Майкл Дж[1]. Наука о гибкости. Литература ббк 75. 0 А52
 Скачать 10.31 Mb. Скачать 10.31 Mb.
|

|
Стимул Локальное Генератор Локальная деполяри- Проведенный (растягивание) изменение тока (передача зация (генератор потенциал проницаемости заряда) потенциала) действия Нервно-сухожильные веретена. Нервно-сухожильные веретена — это чувствительные к сокращению механорецепторы скелетных мышц 112 Г л а в а 6. Нейрофизиология гибкости: невралъная анатомия и физиология    млекопитающих, иннервируемые быстропроводящими афферентными нервными волокнами группы 16, имеющими большой диаметр (Jami, 1992). Впервые нервно-сухожильные веретена были обнаружены и описаны Гольджи в 1903 году. Ввиду технических трудностей нервно-сухожильные веретена не так хорошо изучены, как нервно-мышечные. Тем не менее их важность с точки зрения развития гибкости нельзя недооценивать. млекопитающих, иннервируемые быстропроводящими афферентными нервными волокнами группы 16, имеющими большой диаметр (Jami, 1992). Впервые нервно-сухожильные веретена были обнаружены и описаны Гольджи в 1903 году. Ввиду технических трудностей нервно-сухожильные веретена не так хорошо изучены, как нервно-мышечные. Тем не менее их важность с точки зрения развития гибкости нельзя недооценивать.Местонахождение и структура нервно-сухожильных веретен Нервно-сухожильные веретена находятся почти исключительно у апоневрозов или мышечно-сухожильных соединений, но не в сухожилиях (рис. 6.6). Так, Пенг, исследовав (цитируется по Bavker, 1974) 1337 рецепторов из различных участков мышц кошек, установил, что 92,4 % их располагались у мышечно-сухожильных соединений и только 7,6 % •— в самом сухожилии. 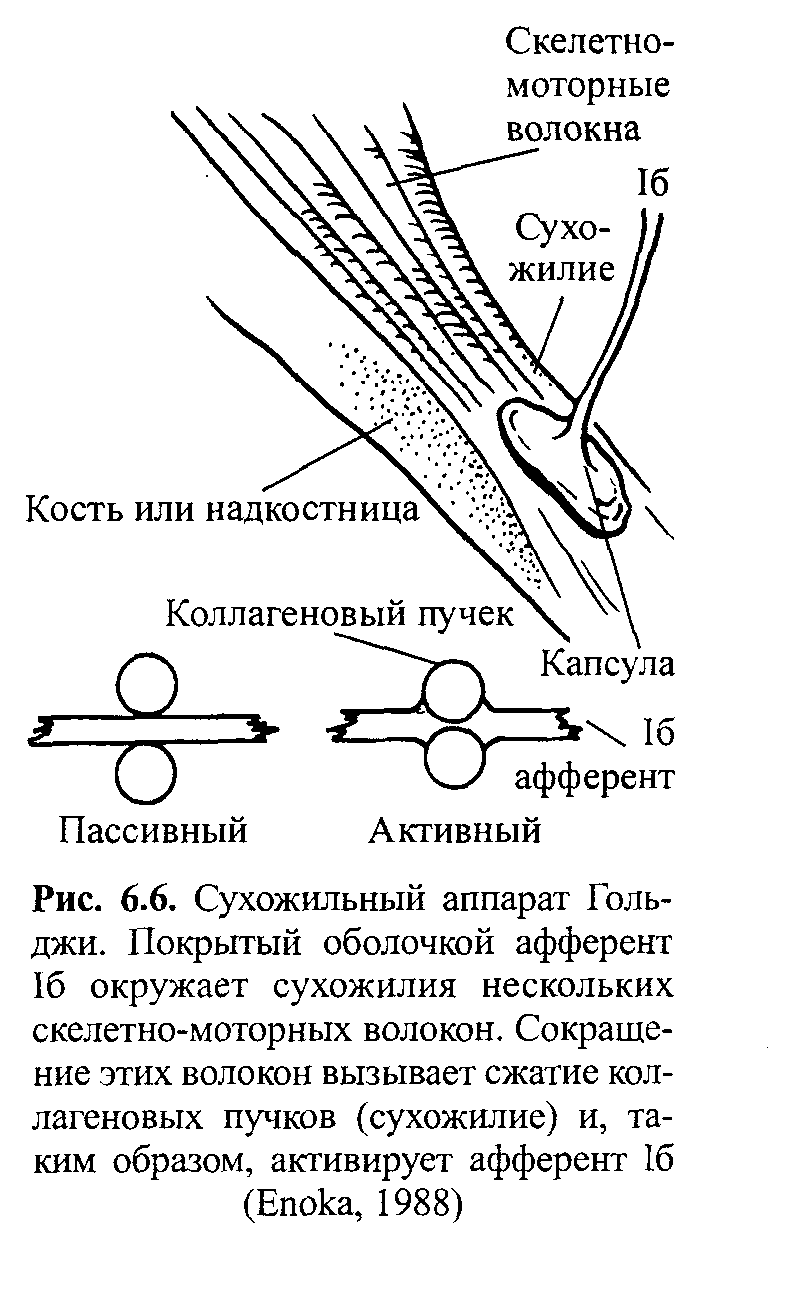 У немлекопитающих нервно-сухожильные веретена представляют собой расположенные вдоль сухожильных пучков не покрытые оболочкой рецепторы (пучки коллагеновых волокон). У млекопитающих же, напротив, они покрыты оболочкой и, как уже упоминалось, располагаются у мышечно-сухожильных или мышечно-апоневротических соединений. Значение этого различия, по мнению исследователей, состоит в следующем. Во-первых, окруженные оболочкой веретена являются более чувствительными к данной величине стимула и более точными в определении и передаче информации в ЦНС. Во-вторых, местонахождение нервно-сухожильных веретен обеспечивает их экстремальную чувствительность к любому изменению напряжения отдельных мышечных волокон, к которым они прикреплены (J.C.Moore, 1984). К,и Нервно-сухожильные веретена располагаются последовательно, в отличие от нервно-мышечных веретен, расположенных параллельно мышечным волокнам. В исследовании Пенга количество мышечных волокон, соединенных последовательно с нервно-сухожильными веретенами, колеблется от 3 до 50. Каждое нервно-сухожильное веретено, как правило, иннервируется отдельным быстропроводящим афферентным нервным волокном группы 16 (Jami, 1992). ИЗ Наука о гибкости Функция нервно-сухожильных веретен На данном этапе мы лишь частично знаем многочисленные функции нервно-сухожильных веретен. Последние исследования показывают, что эти рецепторы являются более сложными, чем первоначально считалось (J.C.Moore, 1984). Сейчас мы знаем, что нервно-сухожильные веретена контролируют все степени мышечного напряжения. В то же время они более всего чувствительны к напряжению, производимому в результате мышечных сокращений (см. рис. 6.6). Предполагают также, что нервно-сухожильные веретена вносят свой вклад в сознательные восприятия. Это предположение основано на том факте, что импульс из нервно-сухожильных веретен достигает коры головного мозга, той его части, которая осуществляет интерпретацию сенсорной активности тела (Roland и Ladegaard-Pedersen, 1977). Кроме того, высказывается мнение, что нервно-сухожильные веретена способствуют снижению мышечного утомления (Вагг, 1979; Kandel, Swartz, 1981; Lundberg, 1975). По мере увеличения напряжения в группе мышечных волокон нервно-сухожильные веретена и их афференты группы 16 посылают увеличивающееся число сигналов в ЦНС. Эти чувствительные нервы заканчиваются в спинном мозгу на небольших интернейронах, которые затем ингибируют клеточные тела мотонейронов, активирующих сокращающуюся мышцу. Данный процесс называется автогенным ингибированием, так как сокращения мышцы ингибируются ее собственными рецепторами. Результирующее снижение мышечной силы уменьшает активацию нервно-сухожильных веретен и количество инги-биторной обратной связи, поступющей в ЦНС из этих мышц. Такое мгновенное ингибирование обратной связи способствует увеличению напряжения в мышцах. Неправильные представления о нервно-сухожильных веретенах За прошедшее время появилось немало ошибочных представлений о нервно-сухожильных веретенах. Одно из них касается взаимосвязи между нервно-сухожильными веретенами и измерением растягивающих усилий. Нервно-сухожильные веретена нередко называют «рецепторами растяжения». Этот термин подразумевает, что пассивное напряжение, сопровождающее растягивание мышцы, может также представлять собой адекватный стимул для нервно-сухожильных веретен. Нервно-сухожильные веретена, действительно, могут активироваться в результате пассивного напряжения, однако их порог для данного вида стимулов является очень высоким. Поэтому для активизации нервно-сухожильных веретен необходимо очень интенсивное растягивание (Houk, Singer, Goldman, 1971). Кроме того, разрядка нервно-сухожильных веретен очень редко продолжается во время поддерживаемого растяжения мышцы. Другое неверное представление относительно нервно-сухожильных веретен заключается в том, что их чувствительность является недостаточ- 114    Г л а в а 6. Нейрофизиология гибкости: невралъная анатомия и физиология Г л а в а 6. Нейрофизиология гибкости: невралъная анатомия и физиологияной для активизации сократительного напряжения. Исследования показали, что нервно-сухожильные веретена проявляют очень низкий порог и весьма ощутимую динамическую чувствительность при тестировании с применением адекватного стимула. Таким образом, нервно-сухожильные веретена способны сообщать об очень незначительных и быстрых изменениях в силах сокращения (Houk и Henneman, 1967; Houk, Singer, Goldman, 1971; Jami, 1992). Некоторое время существовало мнение, что функция нервно-сухожильных веретен заключается в автогенном ингибировании, т. е. ингиби-ровании агонистов и синергистов и содействии антагонистам. В настоящее время доказано, что это лишь одна из многих функций нервно-сухожильных веретен, которым помогают в этом, по меньшей мере, еще два рецептора: низкопороговые рецепторы суставной капсулы и низкопороговые кожные рецепторы (J.C.Moore, 1984). Цель такого аутогенного ингибиро-вания связывают с защитными функциями. 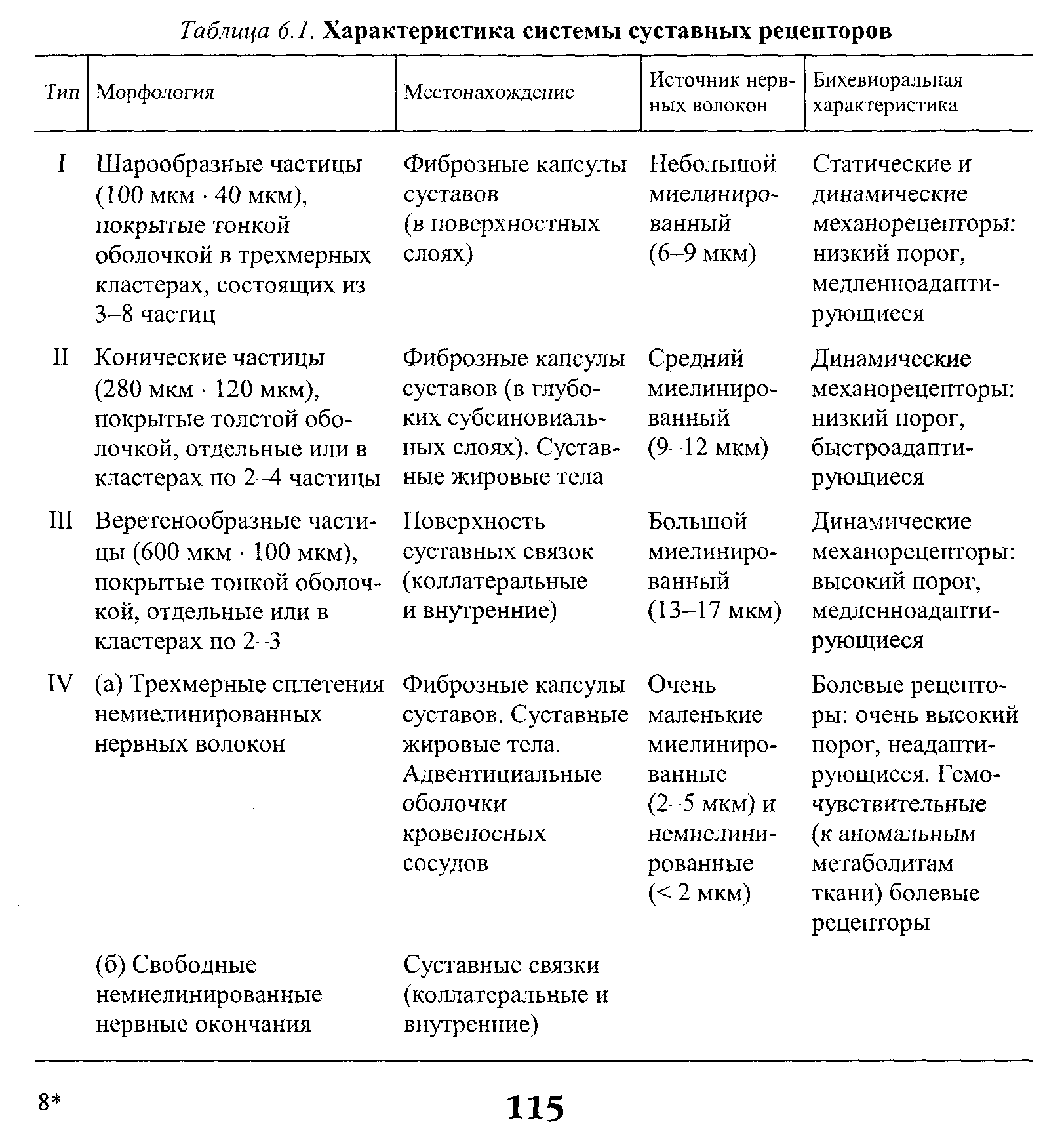 Наука о гибкости Также известно, что влиянию нервно-сухожильных веретен могут противостоять дополнительные сигналы, поступающие из высших центров. Процесс снижения к минимуму влияния нервно-сухожильных веретен называется растормаживанием мотонейронов-агонистов. О важности этого процесса для спорта говорили Брукс и Фехи (1987): «Использование растормаживания применяется в спортивной тренировке с целью довести физическую работоспособность до пределов возможности тканей. В таком виде спорта, как армрестлинг, иногда встречаются разрывы мышц и сухожилий и переломы костей. У высокомотивированных спортсменов и у расторможенных индивидуумов сочетание активного мышечного сокращения и напряжения, производимого соперником, может превысить силу тканей». Суставные механорецепторы (рецепторы суставов). Все синовиальные суставы тела имеют четыре различных нервных окончания. Эти суставные рецепторы ощущают механические силы, действующие на суставы, такие, как давление растяжения, и поэтому их называют суставными механо-рецепторами. Выделяют четыре типа суставных механорецепторов (табл. 6.1). Такая классификация основана на соответствующих морфологических и бихевиоральных характеристиках нервных окончаний. В следующих разделах мы рассмотрим эти четыре вида суставных механорецепторов. Суставные рецепторы типа I Механорецепторы типа I состоят из кластеров тонких, покрытых оболочкой шарообразных частиц. Они находятся главным образом во внешних (поверхностных) слоях фиброзной суставной капсулы. Каждый кластер состоит приблизительно из 8 частиц. Каждая частица иннервируется отдельным миелинированным волокном группы II (диаметром 6-9 мкм). Плотность этих механорецепторов более высокая в проксимальных (например, тазобедренный), чем в дистальных (например, голеностопный) суставах. С физиологической точки зрения частицы типа I ведут себя как низкопороговые медленноадаптирующиеся рецепторы. Следовательно, они реагируют на очень незначительную механическую нагрузку и продолжают выпускать нервные импульсы на протяжении действия механического стимула. Чтобы стимулировать их, достаточно усилие порядка 3 г. Более того, часть этих рецепторов низкого порога всегда активна при любом положении сустава и даже в том случае, когда он неподвижен. Их интенсивность разрядки в покое обычно имеет частоту примерно 10-20 Гц (импульсов в секунду). Механорецепторы типа I имеют целый ряд функций, включая сообщение направления, амплитуды и скорости движений суставов; регуляцию изменений давления в суставе; содействие постуральным и кинестетическим ощущениям; содействие ЦНС в регуляции постурального мышечного тонуса, а также тонуса мышц при движениях сустава; осуществление ин-гибиторного влияния на поток болевой афферентной активности из суставной рецепторной системы типа IV. Рецепторы типа I можно разделить на статические и динамические механорецепторы. 116  Г л а в а 6. Нейрофизиология гибкости: невралъная анатомия и физиология Г л а в а 6. Нейрофизиология гибкости: невралъная анатомия и физиологияСуставные рецепторы типа II Рецептор типа II представлен крупными коническими частицами, имеющими плотную оболочку. Они располагаются в фиброзной суставной капсуле, однако в более глубоких ее слоях. Каждый кластер обычно состоит из 2-А частиц. Кроме того, каждый компонент кластера иннервируется ответвлением миелинированных суставных нервных волокон группы II (диаметром 9-12 мкм). Наибольшая плотность механорецепторов типа II также наблюдается в более дистальных суставах (например, голеностопный), чем в более проксимальных (например, тазобедренный). С физиологической точки зрения рецепторы типа II, подобно рецепторам типа I, имеют низкий порог. Однако они ведут себя как быстроадапти-рующиеся механорецепторы и не разряжаются в покое. Следовательно, они полностью пассивны в неподвижных суставах. Рецепторы типа II статически не разряжаются, поскольку их разрядка зависит от скорости. Поэтому их называют динамическими механорецепторами, или механорецеп-торами ускорения. Во время стимулирования каждый кластер осуществляет кратковременный высокочастотный «всплеск» импульсов в течение менее чем 1 с, а нередко и менее 0,5 с. Главная функция этих рецепторов — определение быстрых изменений в движении, таких, как ускорение и замедление. Суставные рецепторы типа III Механорецепторы типа III представляют собой покрытые тонкой оболочкой частицы, заключенные во внутренние (в пределах суставной капсулы) и внешние (снаружи суставной капсулы) связки большинства суставов. Они отсутствуют в связках позвоночного столба. Это самые большие суставные частицы, которые, подобно нервно-сухожильным веретенам, ведут себя как высокопороговые медленноадаптирующиеся механорецепторы. Они иннервируются миелинированным афферентным аксоном группы I, диаметр которого может достигать 17 мкм. С физиологической точки зрения, механорецепторы суставных связок имеют высокий порог. Рецепторы типа III являются полностью пассивными в неподвижных суставах и реагируют только при генерировании высокого напряжения в суставных связках. При стимулировании рецепторы типа III проявляют частоту разрядки, являющуюся непрерывной функцией величины напряжения. Так как это медленноадаптирующиеся рецепторы, разрядка снижается очень медленно (в течение многих секунд), если поддерживается смещение удаленного сустава или тяговое усилие сустава. Механорецепторы типа III имеют две основные функции. Первая — контроль направления движения. Вторая — значительное рефлекторное ингибирование активности некоторых мышц сустава. Таким образом, они могут служить тормозящим механизмом, предупреждающим чрезмерную нагрузку на сустав. 117 Наука о гибкости Суставные рецепторы типа N В отличие от механорецепторов, нервные окончания типа IV не имеют оболочки. Они делятся на два вида. Окончания типа IVa представлены решетчатыми сплетениями, которые можно обнаружить в жировых прослойках суставов, а также в суставной капсуле. В то же время они полностью отсутствуют в синовиальной ткани, внутрисуставных менисках и суставных хрящах. Рецепторы типа IV6 — это свободные нервные окончания, не связанные со специализированными структурами. Они содержатся во внутренних и внешних связках. Рецепторы типа IVa и IV6 представляют собой систему болевых рецепторов суставных тканей. В нормальных условиях эти рецепторы являются полностью пассивными. Они проявляют активность, когда суставные ткани, в которых они содержатся, подвергаются существенной механической деформации или химическому раздражению. Примерами химических раздражителей являются такие агенты, как брадикинин, простангландин-Е, молочная кислота, полипептиды, гистамины. Эти вещества появляются в условиях ишемии (нехватка крови) и гипоксии (нехватка кислорода). Как утверждает Уайк (1972): «Рецепторы категории IV полностью отсутствуют в синовиальной выстилке каждого изученного сустава, а также в менисках коленного и ви-сочно-нижнечелюстного суставов и межпозвонковых дисках. Таким образом, не существует механизма, посредством которого могла бы возникать боль непосредственно в синовиальной ткани или мениске любого сустава, следовательно, хирургическое удаление синовиальной ткани или мениска сустава не означает, таким образом, удаление чувствительной к боли суставной ткани». Следовательно, подобное хирургическое вмешательство не приведет к снятию боли. РЕФЛЕКСЫ И ДРУГИЕ СПИННОМОЗГОВЫЕ НЕВРАЛЬНЫЕЦЕПИ Рефлекс представляет собой нейронную цепь, состоящую из чувст- вительного мотонейрона, ассоциативного или промежуточного нейрона и мотонейрона со своим эффектом.При действии на оконча- ние рецептора соответствующего стимула инициирует импульс, который проходит вдоль афферентного отростка к спинному мозгу, где он синапсирует с соединяющим нейроном. Мотонейрон возбуж- дается, и нервный импульс проводится по эфферентому волокну в клетку мышцы или железы. Иным словами, рефлекс - это реакции на стимул. Рефлекс растяжение мыцщы. Согласно классическому опредению рефлекса растягивания мышцы, всякий раз, когда мыщца растягива- ется, в действие вступает механизм рефлекса растяжения. Растягива- ние мыщцы приводит к удлинению как экстрафузальных мышечных  волокон, так и 118       Г л а в а 6. Нейрофизиология гибкости: невралъная анатомия и физиология Г л а в а 6. Нейрофизиология гибкости: невралъная анатомия и физиологиянервно-мышечных веретен (т. е. внутрифузальных волокон). Происходящая в нервно-мышечных веретенах деформация активирует первичные и вторичные окончания, в результате чего возникают потенциалы действия в чувствительных нейронах групп 1а и II. Эти нейроны достигают спинного мозга, где заканчиваются на клеточных телах а- (больших) мотонейронов. Если чувствительные афференты обеспечивают достаточную деполяризацию мотонейрона, он генерирует потенциалы действия. Его аксон, идущий к скелетной мышце, передает импульс, вызывающий рефлекторное сокращение. Схематически это показано на рис. 6.7. Аксон 1а передает афферентную информацию из нервно-мышечного веретена в центральную нервную систему. Аксон 16 обеспечивает такую же передачу, но с нервно-сухожильного веретена; а — гомонимная взаимосвязь — нервно-мышечные и нервно-сухожильные веретена, расположенные в мышце, соединяются с а-мотонейронами, которые активируют эту же мышцу. Афферентные и эфферентные аксоны, иннервирующие мышцы, расположенные в правой части тела, входят и выходят из спинного мозга справа, и наоборот; б — такие же соединения группы мышц агонистов-антагонистов (например, подколенные сухожилия и четырехглавые мышцы правой ноги), однако с выделением сложности межнейронных соединений, о — возбудительные соединения, • — ингибиторные воздействия, а — а-мотонейрон, у — у-мотонейрон, i — интернейрон, 1-1 a — ингибиторный интернейрон, la — афферент нервно-мышечного веретена, 16 — афферент нервно-сухожильного веретена, R — клетка Реншоу. 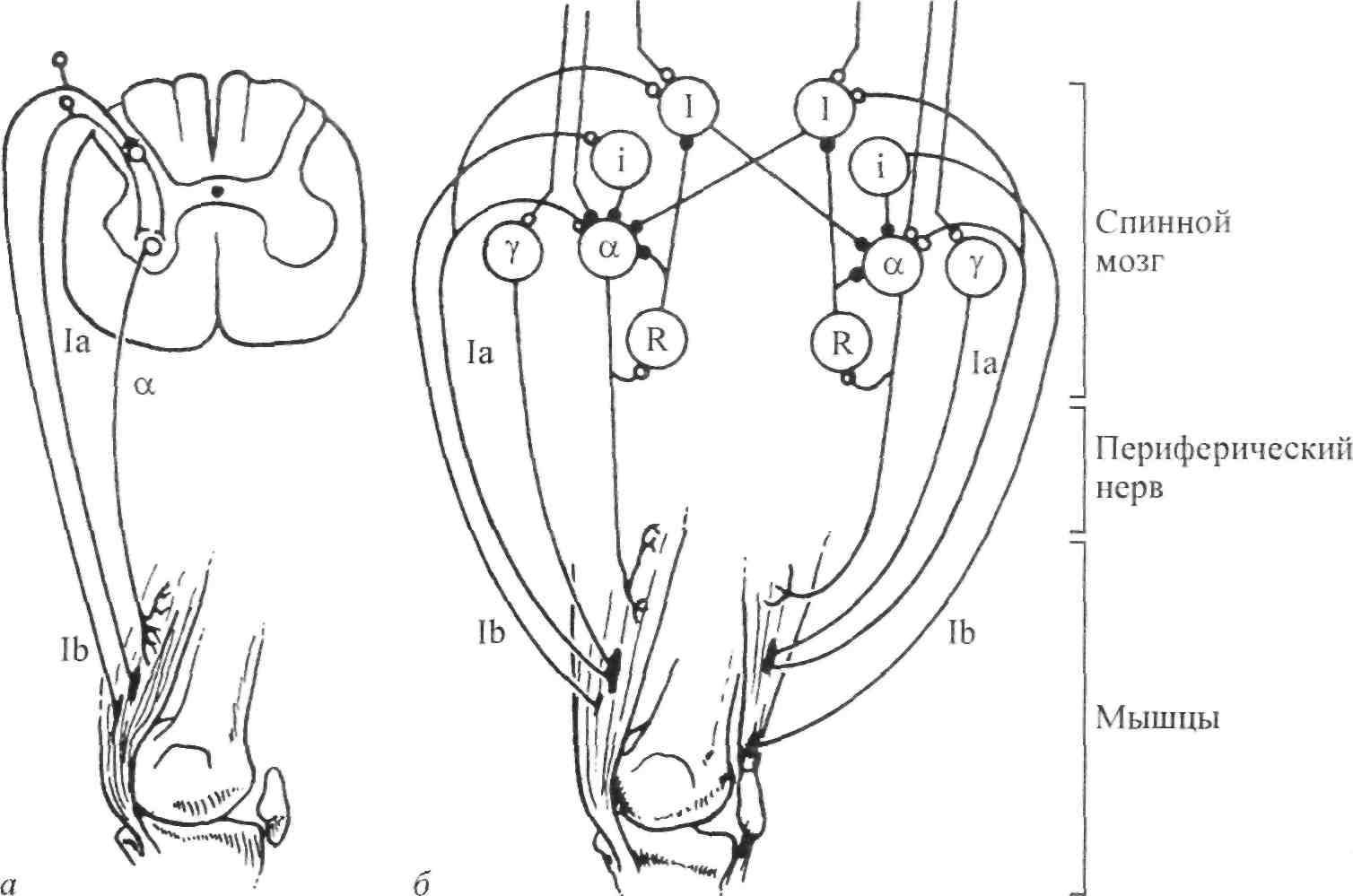 Рис. 6.7. Спинномозговые соединения между чувствительными рецепторами, находящимися в мышце, и а-мотонейронами (Enoka, 1988) 119 Супраспинальный импульс Наука о гибкости Реакция рефлекса растяжения имеет две составляющие: фазную и тоническую. Фазная реакция представляет собой начальный всплеск потенциалов действия, ведущий к быстрому повышению мышечного напряжения, пропорциональному скорости растяжения. Тоническая реакция является заключительной фазой медленного (низкочастотного) генерирования потенциалов, которое продолжается в течение всего растягивания и которое пропорционально количеству растяжений. Классическим примером фазного типа рефлекса растяжения является коленный, или пателлярный, рефлекс. Так, если легко ударить по пателляр-ному сухожилию (находящемуся под коленом), нервно-мышечные веретена, расположенные параллельно мышечным волокнам четырехглавой мышцы, растягиваются, вызывая деформацию нервно-мышечных веретен. Вследствие этого увеличивается интенсивность разрядки афферентов нервно-мышечных веретен группы 1а (первичное окончание возбуждается сильнее, чем вторичное, поскольку первое находится в более растяжимом центральном участке веретена интрафузального мышечного волокна). Сообщение затем направляется в спинной и головной мозг. Завершая рефлекторный акт, спинной мозг направляет эфферентные нервные импульсы в четырехглавую мышцу и вынуждает ее кратковременно сокращаться, что приводит к укорачиванию мышцы и снятию напряжения с нервно-мышечных веретен (рис. 6.8, где: а — влияние гомонимных а-мотонейронов; б — передача реципрокно-ингибиторного рефлекса через Ia-ингибиторный интернейрон (I); в — супраспинальный импульс, поступающий в четырехглавую мышцу, активирует реципрокно-ингибиторную цепочку в подколенных сухожилиях; г — возвратное торможение а-мотонейрона и 1а-ингиби-торного интернейрона через клетку Реншоу (R). I — 1а ингибиторный интернейрон, i — интернейрон, R — клетка Реншоу, а — а-мотонейрон, у — у-мотонейрон). Другим видом рефлекса растяжения является рефлекс статического, или тонического, растяжения. Стимулом при этом служит поддерживаемое растяжение, а реакцией — соответствующее поддерживаемое сокращение. Реакция на поддерживаемое растяжение частично обусловлена действием афферентов группы П. Примером тонической реакции может служить пос-туральная реакция на растяжение, которая проявляется в сокращении икроножной мышцы с целью коррекции чрезмерного смещения центра тяжести вперед в положении стоя. Реципрокная иннервация. Обычно мышцы «работают» в паре: когда одна группа мышц — агонисты — сокращается, противоположная группа мышц — антагонисты — расслабляется. Такая организация называется ре-ципрокной иннервацией. Например, при сгибании руки в локтевом суставе благодаря сокращению двуглавой мышцы трехглавая мышца, выпрямляющая руку в локтевом суставе, должна быть расслабленной. В противном случае обе мышцы противодействовали бы друг другу и движение выполнить невозможно. Если мотонейроны, идущие к одной мышце, получают возбудительные импульсы, ведущие к мышечному сокращению, мотонейроны, идущие к противоположной мышце, должны получить невральные сигналы, сни- 120 Г л а в а 6. Нейрофизиология гибкости: невралъная анатомия и физиология 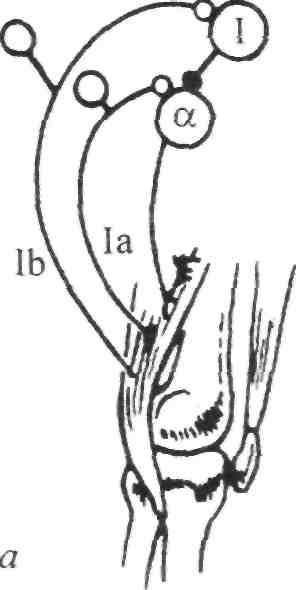 Четырехлавая мышца Подколенные сухожилия 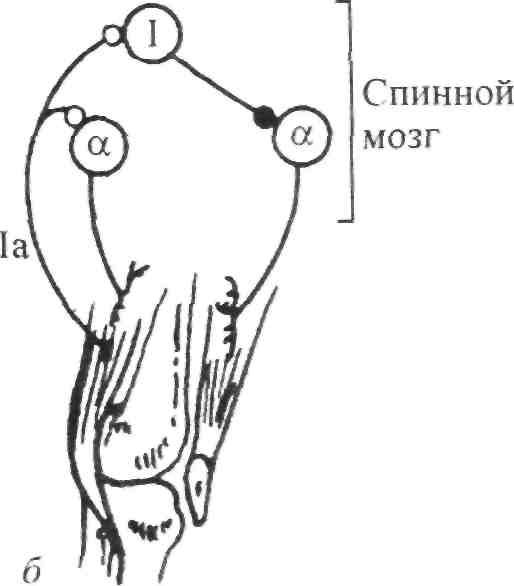 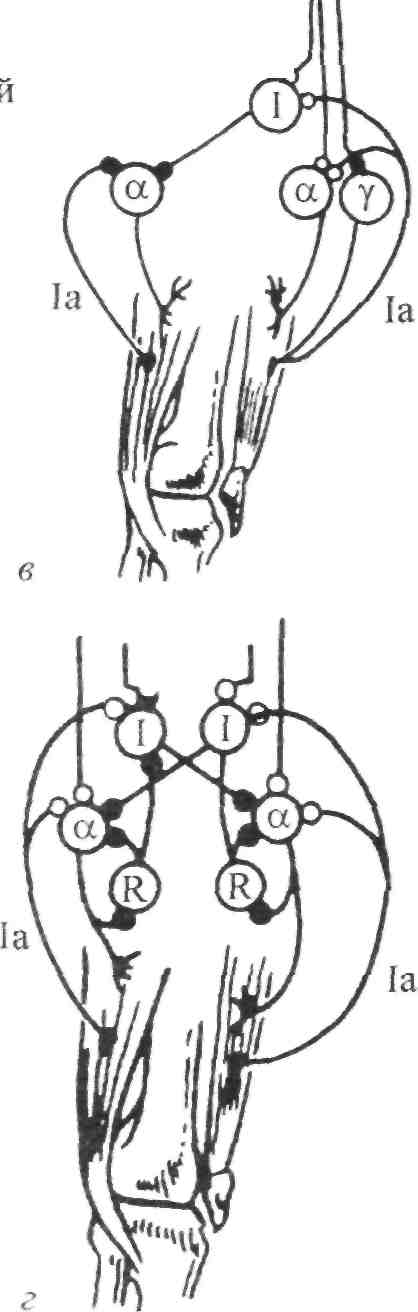 Рис. 6.8. Избранные центральные соединения (• — ингибиторные, о — возбудительные) эффе-рентов нервно-мышечного (1а) и нервно-сухожильного (16) веретена с группой четырехглавой мышцы и мышц подколенных сухожилий (Enoka, 1988) Рис. 6.8. Избранные центральные соединения (• — ингибиторные, о — возбудительные) эффе-рентов нервно-мышечного (1а) и нервно-сухожильного (16) веретена с группой четырехглавой мышцы и мышц подколенных сухожилий (Enoka, 1988)Супраспинальный импульс жающие вероятность их разрядки и производства мышечных сокращении (ингибирование). Антагонисты, таким образом, ингибируются практически в тот же самый момент, когда сокращаются агонисты. Рефлекторное торможение контролируется небольшим ингибиторным нейроном (который находится в спинном мозгу), идущим к мотонейронам, иннервирующим антагонистическую мышцу реципрокной пары. С другой стороны, при растяжении антагонистической мышцы мышца-агонист должна подвергнуться реципрок-ному торможению. Без этой реципрокной иннервации координированная мышечная активность была бы невозможной. Коактивация (сосокращение). Коактивацию можно определить как сокращение двух противоположных мышц с «высоким уровнем активности мышц-агонистов и одновременно с низким уровнем активности мышцы-антагониста в одном и том же суставе» (Solomonow, D'Ambrosia, 1991). Любопытно, что еще в 1909 г. Шеррингтон указывал на то, что мышцы-антагонисты могут сокращаться одновременно. Он объяснил это двойной реципрокной иннервацией. Спустя несколько лет Тилни и Пайк (1925) пришли к выводу, что «мышечная координация зависит главным образом от синхронной со сократительной связи в группе мышц-антагонистов». Левин и Кабат (1952) в результате своих наблюдений отметили, что нормальное 121   Наука о гибкости Наука о гибкостипроизвольное движение человека не дает основания для предположения о том, что реципрокная иннервация играет доминирующую роль в координировании сокращений мышц-антагонистов. Более того, «сокращение скорее закономерность, нежели исключение». В исследованиях последних лет было установлено, что имеет место центральный и периферический контроль коактивации агонистов-антаго-нистов при различных видах движения суставов (DeLuca, 1985; Kudina, 1980; Rao, 1965). Выявив факт существования сосокращения, следует выяснить и его цель. Соломонов и Д'Амбросия (1991), проведя анализ литературы, определили две очевидные цели. Во-первых, сосокращение обеспечивает плавное, тщательно регулируемое и точное движение сустава. Без такого регулирования выполнение двигательных задач повседневной деятельности, а также занятия спортом были бы невозможными. Во-вторых, сосокращение обеспечивает стабильность сустава и делает его более жестким. Следовательно, выполнение движения затрудняется (Enoka, 1988). Таким образом, мышцы обеспечивают значительную и весьма важную основу для сохранения стабильности сустава посредством сосокращения. Аутогенное ингибирование (обратный рефлекс растяжения мышцы). Чем сильнее мышца растягивается, тем выше сопротивление движению. Увеличение сопротивления объясняется рефлексом растяжения мышцы. Однако по достижении определенного предела сопротивление внезапно снижается, подобно тому, как закрывается лезвие перочинного ножа. Это явление нередко называют «реакцией закрытого ножа»; физиологи именуют его удлиненной реакцией. Первоначально считали, что оно вызвано действием нервно-сухожильных веретен. В настоящее время полагают, что оно обусловлено афферентным импульсом нервных окончаний группы II, идущих из нервно-мышечных веретен и, возможно, из миелинированных волокон, передающих болевые ощущения из суставов (Moore, 1984). Для подтверждения этой гипотезы требуются дополнительные исследования. ПЛАСТИЧНОСТЬ НЕВРАЛЬНЫХ ЦЕПЕЙ СПИННОГО МОЗГА Механизмы памяти или длительного адаптационного изменения в ЦНС давно интересуют ученых. Согласно Волпау и Керп (1990): «Спинной мозг в целом и рефлексы спинного мозга в частности давно считают стабильными и стереотипно реагирующими на импульсы, поступающие из периферических или супраспиналь-ных участков. Это распространенное представление является ошибочным. Нейроны и синапсы спинного мозга, как и нейроны и синапсы коры головного мозга и других супраспинальных структур, изменяются в процессе развития и в ответ на повреждения». 122    Г л а в а 6. Нейрофизиология гибкости: невралъная анатомия и физиология Г л а в а 6. Нейрофизиология гибкости: невралъная анатомия и физиологияНе так давно было установлено, что активность нейронов может вызывать постоянные изменения в ЦНС. Эти пластические изменения, такие, как образование новых синаптических соединений или модификация ионной проводимости определенных мембран, по мнению ученых, обусловливают последующие изменения активности ЦНС, которые выражаются в измененном поведении (Wolpaw и Lee, 1989). Изучение этого феномена требует организации и проведения ряда исследований. Метод изучения пластичности и полученные результаты. С начала 1980 гг. ученые стали изучать способность приматов изменять степень рефлекса спинального растяжения, проводя системное исследование анатомических и физиологических субстратов, управляющих памятью. Исследуемый рефлекс, или Ml, представляет собой начальную реакцию на внезапное растяжение мышцы. Рефлекс спинального растяжения — это наиболее простое поведение ЦНС позвоночных (Matthews, 1972). Рефлекс Хоффмана (Н-рефлекс) сопоставим с рефлексом спинального растяжения, за исключением того, что он вызывается непосредственным электрическим стимулированием 1а афферентных волокон, а не механическим растяжением. В серии экспериментов двуглавые мышцы плеча и трехглавые икроножные мышцы обезьян подвергли выработке оперантного условного рефлекса, имплантировав приспособление с ЭМГ-электродной обратной связью и используя в качестве поощрения сок. Обезьяны должны были выполнять задание, предусматривающее продолжительное изменение нейронной активности, влияющее на проводящий путь рефлекса спинального растяжения, и таким образом оно могло произвести след памяти, находящийся в этом проводящем пути (Wolpaw, 1983). Исследования, длившиеся 250 дней, показали способность обезьян увеличивать или снижать степень рефлекса спинального растяжения и Н-рефлекса (рис. 6.9). Начало импульса помечено стрелками, средняя продолжительность обусловленного импульсом выпрямления показана нижним следом. Фоновая (основная) ЭМГ стабильна в течение 4 серий. Амплитуда рефлекса спинального растяжения увеличивается, превышая контрольную, при минимальном (вверх) вызывании рефлекса, снижается при последующем вызывании (вниз) и снова увеличивается при повторном вызывании (вверх). Было установлено, что обезьян можно научить реверсировать измененную реакцию, т. е. проявлять адаптационную пластичность. Наиболее важным открытием было то, что даже после полного рассечения спинного мозга выше пояснично-крестцового участка проводящего пути рефлекса спинального растяжения (что устраняло влияние головного мозга) у животных, которых подвергали выработке оперантного условного рефлекса, по-прежнему наблюдался выработанный рефлекс. Это исследование подтвердило гипотезу о том, что измененная рефлекторная активность со временем модифицирует спинной мозг (Wolpaw, Lee, Carp, 1991). Участок пластичных изменений. По мнению ученых, в настоящее время существует три возможных местонахождения следов памяти спинного мозга, обусловливающих изменения степени спинального рефлекса 123 Наука о гибкости Контроль (дни 1-40) День 40 Рефлекс спинального растяжения (дни 41-70) День 65 Рефлекс спинального 1 растяжения (дни 71-306) День 188 Рефлекс спинального | растяжения (дни 307-393) День 371 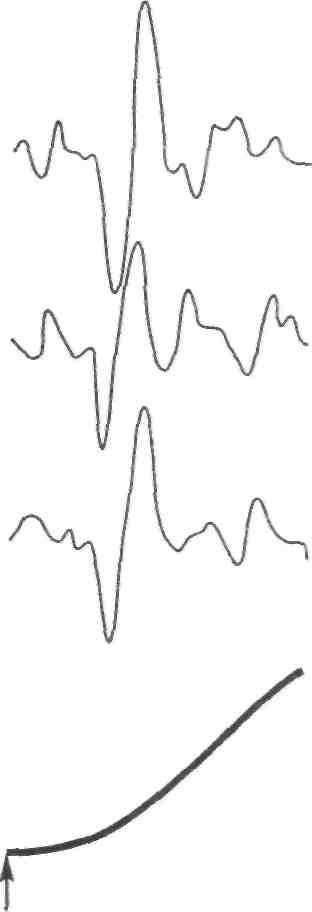 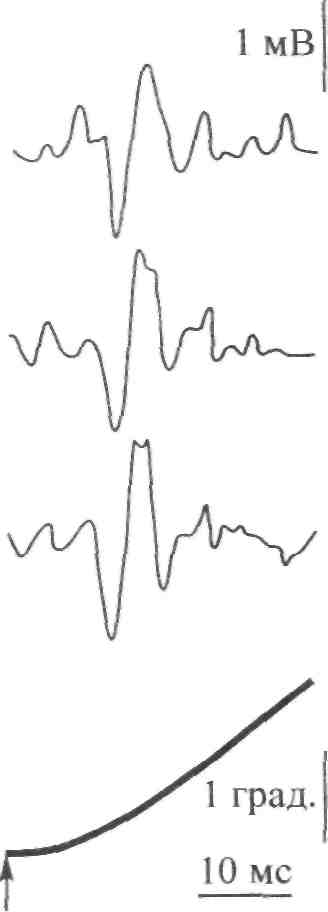 Выпрямление руки в локтевом суставе Пульс в покое Л^АД/ЛЛлД/V Рис. 6.9. Серии отдельных проб ЭМГ обезьяны в условиях контрольного режима — слева; после первоначального вызывания рефлекса спинального растяжения (вверх), после вызывания рефлекса спинального растяжения (вниз), и наконец, после повторного вызывания рефлекса (вверх) — справа (Wolpaw, 1983)растяжения (рис. 6.10). В каждом случае изменение нисходящего влияния фазы I на участок проводящего пути рефлекса спинального растяжения (помечен стрелкой) со временем вызывает изменение фазы II (то есть следа памяти в участке). Наиболее вероятным местом является терминал аф-ферента 1а на мотонейроне. След памяти здесь может быть произведен в результате длительного изменения пресинаптического торможения (рис. 6.10, а). Известно, что передача через 1а синапс ингибируется преси-наптическими импульсами, поступающими из ряда супраспинальных участков (Baldissora, Hultborn, Illert, 1981). Результаты недавних исследований показывают, что кратковременные изменения в пресинаптическом ингибировании играют важную роль в двигательном поведении (Capaday, Stein, 1987a, б). Таким образом, хроническое или длительное изменение этого торможения может привести к модификации терминала 1а. Вольпау и Карп (1990) высказали предположение, что пресинаптическое ингибиро-вание может влиять на обусловленное деполяризацией проникновение кальция и выделение трансмиттера и тем самым изменять величину возбудительного постсинаптического потенциала, производимого в мотонейроне при стимулировании чувствительного афферента 1а. 124 Глава 6. Нейрофизиология гибкости: невральная анатомия и физиологияб Целый мотонейрон в Локальный постсинапти- ческий Пресинапти-ческий о о о* Фаза II Рис. 6.10. Возможное нахождение следов памяти спинного мозга, обусловливающих изменения амплитуды рефлекса спинального растяжения, или Н-рефлекса (Wolpaw and Carp, 1990) В 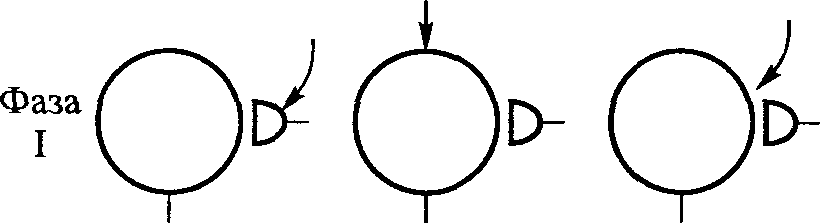 торым возможным источником может быть след памяти, произведенный в результате продолжительного преобразования в мотонейроне, вызывающего изменение его реакции на любой импульс. К примеру, могут измениться свойства мембраны мотонейрона, контролирующие потенциал покоя и сопротивление импульсу, такие, как проницаемость ионов. Вполне понятно, что модификация свойств мембран мотонейрона оказывает влияние на его функцию и помогает определить его реакцию на любой импульс (рис. 6.10, б). торым возможным источником может быть след памяти, произведенный в результате продолжительного преобразования в мотонейроне, вызывающего изменение его реакции на любой импульс. К примеру, могут измениться свойства мембраны мотонейрона, контролирующие потенциал покоя и сопротивление импульсу, такие, как проницаемость ионов. Вполне понятно, что модификация свойств мембран мотонейрона оказывает влияние на его функцию и помогает определить его реакцию на любой импульс (рис. 6.10, б).очень локализованная постсинаптичес- И наконец, третий фактор -кая модификация. Этот процесс, вероятно, может проявляться в изменении рецепторной чувствительности или архитектуры дендритов. Проводящие пути, которые могут вызвать такую избирательную модификацию, в настоящее время не известны (рис. 6.10, в). Клинические перспективы невралъной пластичности С клинической точки зрения представляет интерес вопрос возможности экспериментальной выработки рефлекса спинального растяжения у человека. Согласно Вулфу и Сигалу (1990), данные ряда исследований подтверждают возможность воздействия на нервную систему человека путем мониторинга и обратную связь рефлекса спинального растяжения (Evatt, Wolf, Segal, 1989). Взаимосвязь между невралъной пластичностью и усвоением двигательных навыков Изменения рефлекса спинального растяжения наблюдаются на протяжении всей жизни человека. У детей постепенные изменения рефлекса спинального растяжения происходят в результате усвоения основных двигательных навыков (Myklebust, Gottlieb, Agarwal, 1986). На основании анализа литературы Вольпау, Ли Карп (1991) высказали предположение, что сопоставимые изменения имеют место и на более поздних этапах во время усвоения двигательных навыков, связанных с занятиями балетом (Goode, Van Hoven, 1982), аэробными и анаэробными видами деятельности (Rochcongav, Dassonville, Le Bars, 1979). Таким образом, «медленные, обусловленные физической активностью изменения в спинном мозгу и в каком-либо другом участке ЦНС могут определить в значительной мере процесс усвоения, а 125  Наука о гибкости Наука о гибкоститакже объяснить, почему приобретение многих умений и навыков требует длительной практики» (Wolpaw, Lee и Carp, 1991). Этот факт имеет большое значение для спорта, эргономики, медицины, физиотерапии и т. д. Выявление пластичности спинного мозга имеет особое значение в спортивной медицине. Например, аномальные структуры походки после травмы могут быть причиной болевых ощущений в области стопы, колена, бедра и спины (Day, Wildermuth, 1988; Subotnick, 1979). Более того, эти изменения могут вызвать пластические трансформации в спинном мозгу, затрудняющие и задерживающие полное восстановление. Исследования показывают, что около 33 % больных с хроническими растяжениями имеют остаточные симптомы в течение длительного времени после завершения реабилитации (Bosien, Staples, Russel, 1955; и др., 1982). Было высказано предположение, что последующая функциональная нестабильность может быть вызвана потерей или невосстановлением проприоцепции суставов, конечностей и тела (Freeman, Dean, Hanham, 1965). Вполне возможно, что эта потеря отчасти является следствием пластических изменений в спинном мозгу. Влияние методов растяжения на невральную пластичность Поскольку различные повреждения тела могут привести к пластическим изменениям в спинном мозгу, возникает закономерный вопрос: какие клинические методы в отдельности или в сочетании могут быстро и достаточно эффективно откорректировать физические изменения спинного мозга. Необходимо также изучить возможное влияние на этот процесс различных медикаментозных средств. Взаимосвязь невралъной пластичности со спортом и спортивной реабилитацией В спорте известно, что процесс корректировки неправильно усвоенного требует вдвое больше времени, чем процесс усвоения. Таким образом, при усвоении неправильной техники могут произойти невральные пластические изменения. Поэтому в процессе корректировки нежелательную двигательную структуру предстоит «забыть», чтобы соответствующим образом модифицировать пластические изменения, имевшие место во время начального усвоения. НЕВРОЛОГИЧЕСКИЕ И ДРУГИЕ ФАКТОРЫ, СВЯЗАННЫЕ С ТРЕНИРОВКОЙ ГИБКОСТИ Считается, что прирост мышечной силы обусловлен двумя основными факторами. В то время как мышечная гипертрофия (увеличение размера) имеет место на более поздних этапах тренировочного процесса (Enoka, 1988; Komi, 1986; Sale, 1986), прирост силы, достигаемый в пер- 126 Глава 6. Нейрофизиология гибкости: невральная анатомия и физиология Улучшение соотношения E/F Никаких изменений в активации  Сила б НФ= о—о До тренировок *fПовышенная Соотношение ** E/F не изменилось /  |активация |активация>* х- -х После тренировок В-А С-А МГ = х 100 %    Рис. 6.11. Вклад невральных факторов (а) и мышечной гипертрофии (б) в прирост мышечной силы вследствие тренировочных занятий силовой направленности; оценка вклада (в) невральных факторов (НФ) и мышечной гипертофии (МГ), % (Moritani, de Vries, 1979) Рис. 6.11. Вклад невральных факторов (а) и мышечной гипертрофии (б) в прирост мышечной силы вследствие тренировочных занятий силовой направленности; оценка вклада (в) невральных факторов (НФ) и мышечной гипертофии (МГ), % (Moritani, de Vries, 1979)вые недели тренировочных занятий, отражает повышенную способность активации мотонейронов и, следовательно, имеет невральное происхождение (рис. 6.11). Результаты многочисленных исследований указывают на то, что вскоре после начала тренировочных занятий силовой направленности происходят невральные изменения. Произвольная сила быстро увеличивается прежде, чем мышцы гипертрофируются (Ikai, Fukunaga, 1970; D.A.Jones, Rutherford, 1987; Tesoh, Hjort, Balldin, 1983) и произойдет увеличение напряжения, обусловленное действием электрических сил (Davies, Young, 1983). Этот начальный прирост силы сопровождается увеличением интегрированной ЭМГ (Komi, 1986; Sale, 1986) и интенсификацией рефлексов (Sale и др., 1982; Sale, Upton, McComas, MacDougall, 1983). Что же обусловливает начальное увеличение уровня гибкости в первые недели тренировочных занятий? Влияет ли тип тренировочных занятий (баллистические, статические и т.п.) на сущность этих изменений? Исследования показывают, что у танцоров по сравнению с обычными людьми некоторые рефлексы подвергаются модификации (Goode и Van Hoven, 1982; Nielsen, Crone, Hultborn, 1993). Однако это может быть не связано с невральными факторами. По мнению Кочея, Бурк и Кемен (1991), продолжительные тренировки могут привести к изменению состава соединительного сухожилия, что может повлечь уменьшение нагрузки на аппарат нервно-мышечного веретена. Бирду (1973) удалось показать, что тренировочные занятия аэробной направленности вызывают меньшую степень напряжения на единицу площади поперечного сечения в сухожилиях хвоста крыс. Этот факт указывает на большую фракцию растворимого коллагена. Виидику (1973) удалось продемонстрировать, что растянутое сухожилие проявляет тенденцию оставаться в таком состоянии. Следовательно, любая последующая нагрузка на сухожилие приведет к передаче в мышцу меньшей силы. 127   Наука о гибкости Наука о гибкостиЧто касается поставленного нами вопроса, то ему было посвящено всего одно исследование. Стивене с коллегами (1974) протестировали 232 студента (физвоспитание), выявив 15 испытуемых, имевших наиболее податливую группу мышц подколенных сухожилий, и 15 испытуемых с наиболее жесткой группой. Они обнаружили более мощную ЭМГ-активацию в структурах растяжения подколенных сухожилий и других мышц, а также более раннее начало (последние 40° растягивающего движения) и более продолжительную активность рефлекса мышечного растяжения у лиц с более жесткими группами мышц. У испытуемых с более податливой группой мышц активация рефлекса растяжения приходилась на последние 20° растягивающего движения. Эти данные «говорят в пользу гипотезы о более высокой чувствительности нервно-мышечных веретен, большей возбудимости, направленной вниз цепи веретен, или о повышенной гамма-активности у лиц с более жесткой группой мышц». В другом исследовании Стивене с коллегами (1977) использовали вибрацию (чтобы вызвать тонический вибрационный рефлекс), действующую на сухожилие двуглавой мышцы плеча в течение 2 мин с последующим растяжением. Средняя интегрированная ЭМГ в группах существенно не отличалась. Вместе с тем «... только у лиц с более жесткими группами мышц повторяющиеся растягивающие движения вызывали снижение градиента максимального угла и момент начала растягивающей деятельности (т.е. рефлекс растяжения начинался позже как функция повторяющихся растягивающих движений)». По мнению ученых, эти результаты «могут указывать на то, что упру-говязкие компоненты жесткой мышцы изменяются в результате растягивания, тогда как неврологические компоненты остаются без изменений». У более гибких испытуемых подобные тенденции не наблюдались. Следует упомянуть еще о двух недавно проведенных исследованиях. Вуйнович и Доусон (1994) наблюдали значительное влияние пассивного мышечного растяжения на снижение активности нейронов в спинальном сегменте L5-S1 в результате как статического, так и баллистического растяжения, что коррелирует с более высоким уровнем гибкости. Хальбертсма и Гекен (1994) проанализировали гипотезу, согласно которой упражнения на растягивание удлиняют подколенные сухожилия, изменяя эластичность мышц. Результаты показали, что небольшое, но значимое увеличение растяжимости подколенных сухожилий сопровождалось значительным увеличением растягивающей силы, которую могли выдержать пассивные мышцы подколенных сухожилий. Уровень эластичности, однако, остался таким же. Был сделан вывод, что «упражнения на растягивание не удлиняют подколенные сухожилия и не делают их менее жесткими, а только влияют на толерантность к растяжению». Применение терапевтического растягивания способствует увеличению амплитуды движения. Это объясняется механическим удлинением мышцы и соединительной ткани, а также снижением уровней нейронной 128  Глава 6. Нейрофизиология гибкости: невралъная анатомия и физиология Глава 6. Нейрофизиология гибкости: невралъная анатомия и физиологияактивности. Необходимо провести дополнительные исследования, чтобы определить, в какой степени оно модифицирует упруговязкий компонент мягких тканей и невральные реакции у различных групп населения, а также выявить наиболее эффективные методы, обеспечивающие необходимые результаты. ПЛАНЫ НА БУДУЩЕЕ Нервная система играет ключевую роль в определении качества и количества движения человека. В настоящее время, и особенно в последние годы, наши представления о нервной системе стали более глубокими. Появление новых технологий, быть может, даст нам возможность в ближайшие 10 лет получить информации больше, чем за прошедшие 100 лет. Это позволит специалистам, занимающимся проблемами развития и сохранения оптимального уровня гибкости, создать новые, более эффективные методы, которые существенно повысят качество жизни. РЕЗЮМЕ Структурной и функциональной единицей нервной системы является нейрон. Двумя главными механорецепторами являются нервно-мышечные и нервно-сухожильные веретена. Основными рецепторами растяжения в мышце являются нервно-мышечные веретена. С другой стороны, нервно-сухожильные веретена — главные чувствительные к сокращению механо-рецепторы. Суставные рецепторы, ощущающие действующие на суставы механические силы, называются суставными механорецепторами. На основании морфологических и бихевиоральных критериев эти нервные окончания можно разделить на четыре группы. В основе деятельности нервной системы лежат сложные реакции, называемые рефлексами (например, рефлекс растяжения мышцы, рецип-рокное торможение и обратный рефлекс растяжения). Проведенные в последние годы исследования показывают, что нейронная активность может вызвать стабильные изменения в ЦНС, которые называются пластическими изменениями. Последние имеют практическое значение для процесса реабилитации, усвоения двигательных умений и навыков и развития гибкости. 6-11;  |