Лекции-биология. Курс лекций для студентов, обучающихся на русском языке Рязань 2008
 Скачать 7.1 Mb. Скачать 7.1 Mb.
|

|
Пол как наследственный признак Одним из признаков у многих живых организмов является пол (мужской и женский). Пол - это совокупность морфологических, физиологических, биохимических и поведенческих признаков организма, которые обуславливают репродукцию (воспроизведение). Пол - это генетически обусловленный признак, т.е. пол зародыша определяется генетическими механизмами - комбинацией половых хромосом. Половые хромосомы определяют первичные половые признаки. Под их влиянием формируются половые железы (семенники и яичники), выделяющие мужские и женские половые гормоны. Половые гормоны определяют развитие вторичных половых признаков того или другого пола и проявляются у организмов в период полового созревания (у человека в 12 - 15 лет). Вторичные половые признаки - различие в размерах и пропорциях тела, тембр голоса, степень развития волосяного покрова и др. Определение пола У большинства организмов пол определяется в момент оплодотворения (сингамно) и регулируется хромосомным набором зиготы, его называют хромосомный тип определения пола. У человека и млекопитающих женские особи имеют в кариотипе две Х - хромосомы (ХХ), а мужские Х- и У- хромосомы. Женские особи являются гомогаметными, поскольку формируют лишь один тип гамет, а мужские - гетерогаметными, т.к. образуют два типа гамет. У птиц и бабочек гетерогаметным полом является женский пол, а гомогаметными - мужской. У них половые хромосомы обозначают как Z и W. У некоторых клопов, жуков самки имеют состав половых хромосом ХХ, самцы же несут всего одну из них - (ХО). Совершенно особый тип определения пола имеется у перепончатокрылых, в частности, у пчел. У этих насекомых самки развиваются из оплодотворенных яиц и клетки их тела диплоидны, а самцы развиваются партеногенетически из неоплодотворенных яиц и имеют гаплоидные клетки. Хромосомная теория наследования пола утверждает, что гены определяющие развитие пола, локализованы в половых хромосомах. У человека гены, обуславливающие развитие женского пола, находятся в Х - хромосоме, а гены мужского пола - в У - хромосоме. Основные типы хромосомного определения пола 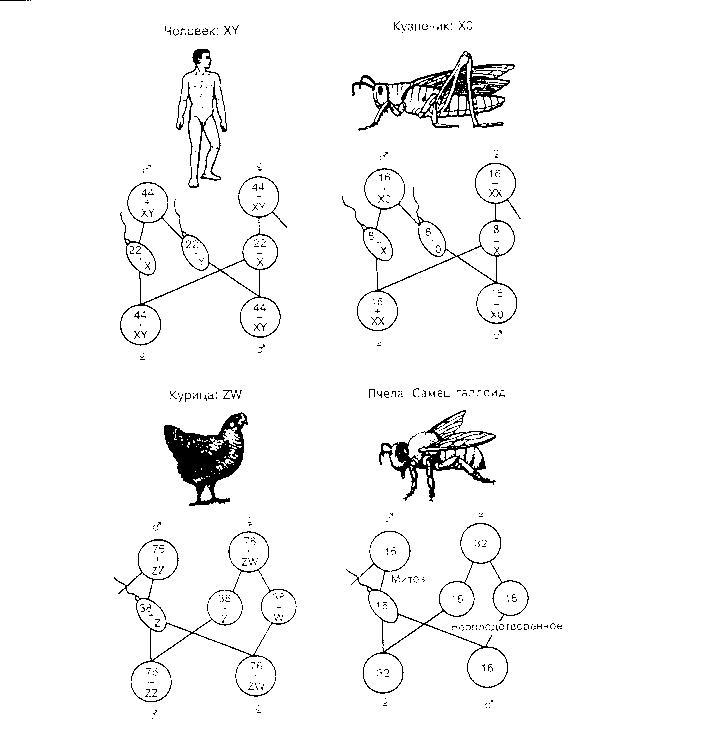 Иначе обстоит дело у дрозофилы. При изучении наследования пола у мухи - дрозофилы было установлено, что у самки две одинаковые Х - хромосомы, а у самца Х- и У- - хромосомы. У дрозофилы гены, определяющие женский пол, находятся в Х - хромосоме, а У - хромосома генетически инертна и гены, определяющие развитие мужского пола, находятся в аутосомах. Оплодотворенные яйцеклетки, имеющие диплоидный набор хромосом и две Х - хромосомы дают начало самкам (2А + ХХ), а имеющие диплоидный набор аутосом и одну Х - хромосому дают самцов(2А+ХУ). Опыты подтвердили, что чем больше наборов аутосом, тем больше были выражены признаки мужского пола. Из этого был сделан вывод, что У - хромосома у мухи - дрозофилы не имеет существенного значения для определения мужского пола. Кроме сингамного способа определения пола существует эпигамный способ дифференцировки пола, который определяется воздействием условий окружающей среды, например, у морского кольчатого червя Bonnelia определяется условиями развития яйцеклеток (прогамный способ, как у коловраток). Наследование признаков, сцепленных с полом и ограниченных полом Сцепленными с полом называют признаки, развитие которых обусловлено генами, расположенными в половых хромосомах. Если ген находится в У-хромосоме, то он наследуется у человека, млекопитающих - от отца к сыну, а у птиц - от матери к дочери. Если ген располагается в Х-хромосоме, наследование будет более сложным. Впервые особенности наследования генов, сцепленных с Х-хромосомой были открыты в опытах на дрозофиле при изучении наследования окраски глаз. Красная окраска глаз доминирует над белой. При скрещивании гомозиготной красноглазой самки с белоглазым самцом в F1 все потомство красноглазое. Р ♀ ХАХА х ♂ХªУ кр. бел. G (XА) (Xª) (У) F1 XАXª х XАУ кр. кр. G (ХА) (Xª) (XА) (У) F2 ХАХА , ХАХª, ХАУ, ХªУ кр. кр. кр. бел. При скрещивании гибридов первого поколения друг с другом в F2 происходит расщепление по фенотипу: 3 красноглазые особи: 1 белоглазая, и белоглазыми бывают только самцы. При реципрокном (обратном) скрещивании, когда самка гомозиготная по гену белоглазости, скрещивается с красноглазым самцом, расщепление в F1 по фенотипу 1:1. При этом белоглазыми будут только самцы. При скрещивании мух F1 между собой в F2 будут мухи с обоими признаками в равном отношении 1:1, как среди самок, так и среди самцов. Р ♀ ХªХª х ♂ХА У бел. кр. G (Хª) (ХА) (У) F1 ХАХª х ХªУ кр. бел. G (ХА) (Хª) (Хª) (У) F2 ХАХª, ХªХª, ХАУ, ХªУ кр. бел. кр. бел. Из результатов опыта Моргана можно заключить, что гены окраски глаз расположены только в половой Х-хромосоме. Результаты скрещивания зависят от того, какой пол несет доминантный признак. Наследование, сцепленное с полом, необходимо отличать от наследования, ограниченного полом. Развитие признаков (ограниченных полом) обусловлено генами, расположенными в аутосомах (а не в половых хромосомах), но на проявление признаков в фенотипе сильно влияет пол, т.е. развитие признака зависит от влияния половых гормонов. Например, облысение со лба, типично для мужчин. У мужчин под действием мужских половых гормонов (тестостерона) ген ведет себя как доминантный. Еще пример, тембр голоса - баритон и бас - характерны только для мужчин. Сцепление генов. Опыты и правило Моргана Изучение сцепленного с полом наследования стимулировало изучение сцепления между генами, находящимися в аутосомах. Для любого организма характерно видовое постоянство хромосом в кариотипе. Генов, определяющих признаки, у организмов намного больше, чем хромосом. Например, у мухи дрозофилы 8 хромосом в соматических клетках, а генов около 1000. Значит, в каждой хромосоме находится много генов. Гены, локализованные в одной хромосоме, образуют группы сцепления. Число групп сцепления равно гаплоидному числу хромосом. Наследование генов, локализованных в одной хромосоме, называется сцепленным наследованием. В начале ХХ века Т. Морган и его сотрудники описали явление сцепления генов - совместную передачу групп генов из поколения в поколение. Опыты проводились на мухах дрозофилах с учетом двух пар альтернативных признаков: Цвет тела – серый (В) и черный (в) Длина крыльев – нормальные (V) и короткие (v) У мухи - дрозофилы окраски тела и длины крыльев находятся в одной паре гомологичных хромосом, т.е. относятся к одной группе сцепления, что и было доказано в опытах. Запись генотипов при сцеплении видоизменяется: генотип записывается BV ═ bv (две черточки означают, что организм диплоидный). При скрещивании мух, имеющих серый цвет тела и нормальные крылья с мухами черного цвета и короткими крыльями в F1 все мухи имели серый цвет тела и нормальные крылья. Запись схемы скрещивания: P ♀BV ♂bv ═ x ═ BV bv сер.нор. чер.кор. G (BV) (bv) BV F1 ═ bv Провели анализирующее скрещивание. В первом случае скрестили гибридного самца с серым телом и нормальными крыльями с черной самкой, имеющей короткие крылья, и в результате скрещивания получили 2 типа потомков, похожих на родителей в соотношении 1:1. Ген серого цвета тела и ген нормального строения крыльев передаются вместе, а ген черного цвета тела и ген коротких крыльев тоже вместе. Этот опыт демонстрирует полное сцепление. Причина его заключается в том, что гены, обуславливающие два различных признака, лежат в одной хромосоме. Это видно на схеме полного сцепления: Р ♀bv ♂BV ═ x ═ bv bv G ( bv) ( BV ) (bv) BV bv F1 ═ ═ bv bv 1:1 Другой результат получается, если для анализирующего скрещивания брать гибридную самку с серым телом и нормальными крыльями и рецессивного по обоим признакам самца. В этом случае появилось четыре типа потомков в соотношении: серых длиннокрылых - 41, 5 %; серых короткокрылых - 8, 5 %, черных длиннокрылых - 8, 5% , черных короткокрылых - 41, 5 %. В этом случае имеет место неполное сцепление: Р ♀BV ♂ bv ═ x ═ bv bv G ( BV) (bv) некроссов. ( bv) (Bv) (bV) кроссов. BV bv Bv bV F1 ═ ═ ═ ═ bv bv bv bv 41,5% 41,5% 8,5% 8,5% Причиной неполного сцепления генов является кроссинговер (перекрест хромосом) и обмен участками между гомологичными хромосомами в профазе I деления мейоза при созревании половых клеток. Кроссинговер происходит у самок, у самцов дрозофил кроссинговер не происходит (установлено, что для особей гетерогаметного пола характерна более низкая частота кроссинговер). При изучении результатов скрещивания получается 4 фенотипа среди потомков: 17%(8, 5+8, 5) особей образовались из кроссоверныхгамет 83%(41, 4+41, 5) особей образовались из некроссоверныхгамет. Частота кроссинговера зависит от расстояния между генами в хромосоме. Расстояние между генами выражается в процентах кроссинговера между ними и обозначается в морганидах. Изучение Морганом сцепления генов представляет собой закономерное биологическое явление. Гены, локализованные в одной хромосоме, наследуются, сцеплено, причем сила сцепления зависит от расстояния между генами. Эта закономерность получила название правило Моргана. . Основные положения хромосомной теории наследственности Основные положения хромосомной теории наследственности сводятся к следующему: - носителями наследственной информации являются хромо- сомы и расположенные в них гены, - гены расположены в хромосоме в линейном порядке друг за другом в определенных локусах. Аллельные гены занимают одинаковые локусы гомологичных хромосом, - гены, расположенные в одной хромосоме, образуют группы сцепления и наследуются преимущественно вместе. Число групп сцепления равно гаплоидному набору хромосом, - между гомологичными хромосомами возможен обмен участ- ками - кроссинговер, который нарушает сцепление генов. Сила сцепления зависит от расстояния между генами, - процент кроссинговера пропорционален расстоянию между генами. За единицу расстояния принимается 1 морганида, которая равна 1% кроссинговера, - при неполном сцеплении в сумме вероятность некроссовер- ных гамет (гибридов) всегда больше, чем 50%, - при расстоянии в 50 морганид и больше признаки наследуют- ся независимо, несмотря на то, что локализованы в 1 хромо- соме. Лекция 11 Молекулярная генетика План 1. Этапы развития молекулярной генетики. 2. Генетический код и его свойства. 3. Функционально-генетическая классификация генов. 4. Функциональная активность генов (экспрессия генов). 5. Регуляция экспрессии генов у про - и эукариот. Молекулярная генетика занимается изучением структурно-функциональной организации генетического аппарата клеток и механизма реализации наследственной информации. Этапы развития молекулярной генетики Молекулярная генетика выделилась из биохимии и сформировалась как самостоятельная наука в 50-х годах прошлого столетия. Рождение этой науки связано с рядом важных биологических открытий: 1). В 20-40 гг. ХХ века было установлено, что носителем наследственной информации в клетке является молекула ДНК, а не белок, как считали раньше. Были получены прямые доказательства роли ДНК в наследственности. Это явления трансформации, трансдукции, половой процесс у бактерий, строение вируса, а также обнаружение почти полной идентичности химического состава ДНК (но не белков) у всех представителей данного биологического вида. Данные о видовой специфичности строения ДНК получены в начале 50-х годов Эрвином Чаргаффом и его сотрудниками. Любая ДНК состоит из четырех типов нуклеотидов, содержащих одно из четырех азотистых оснований: аденин, тимин, гуанин, цитозин. Чаргафф с сотрудниками обнаружил, что соотношения между нуклеотидами, входящими в состав ДНК одинаковы в пределах вида и различаются у разных видов. Кроме того, число адениловых нуклеотидов равно числу тимидиловых, а число гуаниловых равно числу цитозиловых. Это открытие послужило главным ключом к выявлению структуры молекулы ДНК. Трансформация у бактерий – это передача от одной бактериальной клетки другой наследственного материала. В 1928 году Фред Гриффит изучал трансформацию у двух штаммов пневмококка: вирулентного (вызывающего гибель мышей) и авирулентного (не вызывающего гибели). Если убить вирулентные бактерии и вводить их вместе с живыми авирулентными, то часть животных все равно погибнет. Этот опыт свидетельствовал о том, что часть генетического материала из вирулентных бактерий проникла в авирулентные и сделала их вирулентными. После того как вирулентные клетки разделили на отдельные компоненты: углеводы, липиды, белки и ДНК, было обнаружено, что именно ДНК наделило живые бактерии генетическим признаком, которого у них до этого не было. Трансдукция – это явление, заключающееся в том, что вирусы, выходя из бактериальной клетки, в которой они паразитировали, могут захватывать с собой часть ДНК и, перемещаясь в новые клетки передавать им признаки прежних хозяев. Так, при внесении в неподвижные бактерии вирусов, размножавшихся в клетках подвижных культур, некоторые из бактерий становятся подвижными. Половой процесс у бактерий – коньюгация, когда происходит обмен молекулами ДНК, и хотя новые бактерии не образуются, но их наследственный материал изменяется, т.к. происходит рекомбинация генетического материала. Строение вируса или бактериофага: вирус состоит из молекулы ДНК (или РНК), заключенной в белковую оболочку. С помощью радиоизотопов было доказано, что именно ДНК проникает в клетку, а белковая оболочка остается снаружи. В результате образуются новые фаговые частицы. Таким образом, ДНК является носителем генетической информации и дает начало новым бактериофагам. Косвенные доказательства: в соматических клетках разных органов содержится одинаковое количество ДНК, вдвое большее, чем в половых. Количество белков варьирует и не всегда больше, чем в половых. 2).Следующий этап развития молекулярной генетики связан с таким важным открытием, как установление структурной организации молекулы ДНК– в 1953 году. Крик и Уотсон установили, что ДНК состоит из двух спирально-закрученных цепей. В настоящее время принято говорить о первичной, вторичной и третичной структуре ДНК: Первичной структурой ДНК называют линейную полинуклеотидную цепь, в которой мононуклеотиды соединены 3`,5`- фосфоэфирными связями. Модель Крика и Уотсона – это вторичная структура ДНК, основанная на принципе комплементарности и получившая название двойной спирали. Один виток спирали состоит из 10 нуклеотидов, размер витка 3,4 нм. Дополнительное структурирование в пространстве двуспиральной молекулы ДНК (в конденсированных хромосомах) с образованием суперспирали является третичной структурой. 3).Определение направления передачи информации – «один ген – один фермент» - Бидл и Татум – 50-е годы. (В настоящее время – один ген – один полипептид). 4).Расшифровка генетического кода – Ниренберг, Очоа (К 1964 году расшифрованы коды для всех аминокислот). Выяснение механизма экспрессии генов у прокариот и ее регуляции – Франсуа Жакоб и Жан Моно – 50-е годы. 5).70-е годы и до настоящего времени – выявление особенностей экспрессии генов у эукариот. Развитие генетической инженерии. Генетический код и его свойства Генетический код – это система записи информации о последовательности расположения аминокислот в белках с помощью последовательности расположения нуклеотидов в молекуле ДНК. Свойства генетического кода: 1.Код триплетен – каждая аминокислота зашифрована последовательностью трех нуклеотидов, называемых триплетом или кодоном. 2.Код вырожден – каждая аминокислота кодируется более чем одним кодоном. (Исключения – аминокислоты метионин и триптофан. Они кодируются одним кодоном.) 3.Код универсален – одни и те же триплеты кодируют одни и те же аминокислоты у всех организмов. 4.Генетический код не перекрывающийся – триплеты нуклеотидов не перекрывают друг друга. У некоторых вирусов обнаружено перекрывание, но это исключение. 5.Имеются стартовый и терминальный кодоны. Стартовый – единый для всех – АУГ. Терминальный трех видов – УАГ, УАА, УГА. Функционально-генетическая классификация генов В настоящее время ген рассматривается как единица функционирования наследственного материала. Ген – это участок молекулы ДНК, ответственный за синтез одного полипептида. Различают три вида генов: - структурные; - модуляторы; - регуляторы. Структурные гены несут информацию об аминокислотах в белках, ферментах, а также о последовательности нуклеотидов в молекулах р-РНК и т-РНК. Гены-модуляторы влияют на функционирование структурных генов, могут смещать в ту или иную сторону процесс развития признака. Они подразделяются на: 1)ингибиторы или супрессоры (эпистатичные гены), 2)интенсификаторы – могут повышать способность структурных генов к мутациям, 3)модификаторы – могут влиять на структурные гены по типу комлементарности. Гены-регуляторы контролируют синтез регуляторных белков, а также время включения различных структурных генов в процессе индивидуального развития. Свойства генов: 1. Дискретность действия – каждый ген действует как самостоятельная единица наследственности. 2. Стабильность – при отсутствии мутаций ген передается в ряду поколений в неизмененном виде. 3. Специфичность действия – каждый ген влияет на развитие своего определенного признака. 4. Плейотропия – способность одного гена обеспечивать развитие одновременно нескольких признаков (синдром Марфана). 5. Присутствие в виде двух у диплоидных и большего числа аллелей у полиплоидных организмов. 6. Действие гена дозировано, при изменении числа доз гена в организме изменяется признак (например, при болезни Дауна происходит увеличение до трех доз генов 21 хромосомы) |