Курс лекций по биохимии. Лекция Введение в дисциплину
 Скачать 0.7 Mb. Скачать 0.7 Mb.
|

4. Брожение и его основные типыОбразовавшаяся в результате гликолиза ПВК является неустойчивым химическим соединением и всегда подвергается дальнейшим преобразованиям. Пути метаболизации ПВК зависят от того, есть ли в среде кислород, или условия являются анаэробными. В присутствии кислорода метаболизация ПВК осуществляется путем ее окислительного декарбоксилирования, в результате чего образуется ацетилкоэнзим А. При отсутствии кислорода метаболизация ПВК осуществляется путем брожения.Брожением называются процессы превращения пировиноградной кислоты в анаэробных условиях в устойчивые конечные продукты без дополнительного высвобождения энергии.Поскольку свободный кислород, имеющийся на нашей планете, образовался в процессе фотосинтеза, возникшего на более поздних этапах развития жизни на Земле, совершенно очевидно, что анаэробный распад углеводов, т.е. процесс брожения, является более древним типом катаболизма, чем процесс дыхания. На это указывает также то, что по сравнению с дыханием брожение–процесс, энергетически значительно менее выгодный, поскольку для получения одного и того же количества энергии при брожении расходуется значительно больше сахара, чем при дыхании.Тип брожения определяется спецификой организма и наличием соответствующих ферментных систем. Спиртовое брожение При спиртовом брожении на 1-м этапе происходит декарбоксилирование ПВК и превращение ее в уксусный альдегид, а затем образуется этиловый спирт.Спиртовое брожение характерно для дрожжей. Среди них наибольшее значение имеют истинные дрожжи – организмы, принадлежащие к роду Saccharomyces. К числу организмов, вызывающих при определенных условиях спиртовое брожение, принадлежат и так называемые дрожжеподобные организмы (Monilia, Oidium), а также некоторые из плесневых грибов, например, Mucor. Спиртовое брожение находит применение в пивоварении, виноделии и для приготовления дрожжевого теста. При спиртовом брожении всегда в незначительных количествах образуются янтарная кислота, уксусный альдегид, глицерин и так называемые сивушные масла – смесь амилового, изоамилового, бутилового и других спиртов. Наиболее легко подвергаются сбраживанию глюкоза и фруктоза, медленнее – манноза и еще медленнее – галактоза. Пентозы дрожжами не сбраживаются; они могут сбраживаться только некоторыми плесневыми грибами из рода Fusarium. Из дисахаридов хорошим субстратом спиртового брожения являются сахароза и мальтоза. Однако оба этих сахара сбраживаются лишь после предварительного гидролиза на составляющие их моносахариды. В присутствии кислорода спиртовое брожение прекращается и дрожжи получают энергию, необходимую для их развития и жизнедеятельности, путем кислородного дыхания. При этом дрожжи тратят сахар значительно экономнее, чем в анаэробных условиях. Близкой к спиртовому брожению формой анаэробного обмена является уксуснокислое брожение, характерное для некоторых бактерий; в результате такого брожения образуется уксусная кислота. Молочнокислое брожениеМолочнокислое брожение характерно для некоторых видов бактерий и используется для приготовления молочнокислых продуктов (простокваши, ацидофилина, кефира), при изготовлении кваса, хлебных заквасок и «жидких дрожжей» для хлебопечения, при квашении капусты, огурцов, при силосовании кормов. Все микроорганизмы, вызывающие молочнокислое брожение, разделяются на две большие группы. К первой группе принадлежат микроорганизмы, являющиеся настоящими (облигатными) анаэробами и сбраживающие гексозы с образованием молочной кислоты. Эти микроорганизмы получили название гомоферментативных молочнокислых бактерий. Ко второй группе (гетероферментативные молочнокислые бактерии) относятся микроорганизмы, которые кроме молочной кислоты образуют значительные количества других продуктов, в частности уксусной кислоты и этилового спирта. Заметное содержание молочной и уксусной кислот в ржаном тесте и ржаном хлебе объясняется тем, что здесь наряду со спиртовым брожением происходит также молочнокислое брожение. Одновременное протекание в ржаном тесте этих процессов объясняется присутствием в закваске как дрожжей, вызывающих спиртовое брожение, так и молочнокислых бактерий, вызывающих молочнокислое брожение. Подобного рода совместное существование дрожжей и молочнокислых бактерий, оказывающих друг на друга благотворное влияние, наблюдается в целом ряде пищевых продуктов и полуфабрикатов – в хлебном квасе, кумысе, различных кисломолочных продуктах. Молочнокислое брожение у аэробных организмов У многоклеточных организмов образование молочной кислоты может происходить в различных тканях в условиях кислородного голодания. К примеру, у человека молочнокислое брожение идет в мышцах при недостатке кислорода. Сердце при нагрузке на мышцы не успевает их обеспечить кислородом, вследствие чего накапливается лактат (молочная кислота) и мышцы болят. При недостатке кислорода электроны, образующиеся при окислении НАД Н, восстанавливают все белки-переносчики ЭТЦ; в результате новые порции НАД Н не окисляются и НАД восстановленный накапливается; в результате возникает недостаток НАД+ окисленного. НАД окисленный является коферментом дегидрогеназы, участвующей в цикле Кребса и в превращении ПВК в ацетил КоА. В его отсутствие эти дегидрогеназы не активны, т.е. ПВК не может превращаться в ацетил КоА и цикл Кребса блокируется. Таким образом, роль молочнокислого брожения состоит в окислении НАД Н, образующегося при гликолизе, что позволяет гликолизу функционировать. В результате образуется 2 АТФ, поддерживающие некоторое время жизнедеятельность организма. Маслянокислое брожениеБольшинство микроорганизмов, вызывающих маслянокислое брожение, являются анаэробами. Некоторые из них принадлежат к группе облигатных анаэробов, т.е. таких организмов, которые могут жить только в отсутствие кислорода, и для которых последний является ядом. Наряду с масляной кислотой, двуокисью углерода и водородом при маслянокислом брожении образуются этиловый спирт, а также молочная и уксусная кислоты. Маслянокислое брожение в природных условиях происходит в гигантских масштабах на дне болот, в заболоченных почвах, в различного рода илах и всех тех местах, куда ограничен доступ кислорода, и где в результате деятельности маслянокислых бактерий разлагаются огромные количества органического вещества. Существуют и другие схемы брожения, при которых конечными продуктами могут являться уксусная, валериановая, пропионовая а также другие органические кислоты. Распад органического вещества в процесс гликолиза и брожения является не только источником энергии для данного организма, но также источником различных соединений, возникающих в качестве промежуточных продуктов брожения и используемых организмом для многочисленных синтетических реакций. Лекция 8. Аэробное дыхание1. Основные этапы аэробного дыхания Как было отмечено ранее, образовавшаяся в результате гликолиза пировиноградная кислота (пируват) является неустойчивым химическим соединением и всегда подвергается дальнейшей метаболизации. В большинстве случаев (в аэробных условиях) ПВК подвергается полному окислению до диоксида углерода и воды при участии кислорода с полным извлечением энергии и запасанием ее в форме АТФ. Этот процесс называется аэробным дыханием. Аэробное дыхание представляет собой серию достаточно сложных процессов, которые можно разбить на 3 стадии: 1) Окислительное декарбоксилирование пирувата (ПВК) с образованием ацетилкоэнзима А (либо образование его из жирных кислот и аминокислот). 2) Разрушение ацетильных групп в цикле трикарбоновых кисло (цикл Кребса) с высвобождением углекислого газа и атомов водорода. 3) перенос электронов (соответствующих этим атомам водорода к молекулярному кислороду – процесс окисления, сопряженный с фосфорилированием АДФ, в результате которого образуется АТФ). 2. Окислительное декарбоксилирование пирувата (ПВК) Гликолиз, в результате которого образуется пировиноградная кислота, осуществляется в цитоплазме клеток. Затем ПВК попадает в митохондрии клеток. Там она окисляется (в аэробных условиях) и превращается в богатое энергией производное уксусной кислоты – ацетилкоэнзим А. Реакция ускоряется мультиэнзимным комплексом, который называется пируватдегидрогеназным комплексом. 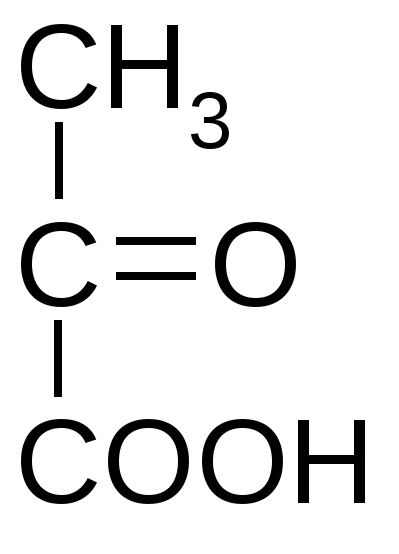 → → → → → → 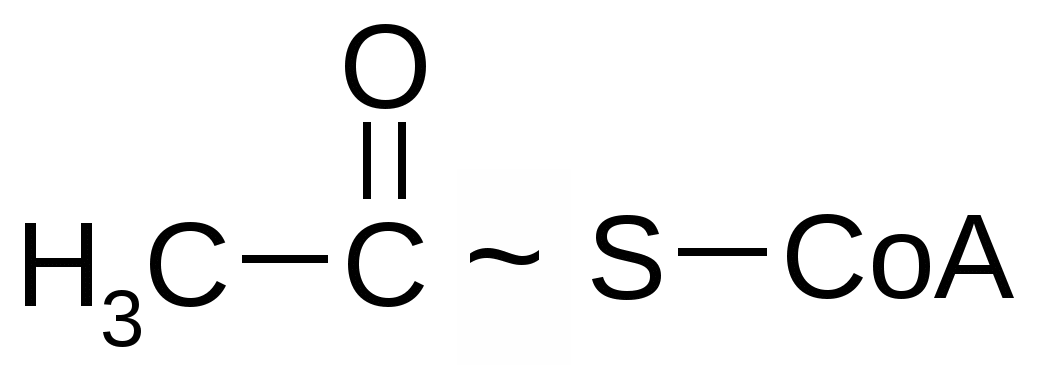 ПВК Ацетилкоэнзим А Процесс этот можно выразить следующим суммарным уравнением: СН3-СО-СООН + НАД+ +НSСоА →СН3-СО-SСоА + СО2 ↑+ НАДН+Н+ ПВК Ацетилкоэнзим А Пируватдегидрогеназный комплекс, катализирующий процесс окислительного декарбоксилирования ПВК, представляет собой мульферментную систему, включающую три фермента, каждый из которых катализирует определенную стадию окислительного декарбоксилированния. Этот процесс идет при участии 5 кофакторов (НАД, ФАД, коэнзим А, тиаминпирофосфат, амид липоевой кислоты), в работе которых участвует 5 витаминов. Следует отметить, что в результате реакции окисления ПВК в образующейся молекуле ацетилкоэнзима А возникают макроэргические связи, которые способствуют его энергетическому обмену в дальнейшем.3. Цикл Кребса (цикл ди- и трикарбоновых кислот, цикл лимонной кислоты)Биологический смысл цикла заключается в расщеплении пировиноградной кислоты до двуокиси углерода и извлечение из нее энергии. Все ферменты цикла Кребса локализованы в митохондриях, в их внутреннем межмембранном пространстве, которое заполнено матриксом – полужидким белковым веществом. На начальном этапе цикла Кребса ацетильный радикал ацетил-КоА соединяется с молекулой щавелевоуксусной кислоты, и при этом образуется трикарбоновая лимонная кислота. Лимонная кислота окисляется в ходе последующих четырех ферментативных реакций. При этом восстанавливаются 3 молекулы НАД+ в НАД*Н, 1 молекула ФАД в ФАД*Н2, и образуется одна молекула гуанозинтрифосфата (ГТФ) с макроэргической фосфатной связью. Энергия ГТФ используется для фосфорилирования АДФ и образования АТФ. Лимонная кислота теряет 2 углеродных атома, за счет которых образуются 2 молекулы СО2. В итоге, в результате семи последовательных реакций, лимонная кислота превращается в щавелевоуксусную кислоту. Образовавшаяся молекула щавелевоуксусной кислоты соединяется с новой молекулой ацетилкоэнзима А, поступающей на этот циклический конвейер ферментов. При этом вновь образуется молекула лимонной кислоты, которая ступенчато окисляется до щавелевоуксусной кислоты, и цикл вновь повторяется. В составе лимонной кислоты как бы сгорает присоединившийся остаток ацетил-КоА. При этом образуется углекислый газ, атомы водорода и электроны переносятся на акцепторы – НАД и ФАД окисленные. Таким образом, энергия химических связей органических веществ (углеводов, жиров и белков) накапливается в молекулах НАД*Н, ФАД*Н2 и АТФ. Непосредственно в ходе цикла Кребса при окислении 2-х молекул ацетил КоА синтезируется только 2 молекулы ГТФ. Синтез остальных молекул АТФ происходит при перемещении электронов по цепи переноса, которое осуществляется при участии комплексов ферментов, сосредоточенных на внутренней мембране митохондрий (так называемое окислительное фосфорилирование). Таким образом, реакции цикла Кребса выполняют следующие функции: 1) Энергетическую. 2) Интегративную (объединяют распад углеводов, белков и липидов). 3) Пластическую (кислоты цикла Кребса являются важными метаболитами, участвующими во многих биохимических реакциях). 4. Окислительное фосфорилирование Окислительное фосфорилирование – фосфорилирование АДФ с образованием АТФ, сопряженное с переносом электронов от восстановителей (НАД Н, ФАД Н2) по электроннотранспортной цепи митохондрий к кислороду. Процесс называется окислительным, поскольку окисляются восстановленные динуклеотиды, и за счет этого появляется АТФ. Электроны от НАД*Н и ФАД*Н2 перемещаются по многоступенчатой цепи переноса электронов к конечному их акцептору – молекулярному кислороду. Схема окислительного фосфорилирования была предложена в 1961 году Митчеллом и получила название «хемиосмотическая теория». Электронно-транспортная цепь (ЭТЦ), или цепь переноса электронов, расположена на внутренней мембране митохондрий. В ее состав входит ряд последовательно расположенных переносчиков электронов, которые отличаются способностью акцептировать электроны. Самый сильный акцептор электронов – кислород расположен в конце цепи. Среди промежуточных переносчиков электронов – кофермент Q, цитохромы b, c1, c, a, a3 и, наконец, О2. Кофермент Q, или убихинон, представляет собой производное бензохинона, способное при участии фермента принимать электроны окисляемого НАД*Н и передавать их на восстанавливаемый цитохром b. Промежуточным переносчиком электронов является фермент, в состав которого входит акцептор электронов – флавинмононуклеотид (ФМН). При переносе электрона с ФМН на кофермент Q происходит первое фосфорилирование АДФ в АТФ. Еще две молекулы АТФ синтезируются на последующих этапах, где акцепторами электронов служат цитохромы. Цитохромы представляют собой белки, к которым присоединены молекулы гема железопорфирина, подобные тем, которые содержатся в составе гемоглобина. В комплексе а и а3, который называют цитохромоксидазой, помимо атомов железа содержатся атомы меди. Атомы железа и меди в этих молекулах и служат попеременно акцепторами и донорами электронов. Как энергия, запасенная в НАД*Н и ФАД*Н2, служит синтезу АТФ? Атомы водорода этих восстановленных соединений транспортируются в мембраны митохондрий, где они оставляют электроны на внутренней стороне мембраны, а катионы Н+ с помощью белковой молекулы переносятся на наружную сторону (энергия электронов НАД Н затрачивается на перенос 2Н+ через мембрану). Поскольку внутренняя мембрана непроницаема для протонов, то они накапливаются с наружной стороны внутренней мембраны. Электроны соединяются с кислородом, в результате чего образуются анионы. Между наружной поверхностью мембраны, где накапливаются катионы Н+, и внутренней, где накапливаются анионы О2-, возникает разность потенциалов. Формируется электрохимический потенциал – особая форма энергии. В мембраны митохондрий встроены молекулы фермента, синтезирующего АТФ (АТФ-аза). Когда разность потенциалов достигает определенной величины, эта энергия затрачивается на изменение конформации АТФ-азы; при этом канал АТФ-азы открывается и по нему протоны возвращаются в матрикс, а в головке АТФ-азы происходит: АДФ +Фн = АТФ Внутри митохондрий катионы Н+, соединяясь с анионами О2-, образуют воду. Многие яды, например, динитрофенол, являются разобщителями сопряженного окислительного фосфорилирования: они открывают канал и протоны свободно возвращаются в матрикс, АТФ не синтезируется и человек погибает. Таким образом, в цикле трикарбоновых кислот образуется углекислый газ и в цепи переноса электронов – вода. Это те же конечные продукты, которые образуются при сжигании органического топлива. Энергетический баланс цикла Кребса ФАД Н2 = 2 АТФ 4 НАД Н =12 АТФ НАД Н2=3 АТФ ФАД = 2 АТФ ГТФ = АТФ ГТФ=АТФ Итого: 15 АТФ (на 1 молекулу ПВК) У аэробов в процессе гликолиза (расщепления глюкозы до 2-х молекул ПВК) образуется 8 молекул АТФ. Таким образом, суммарный энергетический баланс полного окисления 1-й молекулы глюкозы до воды и углекислого газа составляет: 8 АТФ + 2*15 АТФ = 38 АТФ Не только глюкоза может служить источником энергии. Окисляются в клетках и жирные кислоты, которые образуются в результате ферментативного расщепления жиров липазой. В результате окисления жирных кислот в конечном итоге также образуется ацетил-КоА и восстанавливаются акцепторы электронов НАД+ в НАД*Н. При этом происходит восстановление акцепторов еще одного типа – ФАД в ФАД*Н2 Энергия, запасенная в цикле трикарбоновых кислот, в молекулах НАД*Н и ФАД*Н2, также используется дальше для синтеза АТФ. Когда в клетках возникает дефицит и глюкозы, и жирных кислот, окислению подвергается ряд аминокислот. При этом также образуется ацетил-КоА, или органические кислоты, которые окисляются далее в цикле трикарбоновых кислот. Аминокислоты - это последний энергетический резерв, который поступает в «топку» биологического окисления, когда исчерпаны другие резервы. Аминокислоты – дорогой «строительный материал», и они в основном служат для синтеза белков. Существенно, что при окислении глюкозы, жирных кислот и некоторых аминокислот образуется одинаковый конечный продукт – ацетил-КоА. При этом происходит «обезличивание» первичного источника энергии, поскольку ацетил-КоА не имеет никаких следов своего происхождения. Лекция 9. Фотосинтез как основной источник органических веществ на Земле Фотосинтез – единственный процесс на Земле, в результате которого энергия солнечного излучения превращается в энергию химических связей органических соединений. Органическое вещество, синтезируемое автотрофными организмами, передается по цепям питания; таким образом, за его счет существуют все организмы на Земле. 1. Значение фотосинтеза Значение фотосинтеза состоит в: 1) пополнении убыли органических соединений, непрерывно происходящей на нашей планете вследствие жизнедеятельности гетеротрофных организмов, а также производственной деятельности человека; 2) накоплении в продуктах фотосинтеза соответствующих количеств химической энергии; 3) поддержании уровня содержания в атмосфере кислорода, необходимого для существования подавляющего количества организмов, населяющих нашу планету; 4) устранении возможности накопления в атмосфере избытка углекислого газа. 2. Общие представления о химизме фотосинтеза Фотосинтез, протекающий в клетках зеленых растений и некоторых бактерий, является чрезвычайно сложным процессом. Упрощенно его химизм можно выразить в виде следующего уравнения суммарной реакции: квант света ↓ nH2O + nCO2 → → (CH2O)n + nO2↑ Фотосинтез осуществляется в 2 стадии (фазы): световую и темновую: квант света ↓ АТФ, НАДФН CO2 ↓ ↑ ↓ ↓ световые реакции → → → темновые реакции (цикл Кальвина) ↑ ↓ ↓ Н2О О2 (CH2O)n 3. Характеристика фотосинтетического аппарата Растения в своих зеленых частях имеют сложно устроенный фотосинтетический аппарат – фотосинтез происходит в зеленых пластидах - хлоропластах. Содержимое хлоропластов состоит из стромы (матрикса), имеющей белковую природу, и гран – зернистых образований, имеющих форму цилиндриков размером около 2 мкм. Граны состоят из плоских мешочкообразных пузырьков, имеющих двойную мембрану и называемых тилакоидами. В гранах они расположены стопками. В 1-й клетке зеленого листа, как правило, содержится 20-100 хлоропластов, а в каждом гране – 10-100 тилакоидов. Граны связаны между собой мембранными перемычками, которые называются ламеллами. В расчете на сухие вещества в хлоропласте содержится 50% белка, 35% липидов, 7% пигментов. Реакции световой стадии фотосинтеза происходят в тилакоидах, а темновой – в строме. Хлоропласты окрашены в зеленый цвет за счет зеленых пигментов (хлорофилла), которые расположены на внутренней мембране. Молекула хлорофилла состоит из тетрапиррольного кольца; в нем имеется 9 сопряженных двойных связей. Обязательным компонентом хлорофилла является атом магния, соединенных с 4-мя атомами азота пиррольных колец с помощью металлоорганической связи. 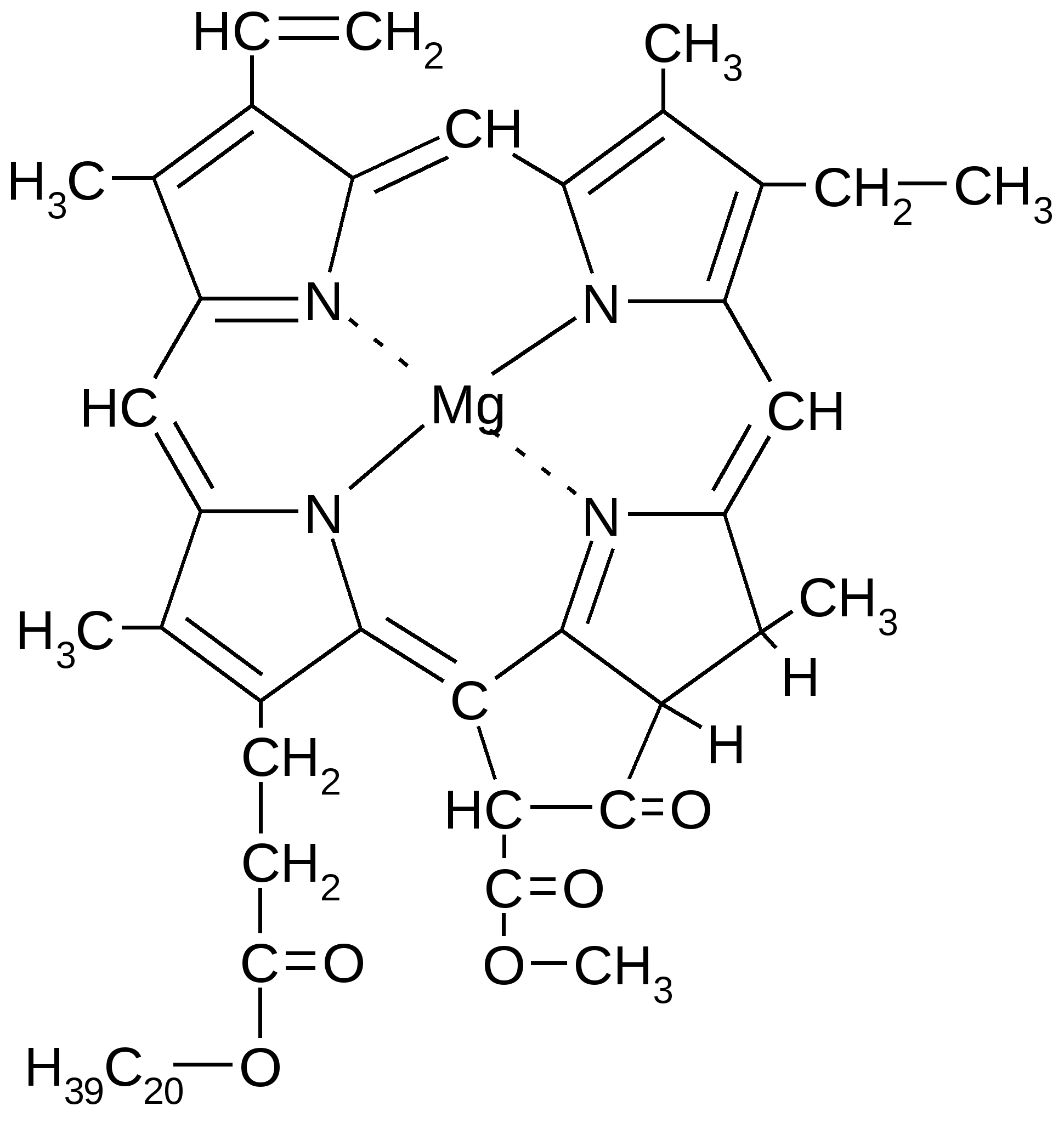 Функции хлорофилла при фотосинтезе:
Помимо хлорофилла к фотосинтетическим пигментам относятся каротиноиды – желтые, оранжевые, красные. Они всегда есть в фотосинтезирующих органах растений. Относятся к терпенам, которые построены на основе изопреновых блоков: 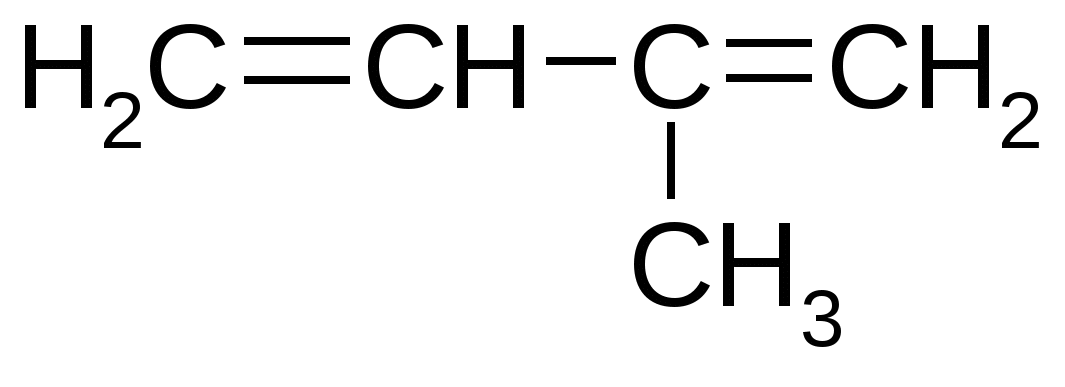 Функции каротиноидов: 1. Участие в поглощении света в качестве дополнительных пигментов. 2. Защита молекул хлорофилла от необратимого фотоокисления, которое может происходить под действием яркого света. Структура фотосинтетического аппарата Пигменты растений организованы в светособирающие комплексы, которые локализованы на внутренних мембранах хлоропластов. Функции светособирающих комплексов: поглощение света и передача энергии возбужденного состояния на реакционный центр. Реакционный центр – белок, содержащий молекулу хлорофилла, способную из возбужденного состояния отдавать электрон. Хлорофилл существует в возбужденном состоянии в течение 10 –12 - 10 – 9 сек. На ярком солнечном свету 1 квант света поглощается не чаще 1 раза за 0,1 сек. В течение остального времени молекула хлорофилла простаивает. Пигменты светособирающего комплекса поглощают энергию света и передают на реакционный центр для ее более производительного функционирования. Передача энергии осуществляется по принципу индуктивного резонанса. Необходимыми условиями для фотосинтеза являются:
Фотосистемы – белковые комплексы, с помощью которых энергия света превращается в энергию химических связей через энергию возбужденного состояния хлорофиллов.
4. Световая фаза фотосинтеза Во время световой фазы фотосинтеза происходят стабилизация и запасание световой энергии и ее трансформация в химическую энергию. Обязательным условием для осуществления световой стадии фотосинтеза является наличие света. Световая фаза фотосинтеза включает 4 основных процесса: 1) фотохимическое возбуждение хлорофилла; 2) фотоокисление (фотолиз) воды до кислорода, протонов и электронов; 3) фотовосстановление НАДФ окисленного до НАДФ восстановленного; 4) фотосинтетическое фосфорилирование (образование АТФ из АДФ и фосфорной кислоты при участии энергии света). Внутренняя мембрана хлоропластов непроницаема для протонов. Реакционный центр ФС 1 под действием кванта света переходит в возбужденное состояние, он окисляется и отдает электроны на ферредоксин, который передает их на фермент редуктазу. Редуктаза окисляет НАДФ+ до НАДФ Н. Под действием кванта света реакционный центр ФС 1 переходит в возбужденное состояние и окисляется, отдавая 2 электрона на комплекс переносчика. Таким образом, в полости тилакоида накапливаются протоны, в результате возникает разность концентраций протонов между сторонами мембраны и разность зарядов. За счет энергии электрохимического потенциала конформация фермента АТФазы изменяется, канал открывается и по нему протоны возвращаются в строму. Одновременно в головке АТФ-азы из АДФ и неорганического фосфата синтезируется АТФ. 5. Темновая фаза фотосинтеза Темновая фаза фотосинтез – это путь восстановления двуокиси углерода до простых сахаров. Этот процесс впервые был изучен американским ученым Кальвином, поэтому в его честь был назван циклом Кальвина. Цикл Кальвина функционирует у высших растений, водорослей, у большинства фототрофных бактерий. Первичным акцептором углекислого газа является рибулозо-1,5-бисфосфат. Ключевым ферментом, обеспечивающим фиксацию СО2, является фермент рибулозобисфосфаткарбоксилазаоксигеназа (Рубиско). Его активность строго коррелирует с общей интенсивностью фотосинтеза. Концентрация его в строме очень высока. На долю этого фермента приходится почти 60% растворимого белка стромы. Цикл Кальвина, весьма напоминающий обращенный пентозофосфатный путь дыхания, состоит из 4-х этапов:
3) Стадия регенерации первичного акцептора СО2. Происходит с участием фосфатов трех-, четырех-, пяти-, шести- и семиуглеродных сахаров. В результате процессов взаимопревращений фосфорилированных сахаров из 5 молекул трехуглеродных сахаров образуются 3 молекулы пятиуглеродных сахаров, которые затем превращаются в рибулозобисфосфат. 4) Стадия синтеза углеводных продуктов фотосинтеза. На этом этапе из фосфоглицеринового альдегида и фосфодиоксиацетона под действием фермента альдолазы синтезируется фруктозо-1,6-бисфосфат, который затем может превращаться или в крахмал, или сахарозу. Для синтеза 1 молекулы глюкозы в цикле Кальвина необходимо 12 молекул НАДФ восстановленного и 18 молекул АТФ. Процесс фотосинтеза осуществляется только на свету. Однако у растений часто возникает потребность в глюкозе, когда фотосинтез невозможен (в темное время суток, при прорастании семян). В этих случаях потребности в глюкозе удовлетворяются либо за счет распада сложных углеводов, либо путем новообразования глюкозы, которое называется глюконеогенезом. Глюконеогенез – процесс образования глюкозы из неуглеводных предшественников: пирувата, аминокислот, глицерина, жирных кислот и др.). По своему химизму этот процесс напоминает обращенный гликолиз. Глюконеогенез происходит и в организмах человека и животных. Однако ферменты, которые катализируют реакции гликолиза, имеются только в клетках печени и надпочечников. Поэтому глюконеогенез может осуществляться только в этих органах, он идет в тех случаях, когда возникает потребность в глюкозе (например, у клеток головного мозга), а запасов гликогена недостаточно и другие возможности обеспечить эти потребности отсутствуют. Раздел 6. ЛИПИДЫ И ИХ ОБМЕН Лекция 10. Классификация липидов, их свойства и биологическая роль Липиды представляют собой обширную группу органических соединений, существенно различающихся по своей химической структуре и функциям. Они не растворяются в воде, а хорошо растворимы в неполярных растворителях (таких, как эфир, хлороформ или бензол). Большинство липидов являются сложными эфирами многоатомных или специфически построенных спиртов с высшими жирными кислотами. 1. Классификация липидов Существует несколько классификаций липидов. Наибольшее распространение получила классификация, основанная на структурных особенностях липидов. По этой классификации различают следующие группы липидов. 1) Простые липиды: сложные эфиры жирных кислот с различными спиртами (например, ацилглицерины, или ацилглицеролы и воски). 2) Сложные липиды: сложные эфиры жирных кислот со спиртами, дополнительно содержащие и другие группы (например, фосфолипиды, включающие глицерофосфолипиды и сфинголипиды; гликолипиды, сульфолипиды, липопротеины и др.). 3) Предшественники и производные липидов (жирные кислоты, глицерол, стеролы и прочие спирты, жирорастворимые витамины, стероидные гормоны и др.) Разделение липидов по физико-химическим свойствам учитывает степень их полярности. По этому признаку липиды делят на нейтральные, или неполярные, и полярные. К первому типу относятся липиды, не имеющие заряда. Ко второму – липиды, несущие заряд и обладающие отчетливыми полярными свойствами (например, фосфолипиды, жирные кислоты). 2. Характеристика основных групп липидов Жирные кислоты Жирные кислоты – алифатические карбоновые (как правило, монокарбоновые) кислоты с длинной неразветвленной углеводородной цепью, состоящей чаще всего из нечетного числа атомов углерода в количестве от 14 до 20. Жирные кислоты с более длинной или более короткой цепочкой, а также с нечетным числом атомов углерода встречаются реже. В свободном состоянии встречаются в следовом количестве, но в организме их много, т.к. они входят в состав липидов. В природе обнаружено свыше 200 жирных кислот, однако в тканях человека и животных в составе простых и сложных липидов найдено около 70 кислот, причем более половины из них – в следовых количествах. По химическому строению жирные кислоты подразделяются на: 1) насыщенные (не имеют двойных связей): общая формула: СnH2nO2 или СН3-(СН2)n -CООН примеры:
2) ненасыщенные: мононенасыщенные и полиненасыщенные (имеют одну или несколько двойных связей):
В ненасыщенных жирных кислотах всегда встречается «цис» – конфигурация. Двойные связи никогда не сопряжены. Температура плавления насыщенных кислот высокая, поэтому они, а также липиды, в которых преобладают насыщенные жирные кислоты, имеют твердую консистенцию. Температура плавления ненасыщенных кислот низкая, поэтому в обычных условиях они имеют жидкую консистенцию, а следовательно, те липиды, в которых они преобладают, также отличаются жидкой консистенцией. Насыщенные жирные кислоты используются в живых организмах в основном как энергетический материал. При их избыточном количестве в рационе человека нарушается липидный обмен и повышается уровень холестерина. Ненасыщенные жирные кислоты в меньшей степени используются как источники энергии, но они играют очень важную роль в процессах метаболизма. Линолевая, линоленовая и арахидоновая кислоты являются незаменимыми, т.е. не синтезируются и должны поступать с пищей. Эти кислоты объединены в витамин F. Применение полиненасыщенных жирных кислот весьма эффективно при лечении гипертонической болезни, сахарного диабета, бронхиальной астмы и других заболеваний. Нейтральные жиры Нейтральные жиры (ацилглицерины, или ацилглицеролы) – сложные эфиры трехатомного спирта (глицерина) и жирных кислот. 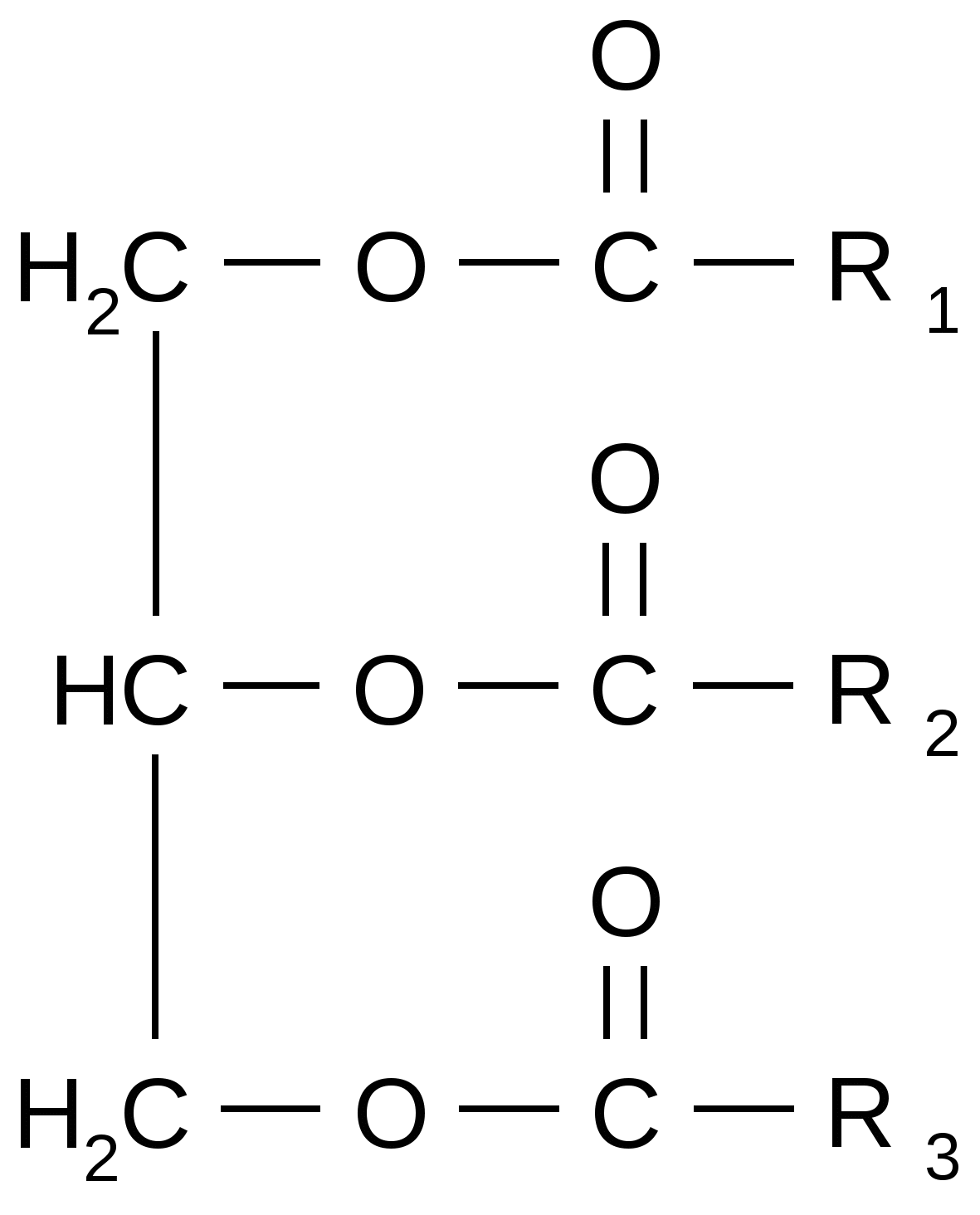 Если все радикалы одинаковые, то жиры называются простыми триацилглицеролами (тристеарин, тристеароглицерин). Если радикалы разные, они называются смешанными. Любой природный жир – это не индивидуальное химическое вещество, а смесь триацилглицеролов и свободных жирных кислот. Если нейтральный жир обогащен насыщенными жирными кислотами, то он имеет твердую консистенцию (как правило, жиры животного происхождения). Если много ненасыщенных кислот – то он жидкий (большинство жиров растительного происхождения). Гидролиз жиров, поступающих с пищей в организм человека, в основном происходит в 12-перстной кишке при участии фермента липазы. Липаза катализирует гидролиз только эмульгированных жиров, поэтому расщеплению жиров предшествует их эмульгирование под действием природных эмульгаторов, роль которых выполняют желчные кислоты, вырабатываемые печенью. Основные свойства жиров:
Фосфолипиды Фосфолипиды в зависимости от того, какой спирт входит в их состав, делятся на фосфоглицеролипиды (или фосфатиды) и фосфосфинголипиды. Фосфоглицеролипиды (фосфатиды) представляют собой сложные эфиры, состоящие из глицерина, жирных кислот, фосфорной кислоты и азотсодержащего спирта. Одним из наиболее распространенных фосфолипидов является лецитин. Главная функция фосфолипидов – они являются структурными компонентами клеточных мембран. Особенно богата фосфолипидами нервная ткань (мозг). Кроме того, они выполняют следующие функции:
Воски Воски – сложные эфиры высших жирных кислот и одноатомных высших спиртов (спиртов с длинной цепью). Воска образуют защитную смазку на коже, шерсти, перьях, покрывают листья и плоды растений, покрывают наружный скелет насекомых. Основная функция – предотвращают испарение избыточного количества влаги. Стероиды Стероиды – вещества, производные от спирта стерола. 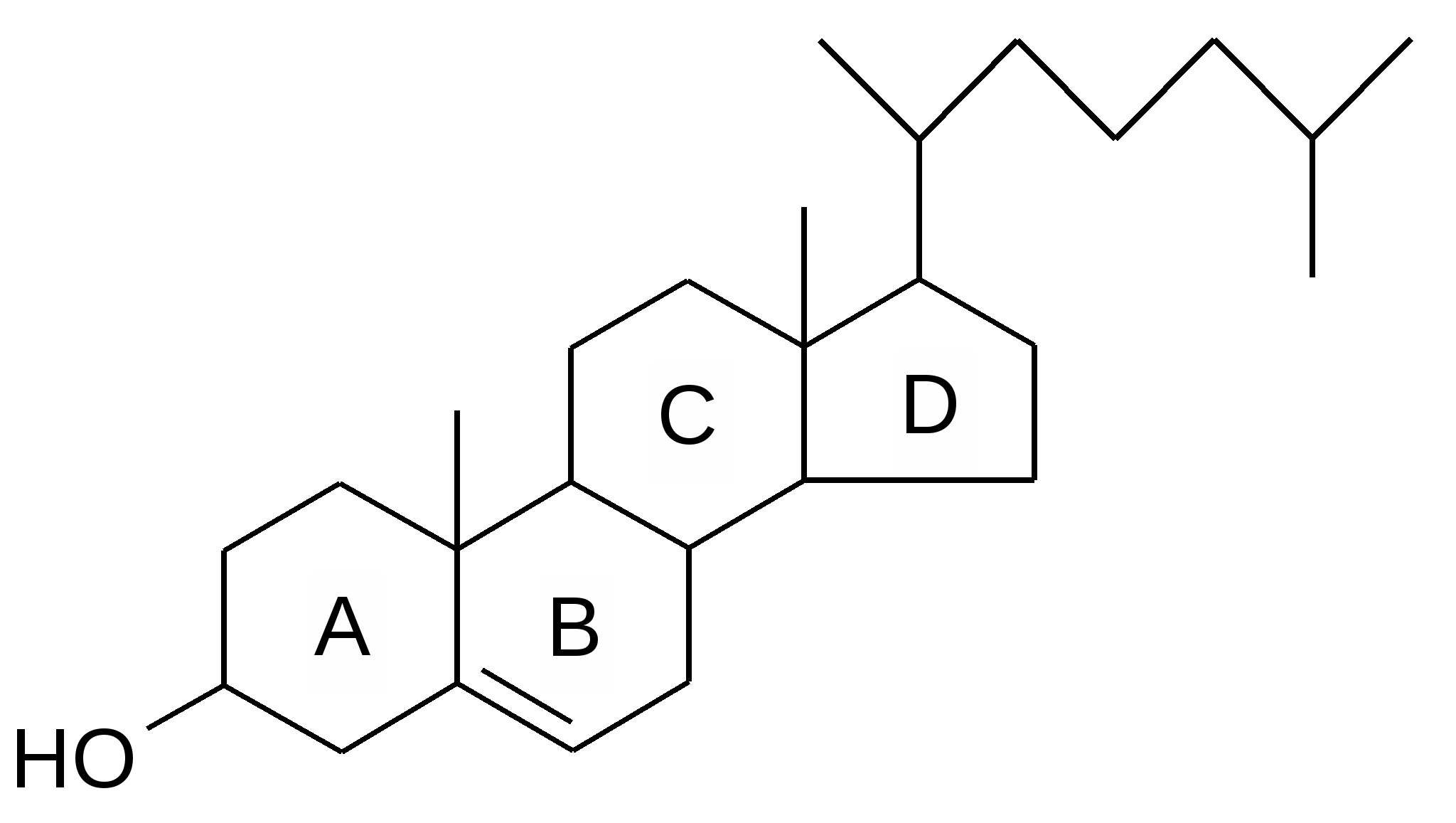 Холестерин, некоторые мужские и женские половые гормоны, витамин Д, желчные кислоты – относятся к стероидам. Холестерин – стероид животного происхождения; в растениях встречается в небольших количествах (семенах и пыльце). Организм нуждается в холестерине; 80% необходимого холестерина синтезируется самим организмом; 20% должно поступать с пищей. Холестерин является предшественником таких физиологически важных веществ, как желчные кислоты, стероидные гормоны, витамин D. Эфиры холестерина и жирных кислот являются транспортной формой жирных кислот. Холестерин накапливается в крови в случае, если в ней находится много насыщенных жирных кислот, что влечет за собой возникновение атеросклероза. Терпены Терпены – это ненасыщенные углеводороды с числом углеродных атомов, кратным 5. Их можно рассматривать как производные основной разветвленной единицы – изопрена. По числу изопреновых единиц в молекуле они делятся на:
К терпенам относится каучук (политерпен), витамин А (дитерпен), каротиноиды (тетратерпены). Эфирные масла (ментол, камфара) – тоже терпены. 3. Основные функции липидов По физиологическому значению липиды делят на резервные и структурные. Резервные липиды депонируются в больших количествах и затем расходуются для энергетических и пластических нужд организма. К резервным липидам относятся ацилглицерины. Все остальные липиды можно отнести к структурным липидам. Структурные липиды не имеют такой энергетической значимости, как резервные липиды. Они участвуют в построении биологических мембран, защитных покровов растений, насекомых и кожи позвоночных. В целом различные группы липидов выполняют следующие функции:
4. Липиды зерна и продуктов его переработки Когда производят определение жира в зерне или муке, его экстрагируют диэтиловым эфиром. При этом в эфирный экстракт переходят все вещества, относящиеся к группе липидов, и в экстракте находится так называемый сырой жир, т.е. смесь жира с жироподобными веществами: фосфатидами, стеролами, восками и каротиноидами. |