норм физ. Лекции наука о механизмах жизнедеятельности организма и его взаимодействия с окружающей средой
 Скачать 1.71 Mb. Скачать 1.71 Mb.
|

|
Вопрос 16 1. Препотенциал - локальный потенциал, возникающий при действии субпороговых раздражителей в тех же участках мембраны, где и потенциал действия, т.е. имеющих потенциалуправляемые каналы. Препотенциал расположен в субпороговой области (между МПП и критическим уровнем деполяризации), имеет фазу деполяризации и реполяризации. 2. Механизмы возникновения препотенциала (локального ответа). При действии субпорогового раздражителя сначала возникает чисто физическая деполяризация мембраны, связанная с сопротивлением и ёмкостью мембраны. Затем возникает добавочная деполяризация, связанная с открытием потенциалуправляемых Na+-каналов и входящим в клетку Na+-током (именно эта деполяризация и называется препотенциалом). Деполяризация способствует открытию также и более медленных потенциалзависимых К+-каналов, что увеличивает выходящий из клетки К+-ток и вызывает затем фазу реполяризации. Во время препотенциала входящий в клетку Na+-ток меньше, чем выходящий из клетки К+-ток через потенциалуправляемые каналы и К+-каналы утечки, поэтому после прекращения действия субпорогового раздражителя препотенциал исчезает. 3. Свойства препотенциала. Амплитуда препотенциала находится в прямой зависимости от силы раздражителя, он возникает в соответствии с законом “силы”. Препотенциалы способны к суммации (временной), если промежутки между раздражителями короче, чем время существования препотенциала. Высокочастнотные субпороговые раздражители могут деполяризовать мембрану до критического уровня и вызвать ПД. Во время препотенциала повышена возбудимость. Распространение препотенциала происходит с затуханием амплитуды на небольшие расстояния. 4. Критический уровень деполяризации (КУД, Екр) - это тот уровень, при котором деполяризация мембраны принимает регенеративный (самоусиливающийся) характер, свидетельствующий о развитии ПД. При этом входящий в клетку Na+-ток равен выходящему из клетки К+-току, что характеризует электрическую нестабильность мембраны, процесс может идти как в сторону деполяризации и образования ПД, так и в сторону реполяризации и ограничиться препотенциалом. Пороговый раздражитель - раздражитель, деполяризующий МПП до КУД. Пороговый потенциал - величина потенциала, равная разности МПП и КУД, он характеризует возбудимость клетки (чем больше ПП, тем больше возбудимость). Вопрос 17 . Возбудимость - способность специализированной ткани ( нервной, мышечной, железистой) отвечать процессом возбуждения на действие раздражителей. Реобаза – это минимальная сила раздражителя, вызывающая возбуждение при его действии в течение не-ограниченно долгого времени. На практике порог и реобаза имеют одинаковый смысл. Чем ниже порог раз-дражения или меньше реобаза, тем выше возбудимость ткани. Хронаксия – это минимальное время действия раздражителя силой в две реобазы, необходимое для возник-новения возбуждения. Этот параметр предложил рассчитывать Л.Лапик, для более точного определения по-казателя времени на кривой силы-длительности. Чем короче полезное время или хронаксия, тем выше возбу-димость, и наоборот. Использование хронаксиметрии в физиологии и медицине. Хронаксиметрия — один из методов диагностики функционального состояния нервов и мышц с помощью постоянного электрического тока. Для характеристики возбудимости ткани необходимо определить мини-мальный порог раздражения (реобазу) и минимальное время, в течение которого ток, по силе или по напря-жению равный удвоенной реобазе, должен возбудить ткань (хронаксия).В медицинской практике чаще всего определяется хронаксия мышц и двигательных нервов. Исследуется также хронаксия и чувствительной сфе-ры. Хронаксия скелетных мышц человека колеблется от 0,1 до 0,7 мс. Хронаксия сгибателей у человека в 1,5-2 раза меньше хронаксии разгибателей. Для оценки возбудимости ткани обычно используются такие критерии, как пороговая сила и хронак-сия. Пороговая сила — это наименьшая сила раздражителя, способная вызвать возбуждение (ПД) при неограни-чении ее действия во времени (рис. 1.5 - проекция точки А на ординату). Сила раздражителя - понятие со-бирательное, оно отражает степень выраженности раздражающего воздействия стимула на ткань. Например, сила электрического тока выражается в амперах (А), температура среды - в градусах (С°), кон-центрация химического вещества - в ммоль/л, сила звука - в децибелах (дБ) и т.д. При использовании в каче-стве раздражителя электрического тока предложенное определение пороговой силы совпадает с понятием «реобаза». Если возбудимость ткани высока, пороговая сила раздражителя мала. Чем выше возбудимость, тем ниже пороговая сила. Большая пороговая сила свидетельствует о низкой возбудимости ткани. При внут-риклеточном раздражении пороговая сила электрического тока для различных клеток равна Ю-7 -Ю-9 А". При медленно нарастающей силе раздражителя возбуждение может не возникнуть даже при достижении боль-шой его силы, значительно превосходящей пороговую, вследствие развития аккомодации (снижения возбу-димости и ПД вплоть до полного его отсутствия при медленно нарастающем стимуле, так как развивается инактивация №-каналов). Важным условием, обеспечивающим возникновение возбуждения при действии раздражителя, является его длительность. Пороговое время — наименьшее время, в течение которого должен действовать раздражитель пороговой силы, чтобы вызвать возбуждение. Чем меньше пороговое время, тем выше возбудимость ткани. Вопрос 18 Потенциал действия - обязательный электрический компонент возбуждения нервных и мышечных клеток, представляющий собой быстрое и высокоамплитудное колебание их мембранного потенциала, часто сопро-вождающееся перезарядкой мембраны, ПД возникает в электровозбудимых участках мембраны при действии раздражителей пороговой и сверхпороговой силы. Схема ПД . В ПД различают высокоамплитудную кратковременную часть — спайк (пик), и более длитель-ную низкоамплитудную часть - следовые потенциалы: следовую деполяризацию и следовую гиперполяри-зацию (они могут быть в разной последовательности и длительности). Спайк состоит из деполяризации мембраны, овершута (пеpескока через нулевой уровень) с переменой заряда мембраны (имеется не во всех ПД), и реполяризации мембраны. Амплитуда ПД от 30 мВ в клетках гладких мышц до 120 мВ в миокарде, длительность от 1 мс в нервных волокнах до 300-500 мс в клетках миокарда. Восходящая фаза ПД состоит из деполяризации мембраны и её перезарядки если есть овершут. Деполяризация мембраны, вызванная раздражителем и достигшая КУД, открывает быстрые m-ворота в по-тенциал управляемых Na+-каналах и образует мощный входящий Na+-ток (до 6 МА/см в кв.), который при-водит к увеличению деполяризации мембраны. Деполяризация ПД при действии как пороговых, так и сверхпороговых раздражителей. Самоусиление можно представить следующим образом: деполяризация- открытие Na+-каналов--входщд Na+ в клетку--еще большая Деполяризация--еще большее открыте Na+-каналов и тд. быстрые т-ворота в потенциалуправляемых Na"-каналах и образует мощный входящий есть овершут). Входящий Na+-mок не только доводит МПП до нуля, но может вызывать перезарядку мембраны (примерно до +20 мB). Продолжающаяся деполяризация мембраны вызывает инактивацию Na+-каналов (закрытие в них медленных һ-ворот), поэтому пик ПД не достигает величины равновесного Na+-потенциала, равной примерно +60 мB. Подчеркивая выдающуюся роль Na+ в развитии ПД, его иногда кратко называют натрие-вым потенциалом, однако в некоторых клетках и участках нейрона возникают кальциевые ПД. Нисходящая фаза ПД состоит из ликвидации перезарядки мембраны (овершута) и реполяризация мембра-ны. Механизмы реполяризации. Деполяризация мембраны в предыдущей фазе открывает потенциалуправляемые К+-каналы. Реакция ворот этих каналов более медленная, чем ворот в Na+-каналах, ответственных за восхо-дящую фазу. Поэтому, хотя активация К+-каналов начинается почти одновременно с Na+-каналами, реально повышение К+-тока развивается несколько позже, и только после вершины спайка этот ток начинает преоб-ладать над входящим Na+-током, ликвидируя перезарядку мембраны (овершут) и восстанавливая поляриза-цию мембраны. Ликвидация инактивации Na+-каналов и восстановление возбудимости клетки. На протяжении нисходящей фазы ПД Na+-ток быстро уменьшается поскольку реполяризация закрывает быстрые активационные ворота в Na+-каналах, при этом инактивацинные ворота открываются, но более медленно, Хотя Na+-канал закрыт, но открытые инактивационные ворота в нем делают клетку в конце фазы реполяризапии способной ответить на действие сверхпороговых раздражителей. Выходящий К+-ток в процессе реполяризации также несколько снижается (но более мелленно чем Na+-ток), он остается повышенным и после возврата MП к уровню покоя. Следовые потенциалы. После высоковольтной части (спайка) ПД развиваются в разной последовательности и степени выраженности следовые потенциалы, которые длительно (десятки и сотни миллисекунд) изменя-ют возбудимость клетки после спайка ПД. Следовая деполяризация связана с резким снижением повышенного К+-тока по механизму отрицательной обратной связи и отражает снижение скорости реполяризации при приближении MП к уровню покоя. В это время возбудимость клетки повышена, так как пороговый потенциал (расстояние между МП и КУД) умень-шен. Следовая гиперполяризация связана с длительным небольшим избытком К+-тока и асимметричной (электрогенной) деятельностью Na/ K+-насоса, активированного входом Na+ в клетку в фазе деполяризации. Возбудимость клетки в это время снижена, так как пороговый потенциал увеличен. Восстановление ионных градиентов в клетке после потенциала действия. Экономичность ПД. Потенциал действия сопровождается небольшим нарушением ионных градиентов клет-ки. Было рассчитано, что во время ПД в клетку входят около 19000 Na+ через 1 мкм в кв. поверхности мем-браны, почти столько же выходит К+. Поэтому ПД существенно не нарушает ионные градиенты клетки, и она может без «подзарядки» генерировать десятки тысяч ПД. Роль Na+/К+ -насоса. В реальных условиях сохранение ионных градиентов клетки даже в условиях интен-сивной импульсной активности легко обеспечивается работой К+/Na+-насоса как во время ПД, так и особен-но в промежутках между ПД. Однако возбужденная клетка вследствие небольшого объема прилегающей межклеточной жидкости создает вокруг себя зону повышенной концентрации К+, влияющей как на эту клетку, так и на расположенные рядом сосуды. Функциональная роль потенциала действия. С помощью ПД в организме кодируется громадное количе-ство информации путем изменения частоты образования ПД и изменения временных промежутков между ПД (частотно-спектральный код). ПД, в отличие oт других потенциалов, может распространиться на боль-шие расстояния (у человека до 1м ). Поэтому в нервной системе ПД являются основным способом получе-ния и передачи информации, выработки в нервных центрах командных кодов, с помощью которых они регу-лируют деятельность организма. В мышечных клетках ПД осуществляют запуск их сокращения, в желези-стых - выработку секрета. Вопрос 19 19.Изменения возбудимости при возбуждении. Фазы возбудимости и их соотношение с фазами потенциала действия. Рефрактерность, её механизмы. Рис. 1 Соотношение фаз возбудимости с фазами потенциала действия 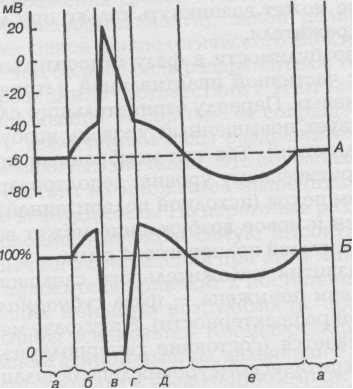 Рис. 2 Соотношение фаз возбудимости с фазами потенциала действия (другой вид)  Для А: а — мембранный потенциал покоя; б — локальный ответ или ВПСП; в — восходящая фаза потенциала действия (деполяризация и инверсия); г — нисходящая фаза потенциала действия (реполяризация); д — отрицательный следовой потенциал (следовая деполяризация); е — положительный следовой потенциал (следовая гиперполяризация) Для Б; а — исходный уровень возбудимости; б — фаза повышенной возбудимости; в — фаза абсолютной рефрактерности; г — фаза относительной рефрактерности; д — фаза супернормальной возбудимости; е — фаза субнормальной возбудимости При развитии потенциала действия происходят фазные изменения возбудимости ткани состоянию исходной поляризации мембраны (мембранный потенциал покоя) соответствует нормальный уровень возбудимости. В период предспайка возбудимость ткани повышена. Эта фаза возбудимости получила название повышенной возбудимости (первичной экзальтации). В это время мембранный потенциал приближается к критическому уровню деполяризации, поэтому дополнительный стимул, даже если он меньше порогового, может довести мембрану до критического уровня деполяризации. В период развития спайка (пикового потенциала) идет лавинообразное поступление ионов натрия внутрь клетки, в результате чего происходит перезарядка мембраны и она утрачивает способность отвечать возбуждением на раздражители даже сверхпороговой силы. Эта фаза возбудимости получила название абсолютной рефрактерности (абсолютной невозбудимости). Она длится до конца перезарядки мембраны и возникает в связи с тем, что натриевые каналы инактивируются. После окончания фазы перезарядки мембраны возбудимость ее постепенно восстанавливается до исходного уровня — фаза относительной рефрактерности. Она продолжается до восстановления заряда мембраны, достигая величины критического уровня деполяризации. Так как в этот период мембранный потенциал покоя еще не восстановлен, то возбудимость ткани понижена и новое возбуждение может возникнуть только при действии сверхпорогового раздражителя. Снижение возбудимости в фазу относительной рефрактерности связано с частичной инактивацией натриевых каналов и активацией калиевых. Периоду отрицательного следового потенциала соответствует повышенный уровень возбудимости (фаза вторичной экзальтации). Так как мембранный потенциал в эту фазу ближе к критическому уровню деполяризации по сравнению с состоянием покоя (исходной поляризацией), то порог раздражения снижен и новое возбуждение может возникнуть при действии раздражителей подпороговой силы. В период развития положительного следового потенциала возбудимость ткани понижена — фаза субнормальной возбудимости (вторичной рефрактерности). В эту фазу мембранный потенциал увеличивается (состояние гиперполяризации мембраны), удаляясь от критического уровня деполяризации, порог раздражения повышается и новое возбуждение может возникнуть только при действии раздражителей сверхпороговой величины. Рефрактерность После окончания возбуждения в нервных или мышечных клетках или, другими словами, после окончания в них потенциала действия наступает временное состояние невозбудимости – рефрактерности. После сокращения сердца очередное сокращение нельзя было вызвать в течении периода, равного десятым долям секунды независимо от амплитуды и длительности раздражающего стимула. В нервных клетках период невозбудимости оказался значительно короче. При уменьшении интервала раздражения между двумя раздражающими электрическими стимулами величина потенциала действия в ответ на второй стимул становится все меньше и меньше. А если повторный стимул наносится во время генерации потенциала действия или сразу же после его окончания, второй потенциал действия не генерируется. Период, в течении которого, потенциал действия на второй раздражающий стимул не возникает, получил название абсолютного рефрактерного периода. Он составляет для нервных клеток позвоночных животных 1,5 – 2 мс. После периода абсолютной рефрактерности наступает относительный рефрактерный период. Он характеризуется: 1) повышенным порогом раздражения по сравнению с исходным состоянием (т.е. для того чтобы возник повторный потенциал действия, необходим ток большей величины) 2) снижением амплитуды потенциала действия. По мере окончания периода относительной рефрактерности возбудимость повышается до исходного уровня, и величина порогового раздражения уменьшается также до первоначального значения. В период абсолютной рефрактерности наблюдается повышенная калиевая проводимость за счёт открывания дополнительных калиевых каналов и снижение натриевой проводимости за счёт инактивации натриевых каналов. Поэтому даже при больших значениях деполяризующего тока не удаётся активировать такое количество натриевых каналов, чтобы выходящий натриевый ток мог бы превысить увеличенный выходящий калиевый ток и снова запустить регенеративный процесс. Во время относительного рефрактерного периода деполяризующий сигнал достаточно большой амплитуды может активировать воротный механизм натриевых каналов так, что несмотря на большое число открытых калиевых каналов натриевая проводимость увеличивается и вновь возникает потенциал действия. Вместе с тем из-за увеличенной проводимости мембраны к ионам калия и остаточной натриевой инактивации повышение мембранного потенциала не будет уже столь близко к значению равновесного натриевого потенциала. Поэтому потенциал действия будет меньшим по амплитуде. Далее следует фаза экзальтации – повышенной возбудимости возникающей в результате, наличия следовой деполяризации. В последующем при развитии следовой гиперполяризации наступает фаза субнормальности – характеризующаяся снижением амплитуды потенциалов действия. Наличие рефрактерных фаз обуславливает прерывистый (дискретный) характер нервной сигнализации, а ионный механизм генерации потенциала действия обеспечивает стандартность нервных импульсов. Вследствие этого изменения внешних сигналов кодируются изменением частоты потенциалов действия. Максимально возможный ритм активности, лимитированный длительностью абсолютной рефрактерной фазы обозначают как лабильность (функциональную подвижность). У нервных волокон лабильность составляет 200 - 400 Гц, а у некоторых чувствительных нервных волокон достигает 1кГц. В случае, когда новый раздражающий импульс приходится на фазу экзальтации реакция ткани становится максимальной – развивается оптимум частоты. При попадании последующего стимулирующего импульса на фазу относительной или абсолютной рефрактерности реакция ткани ослабляется или прекращается вовсе, развивается пессимальное торможение. |