Курс лекций по нормальной физиологии. Ю. И. Савченков. Красноярск Издво , 2012, 470 с
 Скачать 8.6 Mb. Скачать 8.6 Mb.
|

34. 2. Механизм образования условных рефлексов. “Замыкание” временной связи (И.П. Павлов, Э.А. Асратян, П.К. Анохин).Одним из основных элементарных актов высшей нервной деятельности является условный рефлекс. Биологическое значение условных рефлексов заключается в резком расширении числа сигнальных, значимых для организма раздражителей, что обеспечивает несравненно более высокий уровень адаптивного (приспособительного) поведения. Условно-рефлекторный механизм лежит в основе формирования любого приобретенного навыка, в основе процесса обучения. Структурно-функциональной базой условного рефлекса служат кора и подкорковые образования мозга. Сущность условно-рефлекторной деятельности организма сводится к превращению индифферентного раздражителя в сигнальный, значащий, благодаря многократному подкреплению раздражения безусловным стимулом. Благодаря подкреплению условного стимула безусловным ранее индифферентный раздражитель ассоциируется в жизни организма с биологически важным событием и тем самым сигнализирует о наступлении этого события. При этом в качестве эффекторного звена рефлекторной дуги условного рефлекса может выступать любой иннервируемый орган. В организме человека и животных нет органа, работа которого не могла бы измениться под влиянием условного рефлекса. Любая функция организма в целом или отдельных его физиологических систем может быть модифицирована (усилена или подавлена) в результате формирования соответствующего условного рефлекса. Физиологический механизм, лежащий в основе условного рефлекса, схематически представлен на рис. 68. В зоне коркового представительства условного стимула и коркового (или подкоркового) представительства безусловного стимула формируются два очага возбуждения. Очаг возбуждения, вызванный безусловным стимулом внешней или внутренней среды организма, как более сильный (доминантный) притягивает к себе возбуждение из очага более слабого возбуждения, вызванного условным стимулом. После нескольких повторных предъявлений условного и безусловного раздражителей между этими двумя зонами «проторяется» устойчивый путь движения возбуждения: от очага, вызванного условным стимулом, к очагу, вызванному безусловным стимулом. В результате изолированное предъявление только условного стимула теперь приводит к реакции, вызываемой ранее безусловным стимулом. Р 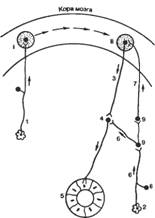 ис. 68. Образование условного рефлекса (схема). I, II — очаги возбуждения в коре большого мозга; 1 — афферентные пути условного сигнала; 2 — рецепторы полости рта; 3 — эфферентные пути; 4 — центр в продолговатом мозге; 5 — слюнная железа; 6, 7 — афферентные пути безусловного сигнала; 8 — чувствительные нейроны; 9 — вставочные нейроны. ис. 68. Образование условного рефлекса (схема). I, II — очаги возбуждения в коре большого мозга; 1 — афферентные пути условного сигнала; 2 — рецепторы полости рта; 3 — эфферентные пути; 4 — центр в продолговатом мозге; 5 — слюнная железа; 6, 7 — афферентные пути безусловного сигнала; 8 — чувствительные нейроны; 9 — вставочные нейроны.В качестве главных клеточных элементов центрального механизма образования условного рефлекса выступают вставочные и ассоциативные нейроны коры большого мозга. И.П. Павлов долгое время считал, что УР-связь замыкается между корковым центром, воспринимающим условный раздражитель, и подкорковым центром безусловного рефлекса. Однако позднее, когда были накоплены новые экспериментальные данные, Павлов пришел к выводу, что замыкание временной связи происходит полностью в коре, между корковым концом анализатора, воспринимающего условный раздражитель, и корковым представительством безусловного рефлекса (кора — кора). После удаления коры условные рефлексы исчезают, а безусловные становятся более грубыми и инертными, менее точными и совершенными, хуже приспособленными к качеству, силе и продолжительности раздражителей. Если удалить у собаки одно полушарие головного мозга, то на поврежденной стороне, несмотря на полную сохранность безусловно-оборонительного рефлекса, не может быть выработан не его базе условный рефлекс, тогда как на здоровой стороне он легко вырабатывается. Последующие нейрофизиологические исследования привели к разработке, экспериментальному и теоретическому обоснованию нескольких различных гипотез об образовании условного рефлекса. Данные современной нейрофизиологии указывают на возможность разных уровней замыкания, формирования условно-рефлекторной связи (кора — кора, кора — подкорковые образования, подкорковые образования — подкорковые образования) при доминирующей роли в этом процессе корковых структур. Физиологический механизм образования условного рефлекса представляет собой сложную динамическую организацию корковых и подкорковых структур мозга. Дальнейшие исследования структуры дуги безусловного рефлекса показали, что она в своей центральной части состоит из многих параллельных ветвей, проходящих через различные уровни ЦНС. Процесс образования условного рефлекса можно рассматривать как корковый синтез двух (или нескольких) безусловных рефлексов. Такое представление о месте и характере замыкания временной связи подтверждается экспериментальными исследованиями Э.А. Асратяна. Если подобрать два безусловных раздражителя примерно одинаково физиологической силы, например, пищевой и оборонительный, а затем сочетать их строго попеременно (то в одной, то в обратной последовательности), то образуется условный рефлекс в необычной форме. Каждый из безусловных раздражителей будет вызывать как двигательную, так и секреторную реакцию. Электрокожное раздражение становится сигналом пищи, а пища - сигналом электрокожного раздражения. Следовательно временные связи могут быть двусторонними. При исследовании роли РФ было установлено, что разрез по всей толще коры полушарий, разобщающий друг от друга центры условного и безусловного раздражителей, не препятствуют осуществлению ранее выработанного рефлекса или образованию нового. Эти факты послужили основанием для утверждения, что замыкание временной связи может осуществляться в РФ по схеме: кора-подкорка-кора. Однако приведенный факт не может служить еще бесспорным и прямым доказательством того, что временные связи замыкаются не в коре. Так, показано, например, что двигательный пищевой условный рефлекс на свет сохраняется у животного после операции "подрезки" участков зрительной и двигательной зон коры, т.е. когда пересекались пути, восходящие к этим участкам от РФ. По-видимому, более верным будет считать, что временные связи могут образовываться на различных уровнях ЦНС, и, как показано в последнее время, даже на уровне вегетативных ганглиев, однако инициативная и ведущая роль в этом процессе в естественных условиях принадлежит корковым клеткам. На основании морфологических и электрофизиологических исследований, в частности, электрической активности клеток различных слоев коры во время образования УР высказывается предположение, что временная связь замыкается, вероятно, на клеточных телах вставочных нейронов. Сам процесс замыкания временной связи заключается, по мнению многих ученых, в том, что ранее бездействующие синапсы вставочных нейронов становятся проходимыми для нервных импульсов при сочетании условного и безусловного раздражителей. Согласно гипотезе конвергентного замыкания условного рефлекса П.К. Анохина индифферентный и безусловный раздражители вызывают генерализованную активацию коры (благодаря участию РФ) т.е. каждый из них вызывает сразу возбуждение разнообразных нейронов в различных областях коры. При сочетании этих раздражителей имеет место конвергенция двух восходящих возбуждений (индифферентного и безусловного) по клеткам коры больших полушарий. Происходит перекрытие этих возбуждений на одном и том же корковом нейроне и их взаимодействие. В результате такого взаимодействия возникают и стабилизируются временные связи на различных корковых элементах, находящихся в различных участках мозга. Физиологический механизм замыкания временной связи. Образование временной связи, по Павлову, является результатом взаимодействия двух одновременно возбуждаемых пунктов коры. Наличие двух очагов возбуждения в коре закономерно вызовет движение процесса возбуждения от более слабого (вызванного индифферентным раздражителем) к более сильному (вызванному безусловным раздражителем). Таким образом, в основе механизма замыкания временной связи Павлов усматривал явление типа проторения пути, суммационного рефлекса, доминанты. Опыты Русинова показывают, что если действием постоянного тока на участок коры мозга вызвать в нем повышенную возбудимость, то он приобретает свойства доминантного и любое раздражение прежде всего вызывает теперь реакцию, связанную с возбуждением такого очага. Если это двигательный центр, то звук вызывает движение лапы. Искусственно созданный доминантный очаг сохраняется некоторое время и после выключения тока. Однако обычно доминантный очаг сохраняется сравнительно недолго, тогда как образовавшаяся временная связь является стойкой. Предполагают поэтому, что доминантный механизм играет роль лишь в первой стадии образования условного рефлекса, в процессе прокладывания временной связи, т.е. в образовании проходимости ранее бездействовавших синапсов вставочных нейронов. Беритов и Ройтбак основным в процессе замыкания временной связи считают морфологические перестройки в ЦНС под влиянием сочетаний (утолщение нейрофибрилл, миэлинизация нервных пресинаптических волокон, появление новых синапсов и т.п.). 34. 3. Учение И.П.Павлова о торможении условных рефлексов: виды торможения, условия их возникновения, примеры; механизм, биологическое значение. Движение и взаимодействие процессов возбуждения и торможения Процессы торможения в коре больших полушарий головного мозга. В основе образования условного рефлекса лежат процессы взаимодействия возбуждений в коре головного мозга. Однако для успешного завершения процесса замыкания временной связи необходимо не только активация участвующих в этом процессе нейронов, но и угнетение деятельности тех корковых и подкорковых образований, которые препятствуют этом процессу. Такое угнетение осуществляется благодаря участию процесса торможения. По своему внешнему проявлению торможение противоположно возбуждению. При нем наблюдается ослабление или прекращение деятельности нейронов, или предотвращается возможное возбуждение. Корковое торможение принято подразделять на безусловное и условное, приобретенное. К безусловным формам торможения относятся внешнее, возникающее в центре в результате его взаимодействия с другими деятельными центрами коры или подкорки, и запредельное, которое возникает в корковых клетках при чрезмерно сильных раздражениях. Эти виды (формы) торможения являются врожденными и проявляются уже у новорожденных. Внешнее безусловное торможение проявляется в ослаблении или прекращении условно рефлекторных реакций при действии каких-либо посторонних раздражителей. Если у собаки вызвать УР на звонок, а потом подействовать сильным посторонним раздражителем (боль, запах), то начавшееся слюноотделение прекратится. Так же тормозятся и безусловные рефлексы (рефлекс Тюрка у лягушки при щипке второй лапы). Случаи внешнего торможения условно рефлекторной деятельности встречаются на каждом шагу и в условиях естественной жизни животного и человека. Сюда относится постоянно наблюдаемое снижение активности и нерешительность действий в новой, необычной обстановке, снижение эффекта или даже полная невозможность деятельности при наличии посторонних раздражителей (шум, боль, голод и т.п.). Внешнее торможение условно-рефлекторной деятельности связано с появлением реакции на посторонний раздражитель. Оно наступает тем легче, и является тем более сильным, чем сильнее посторонний раздражитель и чем менее прочен условный рефлекс. Внешнее торможение условного рефлекса наступает сразу же при первом применении постороннего раздражителя. Следовательно, способность корковых клеток впадать в состояние внешнего торможения является врожденным свойством нервной системы. Это одно из проявлений т.н. отрицательной индукции. Запредельное торможение развивается в корковых клетках при действии условного раздражителя, когда его интенсивность начинает превышать известный предел. При низком уровне работоспособности корковых клеток, например, у животных со слабой нервной системой, у старых и больных животных, наблюдается быстрое развитие запредельного торможения уже при сравнительно слабых раздражениях. То же наблюдается и у животных, доведенных до значительного нервного истощения длительным действием умеренных по силе раздражителей. Запредельное торможение имеет охранительное значение для клеток коры. Виды условного торможения. Условное (внутреннее) торможение развивается в корковых клетках при определенных условиях под влиянием тех же самых раздражителей, которые до этого вызывали условно рефлекторные реакции. При этом торможение возникает не сразу, а после более или менее длительной выработки. Внутреннее торможение, как и условный рефлекс, возникает после ряда сочетаний условного раздражителя с действием определенного тормозящего фактора. Таким фактором является отмена безусловного подкрепления, изменение его характера и т.п. В зависимости от условия возникновения различают следующие виды условного торможения: угасательное, запаздывательное, дифференцировочное и сигнальное ("условный тормоз"). Угасательное торможение развивается при неподкреплении условного раздражителя. Оно не связано с утомлением корковых клеток, так как столь же длительное повторение условного рефлекса с подкреплением не ведет к ослаблению условной реакции. Угасательное торможение развивается тем легче и быстрее, чем менее прочен условный рефлекс, и чем слабее безусловный, на базе которого он выработан. Угасательное торможение развивается тем быстрее, чем меньше интервал между повторяемыми без подкрепления условными раздражителями. Посторонние раздражители вызывают временное ослабление, и даже полное прекращение угасательного торможения, т.е. временное восстановление угашенного рефлекса (растормаживание). Развившееся угасательное торможение вызывает угнетение и других условных рефлексов, слабых и тех, чьи центры расположены близко к центру первично угашенных рефлексов (это явление называется вторичным угашением). Угашенный условный рефлекс через некоторое время сам по себе восстанавливается, т.е. угасательное торможение исчезает. Это доказывает, что угасание связано именно с временным торможением, не с разрывом временной связи. Угашенный условный рефлекс восстанавливается тем быстрее, чем он прочнее и чем слабее он был заторможен. Повторное гашение условного рефлекса происходит быстрее. Развитие угасательного торможения имеет большое биологическое значение, т.к. оно помогает животным и человеку освобождаться от ранее приобретенных условных рефлексов, ставших бесполезными в новых, изменившихся условиях. Запаздывательное торможениеразвивается в корковых клетках при отставлении подкрепления во времени от начала действия условного раздражителя. Внешне это торможение выражается в отсутствии условно рефлекторной реакции в начале действия условного раздражителя и появлении ее после некоторой задержки (запаздывания), причем время этого запаздывания соответствует продолжительности изолированного действия условного раздражителя. Запаздывательное торможение развивается тем быстрее, чем меньше отставание подкрепления от начала действия условного сигнала. Посторонние раздражители вызывают временное растормаживание запаздывательного торможения. Благодаря его развитию условный рефлекс становится более точным, приурочиваясь к нужному моменту при отдаленном условном сигнале. В этом его большое биологическое значение. Дифференцировочное торможение развивается в корковых клетках при перемежающемся действии постоянно подкрепляемого условного раздражителя и неподкрепляемых сходных с ним по физическим свойствам раздражителей. Вновь образовавшийся УР имеет обычно обобщенный, генерализованный характер, т.е. вызывается не только специфическим условным раздражителем (например, тоном 50 гц), но многочисленными сходными с ним раздражителями, адресованными к тому же анализатору (тоны 10-100 гц). Однако если в дальнейшем будут подкрепляться только звуки с частотой 50 гц, а другие оставляться без подкрепления, то через некоторое время реакция на сходные раздражители исчезнет. Иначе говоря, из массы похожих раздражителей нервная система будет реагировать только на подкрепляемый, т.е. биологически значимый, а реакция на другие раздражители затормаживается. Это торможение обеспечивает специализацию условного рефлекса, жизненно важное различение, дифференцировку раздражителей по их сигнальному значению. Дифференцировка вырабатывается тем легче, чем больше разница между условными раздражителями. С помощью этого торможения можно исследовать способность животных различать звуки, фигуры, цвет и т.п. Так, по данным Губергрица, собака может отличить круг от эллипса при соотношении полуосей 8:9. Посторонние раздражители вызывают растормаживание дифференцировочного торможения. Голодание, беременность, невротические состояния, утомление и т.п. также могут приводить к растормаживанию и извращению ранее выработанных дифференцировок. Сигнальное торможение("условный тормоз"). Торможение типа "условный тормоз" развивается в коре при неподкреплении условного раздражителя в сочетании в каким-нибудь добавочным раздражителем, причем условный раздражитель подкрепляется только тогда, когда он применяется изолированно. В этих условиях условный раздражитель в сочетании в посторонним становится, в результате развития дифференцировки, тормозным, а сам посторонний раздражитель приобретает свойство тормозного сигнала (условного тормоза), он становится способным затормаживать любой другой условный рефлекс, если его присоединить к условному сигналу. Сигнальное торможение легко развивается тогда, когда условный и прибавочный раздражитель действуют одновременно. У собаки оно не вырабатывается, если этот интервал больше10 сек. Посторонние раздражители вызывают растормаживание сигнального торможения. Движение и взаимодействие процессов возбуждения и торможения в коре больших полушарий головного мозга. Высшая нервная деятельность определяется сложным взаимоотношением процессов возбуждения и торможения, возникающих в корковых клетках под влиянием разнообразных воздействий из внешней и внутренней среды. Во-первых, нервные процессы могут распространяться (иррадиировать) из места их возникновения на окружающие нервные клетки, причем иррадиация сменяется через некоторое время обратным движением нервных процессов и их сосредоточением в исходном пункте (концентрация). Во-вторых, нервные процессы при своем сосредоточении в определенном месте коры могут вызывать (индуцировать) возникновение противоположного нервного процесса в окружающих соседних пунктах коры (пространственная индукция), а после прекращения нервного процесса индуцировать противоположный нервный процесс в том же самом пункте (временная, последовательная индукция). Иррадиация нервных процессов зависит от их силы. При слабой или высокой интенсивности отчетливо выражена тенденция к иррадиации. При средней силе - к концентрации. По данным Когана, процесс возбуждения иррадиирует по коре со скоростью 2-5 м/сек, тормозный - много медленнее (несколько миллиметров в секунду). Усиление или возникновение процесса возбуждения под влиянием очага торможения называют положительной индукцией. Возникновение или усиление тормозного процесса вокруг (или после) возбуждения называется отрицательной индукцией. Положительная индукция проявляется, например, в усилении условно- рефлекторной реакции после применения дифференцировочного раздражителя или возбуждение перед сном.. Одним из часто встречающихся проявлений отрицательной индукции является торможение УР при действии посторонних раздражителей. При слабых или чрезмерно сильных раздражителях индукция отсутствует. Можно полагать, что в основе явлений индукции лежат процессы, аналогичные электротоническим изменениям. Иррадиация, концентрация и индукция нервных процессов тесно связаны друг с другом, взаимно ограничивая, уравновешивая и укрепляя друг друга, и таким образом обусловливая точное приспособление деятельности организма к условиям внешней среды. |