Лекция 09 (Патологии нервной системы). Лекции Цель
 Скачать 0.54 Mb. Скачать 0.54 Mb.
|

ТИПОВЫЕ ПАТОЛОГИЧЕСКИЕ ПРОЦЕССЫ В НЕРВНОЙ СИСТЕМЕДефицит торможения. Растормаживание В покое и деятельном состоянии нейроны испытывают постоянные тормозные влияния. При возбуждении нейронов происходит ослабление тормозных процессов. Такое растормаживание является дозированным, оно контролируется и соответствует необходимому уровню активности нейрона, поэтому имеет физиологический характер. При растормаживании, имеющем патологический характер, нейрон становится гиперактивным и выходит из-под контроля. Патологическое растормаживание возникает при значительном и неконтролируемом дефиците торможения. Такое состояние имеет место в условиях прямого повреждения тормозных механизмов, при избирательном действии на них некоторых токсинов (например, столбнячного токсина, стрихнина). Дефицит торможения и растормаживание встречаются практически при всех формах патологии нервной системы, поэтому они относятся к типовым патологическим процессам нервной системы. Дефицит торможения играет существенную роль в формировании и деятельности ГПУВ. Характерным экспериментальным синдромом растормаживания является децеребрационная ригидность. Она вызывается, по Шеррингтону, перерезкой ствола мозга между передним и задним четверохолмием. В этих условиях происходит выпадение тормозных влияний со стороны супраспинальных структур и особенно красных ядер, и проявляются возбуждающие тонические влияния вестибулярных ядер Дейтерса на мотонейроны спинного мозга, особенно γ-мотонейроны, которые в норме находятся под тормозным контролем со стороны красных ядер. Перерыв (например, путем перерезки задних корешков) расторможенной, патологически усиленной γ-петли на уровне спинного мозга ведет к исчезновению ригидности соответствующих мышц. Поэтому данный вид децеребрационной ригидности называют также γ-ригидностью (Р. Гранит).
При выпадении тормозных влияний растормаживаются и гиперактивируются прежде всего те нейроны, которые в норме находятся в состоянии тонического возбуждения. У человека и многих животных такими нейронами являются нейроны мышц, выполняющих антигравитационную функцию. Вследствие этого у децеребрированной кошки голова запрокидывается вверх, передние и задние лапы вытягиваются, хвост поднимается и т.п. У человека при выпадении моторных корковых влияний (например, после геморрагического инсульта) возникает спастическая флексорная установка верхней и экстензорная установка нижней конечностей (поза Вернике-Манна). Целый ряд патологических рефлексов возникает в условиях выпадения влияний со стороны коры и подкорковых образований; эти рефлексы являются результатом растормаживания центров спинного или продолговатого мозга. Они представляют собой гиперболизированные неконтролируемые реакции, которые являлись нормальными в раннем постнатальном периоде и затем были подавлены при развитии регулирующих влияний со стороны высших отделов ЦНС. К ним относятся рефлекс Бабинского (растопыривание пальцев ноги вместо их сгибания при раздражении подошвы), хватательный, сосательный и другие рефлексы. При полном перерыве спинного мозга могут проявляться заложенные генетически и подавленные с возрастом спинальные автоматизмы в виде сравнительно координированных сгибательноразгибательных движений конечностей. Если растормаживаются и гиперактивируются тормозные нейроны, то возникает патологически усиленный тормозной эффект, который может проявиться в виде угнетения и выпадения функции. Денервационный синдромДенервационный синдром представляет собой комплекс изменений, возникающих в постсинаптических нейронах, органах и тканях после выпадения нервных влияний на эти структуры. Денервированная структура (мышца, нейрон) приобретает повышенную чувствительность к физиологически активным веществам (закон Кеннона-Розенблюта). Основным проявлением денервационного синдрома в мышце является исчезновение концевой пластинки - зоны мышечного волокна, где сосредоточен весь его холинергический аппарат. Вместо нее появляются новые ацетилхолиновые рецепторы на всем протяжении мышечного волокна, и в связи с этим происходит повышение общей чувствительности к ацетилхолину всего волокна (А.Г. Гинецинский, Н.М. Ашмарина). Этот эффект связан главным образом с выпадением трофических влияний нерва. Другой характерный признак - фибриллярные подергивания денервированной мышцы. Этот эффект отражает реакцию мышечных денервированных волокон на поступающий к ним из разных сторонних источников ацетилхолин.
При денервации в мышце и других тканях появляются свойства, присущие ранним, в частности, эмбриональным стадиям развития. Это явление возникает как результат патологического растормаживания супрессированных в норме генов. Деафферентация Импульсация, поступающая в нейрон, из какого бы источника она ни исходила, является для нейрона афферентной. Выключение этой афферентации представляет собой деафферентацию нейрона. Последняя может быть обусловлена либо выпадением поступающей импульсации (при перерыве нервных путей, нарушении выделения нейромедиаторов пресинаптическими окончаниями), либо блокадой воспринимающих рецепторов на постсинаптическом нейроне (при действии токсинов, фармакологических средств и др.). Многие явления при деафферентации нейрона представляют собой выражение денервационного синдрома. Полной деафферентации нейрона не происходит, так как нейроны ЦНС обладают огромным количеством афферентных входов. Тем не менее и при частичной деафферентации возникает повышение возбудимости нейрона или его отдельных участков и нарушение тормозных механизмов. В силу этого при деафферентации группа нейронов может приобрести свойства ГПУВ. В клинике под феноменом деафферентации имеют в виду синдромы, возникающие при выпадении афферентной стимуляции с периферии. Эти синдромы можно воспроизвести в эксперименте путем перерезки соответствующих задних корешков спинного мозга. Движения конечности, иннервируемой деафферентированными таким образом сегментами спинного мозга, становятся размашистыми, плохо координированными. Кроме того, такая конечность способна осуществлять спонтанные движения в такт с дыханием (феномен Орбели-Кунстман), что обусловлено растормаживанием и повышением возбудимости деафферентированных нейронов спинного мозга.
Спинальный шок Спинальный шок возникает после перерыва спинного мозга и представляет собой глубокое, но обратимое угнетение двигательных и вегетативных рефлексов, осуществляющихся ниже перерыва. Угнетение рефлексов связано с выпадением активирующей стимуляции со стороны головного мозга. У лягушек, у которых зависимость спинного мозга от головного значительно меньше, чем у высших животных, спинальный шок длится несколько минут, у человекообразных обезьян и человека - несколько месяцев. У человека полная арефлексия после перерыва спинного мозга является начальной стадией полной параплегии. В дальнейшем происходит постепенное восстановление двигательных и вегетативных рефлексов. Вначале появляются сгибательные рефлексы пальцев, имеющие характер патологических рефлексов (рефлекс Бабинского и др.), после этого осуществляются более значительные и затем генерализованные спинальные рефлексы и движения типа спинальных автоматизмов. Нарушение нервной трофики. Нейродистрофический процесс Трофика клетки и дистрофический процесс. Трофика клетки - комплекс процессов, обеспечивающих ее жизнедеятельность и поддержание генетически заложенных свойств. Расстройство трофики представляет собой дистрофию, развивающиеся дистрофические изменения составляют дистрофический процесс. Нейродистрофический процесс. Это развивающееся нарушение трофики, которое обусловлено выпадением или изменением нервных влияний. Оно может возникать как в периферических тканях, так и в самой нервной системе. Выпадение нервных влияний заключается: 1) в прекращении стимуляции иннервируемой структуры в связи с нарушением выделения или действия нейромедиатора; 2) в нарушении секреции или действия комедиаторов - веществ, которые выделяются вместе с нейромедиаторами и играют роль нейромодуляторов, обеспечивающих регуляцию рецепторных, мембранных и метаболических процессов; 3) в нарушении выделения и действия трофогенов. Трофогены (трофины) - вещества различной, преимущественно белковой природы, осуществляющие собственно трофические эффекты поддержания жизнедеятельности и генетически заложенных свойств клетки. Источником трофогенов являются: 1) нейроны, из которых трофогены поступают с антероградным (ортоградным) аксоплазматическим током в клетки-реципиенты (другие нейроны или иннервируемые ткани на периферии); 2) клетки периферических тканей, из которых трофогены поступают по нервам с ретроградным аксоплазматическим током в нейроны ; 3) глиальные и шванновские клетки, которые обмениваются с нейронами и их отростками трофическими веществами. Вещества, играющие роль трофогенов, образуются также из сывороточных и иммунных белков. Трофическое воздействие могут оказывать некоторые гормоны. В регуляции трофических процессов принимают участие пептиды, ганглиозиды, некоторые нейромедиаторы.
К нормотрофогенам относятся различного рода белки, способствующие росту, дифференцировке и выживанию нейронов и соматических клеток, сохранению их структурного гомеостаза (например, фактор роста нервов). 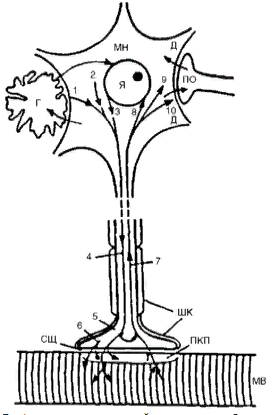 Трофические связи мотонейрона и мышцы. Вещества из тела мотонейрона (МН), его мембраны 1, перикариона 2, ядра 3 транспортируются с антероградным аксоплазматическим током 4 в терминаль 5. Отсюда они, а также вещества, синтезируемые в самой терминали 6, поступают транссинаптически через синаптическую щель (СЩ) в концевую пластинку (КП) и в мышечное волокно (МВ). Часть неиспользованного материала поступает обратно из терминали в тело нейрона с ретроградным аксоплазматическим током 7. Вещества, образующиеся в мышечном волокне и концевой пластинке, поступают транссинаптически в обратном направлении в терминаль и далее с ретроградным аксоплазматическим током 7 в тело нейрона - к ядру 8, в перикарион 9, к мембране дендритов 10. Некоторые из этих веществ могут поступать из дендритов (Д) транссинаптически в другой нейрон через его пресинаптическое окончание (ПО) и из этого нейрона далее в другие нейроны. Между нейроном и мышцей происходит постоянный обмен веществами, поддерживающими трофику, структурную целостность и нормальную деятельность обоих образований. В этом обмене принимают участие глиальные клетки (Г). Все указанные образования создают регионарную трофическую систему (или трофический контур) изменения клеток-реципиентов (патотрофогены, по Г.Н. Крыжановскому). Такие вещества синтезируются, например в эпилептических нейронах - поступая с аксоплазматическим током в другие нейроны, они могут индуцировать у этих нейронов-реципиентов эпилептические свойства. Патотрофогены могут распространяться по нервной системе, как по трофической сети, что является одним из механизмов распространения патологического процесса. Патотрофогены образуются и в других тканях |