромосом и деления клетки. Полный ответ: Центриоли, обычно расположенные в паре (диплосома), и окруженные зоной более светлой цитоплазмы, от которой радиально отходят тонкие фибриллы (центросфера). Совокупность центриолей и центросферы называют клеточным центром. Совокупность центриолей и центросомы называют клеточным центром. К 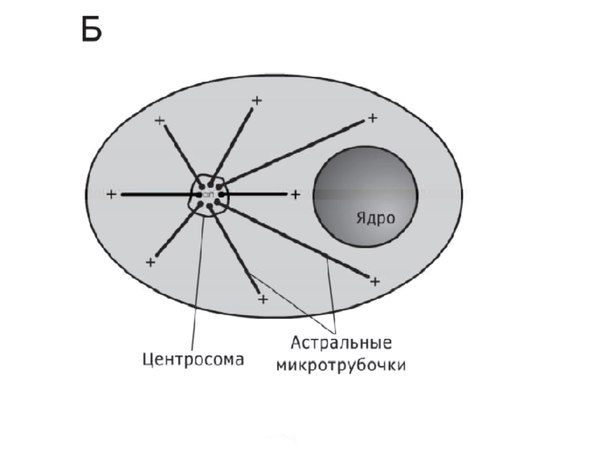 леточный центр (центросома)— немембранный органоид, главный центр организации микротрубочек и регулятор хода клеточного цикла в клетках эукариот. Важная роль его заключается в клеточном делении, в частности, в образовании митотического веретена. Впервые обнаружена в 1883 году Теодором Бовери, который назвал её «особым органом клеточного деления. В клетке содержится одна или две центросомы. леточный центр (центросома)— немембранный органоид, главный центр организации микротрубочек и регулятор хода клеточного цикла в клетках эукариот. Важная роль его заключается в клеточном делении, в частности, в образовании митотического веретена. Впервые обнаружена в 1883 году Теодором Бовери, который назвал её «особым органом клеточного деления. В клетке содержится одна или две центросомы.  Модель центриоли. Изображены девять триплетов микротрубочек. В центросоме центриоли расположены под прямым углом друг к другу. В ходе фазы S клеточного цикла центриоли дуплицируются. При этом образовавашиеся дочерние центриоли располагаются перпендикулярно по отношению к материнским. В митозе пары центриолей, каждая из которых состоит из первоначальной и вновь образованной, расходятся к полюсам клетки и участвуют в образовании митотического веретена. К  аждая центриоль построена из 27 цилиндрических элементов (микротрубочек), сгруппированных в 9 триплетов образованных в результате полимеризации белка тубулина. Девять триплетов микротрубочек расположены по окружности, образуя таким образом полый цилиндр. аждая центриоль построена из 27 цилиндрических элементов (микротрубочек), сгруппированных в 9 триплетов образованных в результате полимеризации белка тубулина. Девять триплетов микротрубочек расположены по окружности, образуя таким образом полый цилиндр. В интерфазе клеточного цикла центросомы ассоциированы с ядерной мембраной. В профазе митоза ядерная мембрана разрушается, центросома делится, и продукты её деления (дочерние центросомы) мигрируют к полюсам делящегося ядра. Микротрубочки, растущие из дочерних центросом, крепятся другим концом к так называемым кинетохорам на центромерах хромосом, формируя веретено деления. По завершении деления в каждой из дочерних клеток оказывается только по одной центросоме. Помимо участия в делении ядра, центросома играет важную роль в формировании жгутиков и ресничек. Центриоли, расположенные в ней, выполняют функцию центров организации для микротрубочек аксонем жгутиков. У организмов, лишенных центриолей (например, у сумчатых ), жгутики не развиваются. Веретено́ деле́ния — динамичная структура, которая образуется в митозе и мейозе для обеспечения сегрегации хромосом и деления клетки. Т 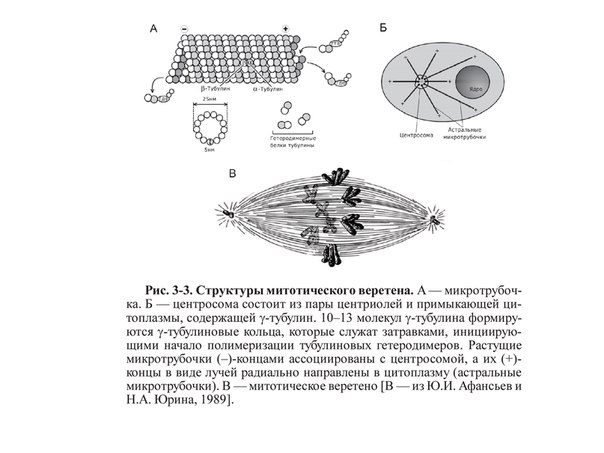 ипичное веретено является биполярным — между двумя полюсами образуется веретенообразная система микротрубочек. Микротрубочки веретена присоединяются к кинетохорам хроматид в области центромер и обеспечивают движение хромосом по направлению к полюсам. ипичное веретено является биполярным — между двумя полюсами образуется веретенообразная система микротрубочек. Микротрубочки веретена присоединяются к кинетохорам хроматид в области центромер и обеспечивают движение хромосом по направлению к полюсам.Веретено образуют три основных структурных элемента: микротрубочки, полюса деления и хромосомы. В организации полюсов деления у животных участвуют центросомы, содержащие центриоли. Важную роль в формировании веретена играют моторные белки, относящиеся к семействам динеинов и кинезинов. Полноценное веретено деления образуется на стадии прометафазы после разрушения ядерной мембраны. Веретено деления типичной клетки млекопитающих состоит из трёх структурных элементов — центросом, микротрубочек и хромосом, — которые образуют симметричную биполярную структуру. На полюсах веретена располагаются центросомы — небольшие органеллы, функционирующие как центры организации микротрубочек. Каждая центросома состоит из пары центриолей, окруженных множеством разных белков. Между полюсами веретена находятся конденсированные хромосомы, состоящие из пары хроматид, скреплённых в области центромеры. На цетромерных участках хромосом находятся кинетохоры — сложные структуры, отвечающие за прикрепление к микротрубочкам веретена[1]. Веретено деления состоит из двух полуверетён. Полуверетено образуется из поляризованных микротрубочек. Отрицательные минус-концы микротрубочек собираются на полюсах веретена вокруг центросом. Плюс-концы микротрубочек отдаляются от двух полюсов и пересекаются в средней экваториальной части веретена. У большинства позвоночных полуверетено состоит из 600—750 микротрубочек, 30—40 % которых заканчиваются на кинетохорах. Микротрубочки, которые соединяют полюса веретена с кинетохорами хромосом, называютсякинетохорными. Причём каждый кинетохор при образовании веретена связывается с множеством микротрубочек и образует кинетохорный пучок. Микротрубочки, которые располагаются между полюсами и не присоединяются к кинетохорам, называютсямежполюсными. Часть микротрубочек веретена образует вокруг каждого полюса радиальные структуры, называемые звёздами или астерами. Такие микротрубочки называются астральными. Вопрос №10 Митохондрии Краткий обзор (из методички) – Двумембранные образование, имеющие собственную ДНК, предположительно возникли из прокариот после объединение с эукариотическими клетками в результате эволюции и последующего с ними сосуществования (симбиоза). Обеспечивают синтез АТФ за счет реакций окислительного фосфорилирования. Митохондрии контролируют внутриклеточное содержание ионов кальция, обеспечивают образование тепла, а также запрограммированной гибели клеток. Основная часть (из инета) - Это органеллы размером с бактерию. Митохондрия ограничена двумя мембранами - гладкой внешней и складчатой внутренней, имеющей очень большую поверхность. Складки внутренней мембраны глубоко входят в матрикс митохондрий, образуя поперечный перегородки - кристы. Пространство между внешней и внутренней мембранами обычно называют межмембранным пространством. Мембраны митохондрий содержат интегральные мембранные белки. Во внешнюю мембрану входят порины, которые образуют поры и делают мембраны проницаемыми для веществ с молекулярной массой до 10 кДа. Внутренняя же мембрана митохондрий непроницаема для большинства молекул; исключение составляют О2, СО2, Н20. Внутренняя мембрана митохондрий характеризуется необычно высоким содержанием белков (75%).В их число входят транспортные белки-переносчики, ферменты, компоненты дыхательной цепи и АТФ-синтаза. Кроме того, в ней содержится необычный фосфолипид кардиолипин. Матрикс также обогащен белками, особенно ферментами цитратного цикла. Метаболические функции Митохондрии являются «силовой станцией» клетки, поскольку за счет окислительной деградации питательных веществ в них синтезируется большая часть необходимого клетке АТФ (АТР). В митохондриях локализованы следующие метаболические процессы: превращение пирувата в ацетил-КоА, катализируемое пируватдегидрогеназным комплексом: цитратный цикл; дыхательная цепь, сопряженная с синтезом АТФ (сочетание этих процессов носит название «окислительное фосфорилирование»); расщепление жирных кислот путем β-окисления и частично цикл мочевины. Митохондрии также поставляют клетке продукты промежуточного метаболизма и действуют наряду с ЭР как депо ионов кальция, которое с помощью ионных насосов поддерживает концентрацию Са2+ в цитоплазме на постоянном низком уровне (ниже 1 мкмоль/л). Главной функцией митохондрий является захват богатых энергией субстратов (жирные кислоты, пируват, углеродный скелет аминокислот) из цитоплазмы и их окислительное расщепление с образованием СО2 и Н2О, сопряженное с синтезом АТФ. Вопрос 11. Рибосомы. Полирибосомы. Митохондриальные рибосомы. Краткий обзор: Рибосома- немембранный двухсубъкдиничный (малая и большая субъединицы) органоид, состоящий из рРНК и белков. Рибосомы имеют сферическую или слегка эллипсоидную форму. Полисома, или полирибосома — несколько рибосом, одновременно транслирующих одну молекулу мРНК. Рибосомы митохондрий, или миторибосомы, ассоциированы с митохондриальным матриксом. Основная часть: Рибосома- немембранный двухсубъкдиничный (малая и большая субъединицы) органоид, состоящий из рРНК и белков. Рибосомы имеют сферическую или слегка эллипсоидную форму. Служит для биосинтеза белка из аминокислот. Рибосомы подразделяются на свободные и связанные с мембранами ЭПС и наружной ядерной мембраной. Свободные рибосомы синтезируют синтезируют белки для самой клетки, а связанные- на экспорт. Синтез рибосом у эукариот происходит в специальной внутриядерной структуре —ядрышке. Полисома, или полирибосома — несколько рибосом, одновременно транслирующих одну молекулу мРНК. Рибосомы митохондрий, или миторибосомы, ассоциированы с митохондриальным матриксом. Вопрос №12 Аппарат Гольджи Краткий обзор. Аппарат Гольджи — это система внутриклеточных мембранных структур: цистерн и пузырьков, в которых накапливаются вещества, синтезированные на мембранах ЭПС. Вещества доставляются в комплекс Гольджи в мембранных пузырьках, которые отшнуровываются от эндоплазматической сети и присоединяются к цистернам комплекса Гольджи. Здесь эти вещества претерпевают различные биохимические превращения, а затем снова упаковываются в мембранные пузырьки, и большая их часть транспортируется к цитоплазматической мембране. Мембрана пузырьков сливается с цитоплазматической мембраной, а содержимое выводится за пределы клетки посредством экзоцитоза. В комплексе Гольджи растительных клеток синтезируются полисахариды клеточной стенки (оболочки). Еще одна важная функция комплекса Гольджи — это образование лизосом. Комплекс Гольджи был открыт в 1898 г. итальянским гистологом Камилло Гольджи В нервных клетках. Основная часть. Комплекс Гольджи — это место конденсации и накопления продуктов секреции, вырабатываемых в других участках клетки, в основном в ЭПС. На наружной, вогнутой стороне стопки из пузырьков постоянно формируются новые цистерны, а на внутренней стороне цистерны превращаются обратно в пузырьки. При световой микроскопии он может распределяться в виде сложных сетей или отдельных диффузно расположенных участков (диктиосом). Форма и положение органеллы не имеют принципиального значения и могут изменяться в зависимости от функционального состояния клетки. При электронной микроскопии видно, что комплекс состоит из скоплений плоских цистерн В целом комплекс Гольджи участвует в сегрегации — это разделение, отделение определенных частей от основной массы, и накоплении продуктов, синтезированных в ЭПС, в их химических перестройках, созревании. Секреторная функция комплекса Гольджи заключается в том, что синтезированный на рибосомах экспортируемый белок, отделяющийся и накапливающийся внутри цистерн ЭПС, транспортируется в вакуоли пластинчатого аппарата. Комплекс Гольджи может резко увеличиваться в размерах в клетках, активно осуществляющих секреторную функцию,обычно сопровождается развитием ЭПС, а в случае синтеза белков — ядрышка. Во время деления клетки комплекс Гольджи распадается до отдельных цистерн или пузырьков, которые распределяются между двумя делящимися клетками и в конце телофазы восстанавливают структурную целостность органеллы. Вне деления происходит непрерывное обновление мембранного аппарата за счет пузырьков, мигрирующих из ЭПС и дистальных цистерн диктиосомы за счет проксимальных компартментов. о содержимого в процессе лизосомного экзоцитоза. Вопрос 13. Лизосомы- одномембранные структуры, образуются путем слияния перинуклеарных эндосом,содержащих лизосомные гидролазы и лизосомные мембранные белки, с везикулами, подлежащими деградации ( периферической эндосомой, фагосомой или аутофагоцитозной вакуолью). * перинуклеарные эндосомы образуются при слиянии везикул, содержащие лизосомные гидролазы после их синтеза в гранулярной эпс и процессинга в комплексе Гольджи, и везикул, в мембрану которых встроены специфические лизосомные мембранные белки. * периферические эндосомы образуются в результате эндоцитоза. *мультивезикулярные тельца образуются при слиянии перинуклеарной и периферической эндосом. * фаголизосома образуется при слиянии перинуклеарной эндосомы и фагосомы. *аутофаголизосома образуется при слиянии перинуклеарной эндосомы и аутофагоцитозной вакуоли, содержащей подлежащие деградации эндогенные молекулы и органеллы. * остаточные тельца - лизосомы любого типа, содержащие непереваренный материал ( липофусцин, гемосидерин) . Функция: Катализ гидролитического ( в водной среде) расщепления нуклеиновых кислот, белков, жиров, полисахаридов и мукополисахаридов, других химических соединений при низких значениях pH. В цитобиологических понятиях - внутриклеточное переваривание веществ и структур. Пероксисомы - одномембранные органеллы, пузырьки с электроноплотной сердцевиной. В составе мембраны органеллы находятся специфичные белки - пероксины, а в матриксе - более 40 ферментов, каиализирующих анаболичиские ( биосинтез желчных кислот, H2O3, зависимое дыхание, деградация ксенобиотиков) процессы. Вопрос 14. Происхождение, строение, функции лизосом Краткий обзор: Лизосомы – одномембранные структуры, образующиеся путем слияния перинуклеарных эндосом, содержащих лизосомные гидролазы и лизосомные мембранные белки, с везикулами, подлежащими деградации, (периферической эндосомой, фагосомой или аутофагоцитозной вакуолью). Происхождение: образуются путем слияния перинуклеарных эндосом (образуются при слиянии везикул, содержащих лизосомные гидролазы, синтезированные в гранулярной ЭПС и прошедшие процессинг в комплексе Гольджи, и везикул, в мембрану которых встроены специфические лизосомные белки) с везикулами, подлежащими деградации: периферической эндосомой (образована результате эндоцитоза), фагосомой (образована в результате фагоцитоза) или аутофагоцитозной вакуолью (содержит подлежащие деградации эндогенные молекулы и органеллы). Строение: округлые пузырьки, окруженные одинарной мембраной, диаметр = 0,2-0,4 мкм. Для поддержания кислой среды внутри лизосомы в ее мембране имеется протонный насос ( 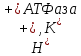 . Иногда мембрана покрыта снаружи волокнистым белковым комплексом. Лизосомы содержат различные кислые гидролазы. . Иногда мембрана покрыта снаружи волокнистым белковым комплексом. Лизосомы содержат различные кислые гидролазы.Функции: участвуют во внутриклеточном пищеварении. Участвуют в переваривании материала внутриклеточных компонентов (аутофагия) или частиц, различными путями попавших в клетку (гетерофагия). Способны к секреции своего содержимого в процессе лизосомного экзоцитоза. Полный ответ: Лизосомы – одномембранные структуры, образующиеся путем слияния перинуклеарных эндосом, содержащих лизосомные гидролазы и лизосомные мембранные белки, с везикулами, подлежащими деградации, (периферической эндосомой, фагосомой или аутофагоцитозной вакуолью). Перинуклеарные эндосомыобразуются при слиянии везикул, содержащих лизосомные гидролазы после их синтеза в гранулярной эндоплазматической сети и процессинга в комплексе Гольджи, и везикул, в мембрану которых встроены специфические лизосомные белки. Окаймленные везикулы (пузырьки) – одномембранные пузырьковидные образования, окруженные белковой оболочкой, прилежащей к наружной поверхности мембраны везикулы. Могут быть окружены клатрином (транспортируют белки из транс-компартмента комплекса Гольджи или вещества, поступающие в клетку путем эндоцитоза) или не иметь клатриновой оболочки (их оболочка из высокомолекулярного белкового комплекса – коатомера, транспортируют белки из гранулярной ЭПС в комплекс Гольджи, из одной цистерны комплекса Гольджи в другую или из комплекса Гольджи в плазматическую мембрану). Периферические эндосомы образуются в результате эндоцитоза. Аутофагоцитозная вакуоль содержит подлежащие деградации эндогенные молекулы и органеллы. Типы лизосом: Мультивезикулярные тельца образуются при слиянии перинуклеарной и периферической эндосом. Фаголизосома образуется при слиянии перинуклеарной эндосомы и фагосомы. Аутофаголизосома образуются при слиянии перинуклеарной эндосомы и аутофагоцитозной вакуоли. Остаточные тельца – лизосомы любого типа, содержащие непереваренный материал. Происхождение: образуются путем слияния перинуклеарных эндосом (образуются при слиянии везикул, содержащих лизосомные гидролазы, синтезированные в гранулярной ЭПС и прошедшие процессинг в комплексе Гольджи, и везикул, в мембрану которых встроены специфические лизосомные белки) с везикулами, подлежащими деградации: периферической эндосомой (образована результате эндоцитоза), фагосомой (образована в результате фагоцитоза) или аутофагоцитозной вакуолью (содержит подлежащие деградации эндогенные молекулы и органеллы). Строение: округлые пузырьки, окруженные одинарной мембраной, диаметр = 0,2-0,4 мкм. Для поддержания кислой среды внутри лизосомы в ее мембране имеется протонный насос ( . Иногда мембрана покрыта снаружи волокнистым белковым комплексом. Лизосомы содержат различные кислые гидролазы.Функции: участвуют во внутриклеточном пищеварении. Участвуют в переваривании материала внутриклеточных компонентов (аутофагия) или частиц, различными путями попавших в клетку (гетерофагия). Способны к секреции своего содержимого в процессе лизосомного экзоцитоза. Вопрос 15. Аксонема: молекулярное строение, роль в организации реснички и жгутика Строение аксонемы Динеин Базальное тельце Аксонема - немембранная сократительная органелла – основной структурный элемент реснички и жгутика. Состоит из 9 периферических пар микротрубочек и двух расположенных центрально одиночных микротрбуочек. Обладающий АТФазной активностью, белок динеин - компонент тубулин - динеинового хемомеханического преобразователя – входит в состав ручек, связанных с периферическими микротрубочками. Матрицей для организации аксонемы служит базальное тельце –аналог центриоли. Вопрос №16 Молекулярные моторы. Акто-миозиновый, тубулин – динеиновый, тубулин – кинезиновый. Молекулярные моторы - Это АТФазы (динеины и кинезины), одним доменом связывающиеся с тубулином микротрубочек, а другим - с различными мембранными органеллами или макромолекулами. За счёт расщепления АТФ моторные белки перемещаются вдоль микротрубочек и таким образом транспортируют органеллы и макромолекулы. а) В акто-миозиновом молекулярном моторе происходит расщепление АТФ при взаимодействии актина тонких нитей с головкой миозина, отходящей от миозиновой (толстой) нити. б) Тубулин-динеиновый хемомеханический преобразователь отвечает за направленный транспорт макромолекул и органелл к (-)-концу микротрубочек. приводит в движение жгутик сперматозоида и реснички мерцательных клеток. в) Тубулин-кинезиновый хемомеханический преобразователь обеспечивает внутриклеточный транспорт органелл и перемещение хромосом вдоль микротрубочек в ходе клеточного деления. Перемещение органелл вдоль микротрубочек с участием кинезинов осуществляется в направлении (+)-конца микротрубочек. Вопрос 17. Макромолекулярные комплексы цитоплазмы: протеосомы, апоптосомы. Кратко: апоптосомы – активаторы каспаз при регулируемой гибели клеток. протеосомы – комплексы нелизомальных протеаз, осуществляющие вместе с убиквитинами деградацию короткоживущих белков. Полный ответ: Апоптосома — крупная четвертичная белковая структура, формирующаяся внутри клетки в процессе апоптоза. Сборка апоптосомы запускается высвобождением цитохрома c из митохондрий в ответ на внутренние либо внешние про-апоптозные стимулы. Апоптосома активирует инициаторные каспазы, запускающие каскад апоптозных реакций. Протеасомы в клетках «укрывают» самокомпартментирующиеся ферменты – протеазы (пептидазы), которые участвуют во внелизосомном расщеплении белков. Протеасомы – это мультимерные гетеробелковые агрегаты «цилиндрической формы», образующихся путем самосборки. Протеазы в них занимают внутреннюю зону, а снаружи располагаются белки – «проводники» или шапероны. Шапероны выполняют функцию опознания белков, подлежащих протеолитическому расщеплению, и их «допуск» внутрь протеасомы к протеазам. Известно, что протеасомы обеспечивают деградацию циклина В в анафазе митоза. Макромолекулярные комплексы цитоплазмы: протеосомы, апоптосомы. Кратко: Протеасома —очень крупная мультисубъединичная протеаза, присутствующая в эукариот. В эукариотических клетках протеасомы содержатся и в ядре, и в цитоплазме. Основная функция протеасомы — протеолитическая деградация ненужных и повреждённых белков до коротких пептидов, которые затем могут быть расщеплены до отдельных аминокислот. Деградация 80—90 % внутриклеточных белков происходит при участии протеасомы. Для того чтобы белок-мишень расщепился протеасомой, он должен быть помечен путём присоединения к нему маленького белка убиквитина. Присоединение первой молекулы убиквитина к белку служит для лигаз сигналом для дальнейшего присоединения молекул убиквитина. В результате к белку оказывается присоединена полиубиквитиновая цепь, которая связывается с протеасомой и обеспечивает расщепление белка-мишени. В целом вся эта система получила название убиквитин-зависимой деградации белка. Протеасомальная деградация белка важна для протекания многих клеточных процессов, включая клеточный цикл, регуляцию экспрессии генов и ответ на окислительный стресс. Полный: Термин "апоптоз", предложенный в 1972 г. английскими учеными J.F.R. Кеrr, А.Н. Wyllie и A.R. Currie, состоит из двух греческих слов и означает в буквальном смысле "отделение лепестков от цветов", а применимо к клетке - особый тип смерти путем разделения ее на части ("апоптозные тельца"), которые впоследствии фагоцитируются соседними клетками разного типа, с целью не допустить появление воспалительной реакции. Неконтролируемый апоптоз может привести к раку, аутоиммунным и нейродегенеративным заболеваниям. Центральным пунктом апоптоза является митохондрия. Она играет большую роль в апоптозе, вызываемом различными раздражителями; объединяет сигналы клеточной смерти посредством белков семейства Bcl-2 (белков, в зависимости от ситуации стимулирующих или подавляющих апоптоз) и активирует выход цитохрома С(белка, переносящего электроны между белковыми комплексами во внутренней мембране митохондрии), в результате чего митохондриальная мембрана становится проницаемой. Механизмы, которые приводят к этой проницаемости, ещё не до конца изучены. Выделяясь из митохондрий, цитохром С включается в составапоптосомы. Апоптосома - крупная четвертичная белковая структура, формирующаяся внутри клетки в процессе апоптоза. Содержит Цитохром из митохондрий, выделяющийся в ответ на внутренние либо внешние про-апоптозные стимулы. Апоптосома активирует инициаторные каспазы, запускающие каскад апоптозных реакций. Протеосома - очень крупная мультисубъединичная протеаха, присутствующая в клетках. В эукариотических клетках содержатся в ядре и цитоплазме. Основная функция – протеолитическая деградация ненужных и поврежденных белков до коротких пептидов, которые затем могут быть расщеплены до отдельных аминокислот Апоптосома - крупная четвертичная белковая структура, формирующаяся внутриклеткив процессеапоптоза. Сборка апоптосомы запускается высвобождениемцитохрома cизмитохондрийв ответ на внутренние либо внешние про-апоптозные стимулы. Апоптосома активирует инициаторныекаспазы, запускающие каскад апоптозных реакций. Вопрос 18. Жидкостно-мозаичная модель биологической мембраны В 1972 г. Сингер и Николсон (Singer, Nicolson) предложили жидкостно-мозаичную модель мембраны, согласно которой белковые молекулы плавают в жидком фосфолипидном бислое. Они образуют в нем как бы своеобразную мозаику, но поскольку бислой этот жидкий, то и сам мозаичный узор не жестко фиксирован; белки могут менять в нем свое положение. Покрывающая клетку тонкая мембрана напоминает пленку мыльного пузыря — она тоже все время «переливается». Ниже суммированы известные нам данные, касающиеся строения и свойств клеточных мембран. 1. Толщина мембран составляет около 7 нм. 2. Основная структура мембраны — фосфолипидный бислой. 3. Гидрофильные головы фосфолипидных молекул обращены наружу — в сторону водного содержимого клетки и в сторону наружной водной среды. 4. Гидрофобные хвосты обращены внутрь — они образуют гидрофобную внутреннюю часть бислоя. 5. Фосфолипиды находятся в жидком состоянии и быстро диффундируют внутри бислоя — перемещаются в латеральном направлении. 6. Жирные кислоты, образующие хвосты фосфолипидных молекул, бывают насыщенными и ненасыщенными. В ненасыщенных кислотах имеются изломы, что делает упаковку бислоя более рыхлой. Следовательно, чем больше степень ненасыщенности, тем более жидкую консистенцию имеет мембрана. 7. Большая часть белков плавает в жидком фосфолипидном бислое, образуя в нем своеобразную мозаику, постоянно меняющую свой узор. 8. Белки сохраняют связь с мембраной, поскольку в них есть участки, состоящие из гидрофобных аминокислот, взаимодействующих с гидрофобными хвостами фосфо-липидов; вода из этих мест выталкивается. Другие участки белков гидрофильны. Они обращены либо к окружению клетки, либо к ее содержимому, т. е. к водной среде. 9. Некоторые мембранные белки лишь частично погружены в фосфолипидный бислой, тогда как другие пронизывают его насквозь. 10. К некоторым белкам и липидам присоединены разветвленные олигосахаридные цепочки, играющие роль антенн. Такие соединения называются соответственно гликопротеинами и гликолипидами. 11. В мембранах содержится также холестерол. Подобно ненасыщенным жирным кислотам он нарушает плотную упаковку фосфолипидов и делает их более жидкими. Это важно для организмов, живущих в холодной среде, где мембраны могли бы затвердевать. Холестерол делает мембраны также более гибкими и вместе с тем более прочными. Без него они бы легко разрывались. 12. Две стороны мембраны, наружная и внутренняя, различаются и по составу, и по функциям. Химический состав мембран.С помощью световой и электронной микроскопии в клетках выявлены разнообразные мембранные структуры. Все они имеют сходный химический состав и принцип организации, но в зависимости от типа мембран и их функций соотношение химических компонентов и детали строения могут отличаться. Мембраны состоят из липидов, белков и углеводов (рис.16). Липиды составляют в среднем 40% сухой массы мембран. Среди них преобладают фосфолипиды (до 80%). Основным функциональным компонентом биологических мембран являются белки. Но только образовав прочные комплексы с липидами, они способны проявлять активность. Поверхностные белки (около 30% от общего количества мембранных белков) размещены на наружной и внутренней поверхностях мембран и связанные с последними электрическими силами непосредственно или через двухвалентные катионы, преимущественно Са2 + и Mg2 +. Они легко отделяются от мембран после разрушения клеток. Внутренние белки (почти 70% общего количества мембранных белков) погружены в двойной слой липидов на разную глубину, а в некоторых случаях пересекают мембрану насквозь. Такие белки связывают обе поверхности мембраны. Углеводы входят в состав мембран не самостоятельно, а образуют комплексы с белками или липидами. Организация биологических мембран. Сейчас общепринятой является модель растворимо-мозаичной строения мембран (рис.16). Такое название произошло от того факта, что около 30% липидов тесно связаны с внутренними белками, а остальное - находится в жидком состоянии, где «плавают» липопротеиды. Молекулы липидов размещены в виде двойного слоя, их полярные гидрофильные «головки» обращены к внешней и внутренней сторон мембран, а гидрофобные неполярные «хвосты» - внутрь. Если посмотреть на мембрану сверху, то она напоминает мозаику, созданную полярными «головками» липидов, поверхностными и внутренними белками. Толщина мембран варьирует в довольно широких пределах в зависимости от их типа. Мембраны клеток эукариот и прокариот сходны по строению. Между молекулами белков или их частями часто существуют поры (канальцы), заполненные водой. Молекулы, входящие в состав мембран, способные перемещаться, благодаря чему мембраны быстро возобновляются за незначительных повреждений, образуются над оголенными участками цитоплазмы, могут легко сливаться друг с другом, растягиваться и сжиматься, например, при движении клеток или изменения их формы. Вопрос №19 Рецепторная функция плазматической мембраны Краткий обзор: Гликокаликс-это внешний по отношению к липопротеидной мембране слой, содержащий полисахаридные цепочки мембранных интегральных белков - гликопротеидов. Одной из важнейших функций плазмалеммы является обеспечение коммуникации (связи) клетки с внешней средой посредством присутствующего в мембранах рецепторного аппарата, имеющего белковую или гликопротеиновую природу. Основная функция рецепторных образований плазмалеммы - распознавание внешних сигналов, благодаря которым клетки правильно ориентируются и образуют ткани в процессе дифференцировки. С рецепторной функцией связана деятельность различных регуляторных систем, а также формирование иммунного ответа. Основная часть: В качестве таких рецепторов на поверхности клетки могут выступать белки мембраны или элементы гликокаликса — гликопротеиды. Такие чувствительные к отдельным веществам участки могут быть разбросаны по поверхности клетки или собраны в небольшие зоны. Разные клетки животных организмов могут обладать разными наборами рецепторов или же разной чувствительностью одного и того же рецептора. Роль многих клеточных рецепторов заключается не только в связывании специфических веществ или способности реагировать на физические факторы, но и в передаче межклеточных сигналов с поверхности внутрь клетки. В настоящее время хорошо изучена система передачи сигнала клеткам с помощью некоторых гормонов, в состав которых входят пептидные цепочки. Гормон взаимодействует специфически с рецепторной частью этой системы и, не проникая внутрь клетки, активирует аденилатциклазу (белок, лежащий уже в цитоплазматической части плазматической мембраны), которая синтезирует циклический АМФ. Последний активирует или ингибирует внутриклеточный фермент или группу ферментов. Таким образом, команда (сигнал от плазматической мембраны) передается внутрь клетки. Эффективность этой аденилатциклазной системы очень высока. Так, взаимодействие одной или нескольких молекул гормона может привести за счет синтеза множества молекул цАМФ к усилению сигнала в тысячи раз. В данном случае аденилатциклазная система служит преобразователем внешних сигналов. Разнообразие и специфичность наборов рецепторов на поверхности клеток приводят к созданию очень сложной системы маркеров, позволяющих отличать свои клетки (той же особи или того же вида) от чужих. Сходные клетки вступают друг с другом во взаимодействия, приводящие к слипанию поверхностей (конъюгация у простейших и бактерий, образование тканевых клеточных комплексов). При этом клетки, отличающиеся набором детерминантных маркеров или не воспринимающие их, либо исключаются из такого взаимодействия, либо (у высших животных) уничтожаются в результате иммунологических реакций. С плазматической мембраной связана локализация специфических рецепторов, реагирующих на физические факторы. Так, в плазматической мембране или в ее производных у фотосинтетических бактерий и синезеленых водорослей локализованы белки-рецепторы (хлорофиллы), взаимодействующие с квантами света. В плазматической мембране светочувствительных клеток животных расположена специальная система фоторецепторных белков (родопсин), с помощью которых световой сигнал превращается в химический, что в свою очередь приводит к генерации электрического импульса. |