Соотношение длина-напряжение и нагрузка-деформация. Длина мягкой ткани зависит от отношения внутренней силы, развиваемой тканью, ко внешней силе, обусловленной сопротивлением развитию внутренней силы или нагрузкой. Если внутренняя сила превышает внешнюю, ткань сокращается. Если же внешняя сила превышает внутреннюю, ткань удлиняется.
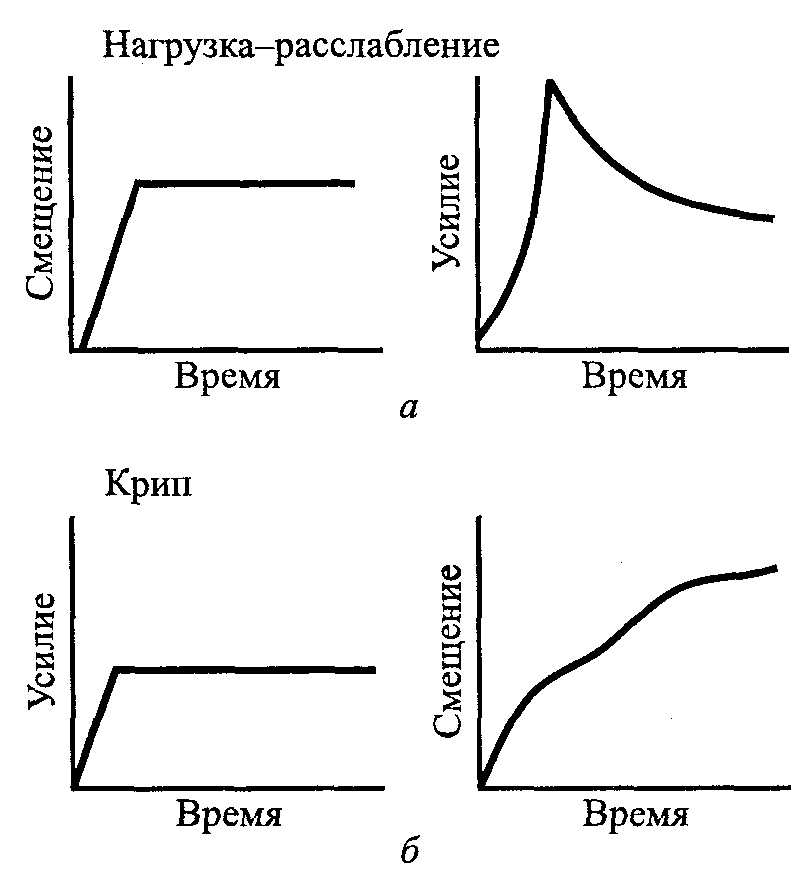
Нагрузка-расслабление и крип при пассивном напряжении. Живые ткани характеризуются наличием зависимых от времени механических свойств. К ним относятся нагрузка-расслабление и крип. Если находящуюся в состоянии покоя мышцу внезапно растянуть и постоянно удерживать достигнутую длину, то через некоторое время произойдет медленное снижение напряжения. Это поведение называют нагрузка-расслабление (рис. 5.8, а). С другой стороны, удлинение, которое происходит при воздействии постоянной силы или нагрузки, называется крип (рис. 5.8, б).
Каким же образом эти зависимые от времени механические свойства действуют на мышечные клетки и соединительные ткани. Несомненный интерес представляют следующие вопросы:
Как передается растягивающая сила через саркомер и структуры различных соединительных тканей?
Как влияет растягивающая сила на сарколемму, саркоплазму и цитос-келет саркомера?
Где и через какие структуры саркомера осуществляется явление крип и нагрузка-расслабление?
6,,
81
Наука о гибкости
• Каково взаимоотношение (если таковое существует) между крипом и нагрузкой-расслаблением в саркомере и градиентами давления, потоком жидкости и потенциалами течения структур различных соединительных тканей?
Рис. 5.8. Реакция тканей на усилие: нагрузка-расслабление имеет место при снижении усилия в случае, если длина ткани сохраняется постоянной (а); крип представляет собой удлинение, имеющее место при действии постоянной силы в течение периода времени (б)
Молекулярный механизм эластичной реакции соединительной ткани. Соединительные ткани представляют собой сложные материалы, которые, соединяясь, образуют длинные гибкие цепочки. Двумя важнейшими переменными, влияющими на жесткость (или эластичность) соединительных тканей, являются расстояние между поперечными соединениями и температура. Представим, например, длинную гибкую молекулу, состоящую из определенного числа сегментов. Количество сегментов обозначим буквой п. Каждый сегмент имеет определенную длину, обозначенную буквой а. Допустим, что каждый сегмент является жестким, тогда как суставы между сегментами — гибкими. Также допустим, что молекулы сегментов свободно передвигаются. Все молекулы двигаются сравнительно хаотично. Однако при снижении температуры их движение становится не таким свободным. При достижении температуры абсолютного нуля (-273° С) движение прекращается. Вследствие хаотичного движения молекул в определенный момент расстояние от одного конца сегмента до другого может иметь значение от О (если концы соприкасаются) до па (если молекулы вытянуты). Наиболее вероятная длина молекулы равна п1/2а.В «нормальном» состоянии молекулярные цепочки сети продолжают двигаться. Расстояние между концами конкретной цепочки изменяется, однако среднее расстояние в образце, содержащем много цепочек, всегда будет п1/2а.Рассмотрим рис. 5.9. Допустим, что на соединительную ткань действует внешняя растягивающая сила (5.9, а). Сетка подвергнется деформации (рис. 5.9, б), и цепочки расположатся в направлении растяжения. Следовательно, цепочки, расположенные в направлении растягивающей силы (например, АВ), будут иметь среднюю длину больше п"2а. Цепочки же, расположенные поперек направления растяжения (ВС), будут иметь среднюю длину меньше п"2а. В результате этого расположение уже не является хаотичным. После устранения действия силы цепочки снова принима- 82
     
Глава 5 ■ Механические и динамические свойства мягких тканей
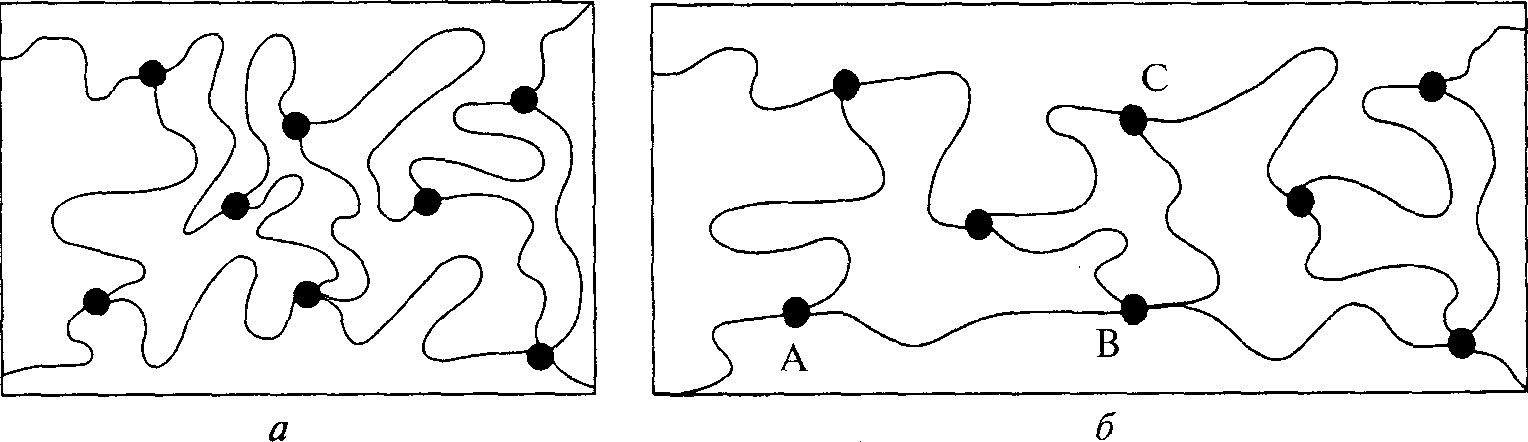
Рис. 5.9. Диаграмма резинового полимера. Синусоидой показаны молекулы полимера, точками — поперечные соединения (Alexander, 1988)
ют хаотичную конфигурацию. Таким образом, соединительная ткань восстанавливает свою исходную форму; она эластично возвращается к исходному уровню.
Р.М.Александер (1988) пишет:
«Теория, созданная на основании этих идей, позволяет определить величину силы, необходимую для уравновешивания деформированной сети и, следовательно, модуля упругости. Модуль сдвига G и модуль Юнга Е можно получить из уравнения
G = NkT = E/3,
где N — количество цепочек на единицу объема материала; к — постоянная Больцмана; Т — абсолютная температура. Особую роль играет число цепочек. Если имеется большее число поперечных соединений, разделяющих молекулы на множество более коротких цепочек, жесткость материала увеличивается. Кроме того, модуль пропорционален абсолютной температуре, поскольку энергия, связанная со скручиванием (сплетением) молекул, увеличивается при повышении температуры. Также по мере повышения температуры увеличивается давление газа при постоянном объеме, так как при этом увеличивается количество кинетической энергии молекул».
Данные исследований, касающиеся растягивания соединительной ткани. При воздействии растягивающей силы на соединительную ткань или мышцу ее длина увеличивается, а площадь поперечного сечения (ширина) уменьшается. Существуют ли такие виды сил или состояний, при которых прилагаемая сила может обеспечить оптимальное изменение соединительной ткани? Сапега с коллегами (1981) отмечают следующее:
«При непрерывном воздействии растягивающих сил на модель организованной соединительной ткани (сухожилие) время, в течение которого происходит необходимое растягивание ткани, обратно пропорционально прилагаемым силам (C.G.Warren,
83
   Наука о гибкости Наука о гибкости
Lehmann, Koblanski, 1971,1976). Таким образом, при использовании метода растягивания с небольшой силой требуется больше времени, чтобы достичь такой же степени удлинения, как при использовании метода растягивания с большой силой. Однако процент удлинения ткани, который имеет место после устранения растягивающего усилия, оказывается выше при использовании продолжительного метода с небольшой силой (C.G.Warren и др., 1971, 1976). Кратковременное растягивание с большой силой способствует восстанавливающейся деформации эластичной ткани, тогда как продолжительное растягивание с небольшой силой —; остаточной, пластической деформации (С. G. Warren и др., 1971, 1976; Labon, 1962). Результаты лабораторных исследований показывают, что при постоянном удлинении структур соединительной ткани имеет место определенное механическое ослабление, хотя разрыв и не происходит (C.G.Warren и др., 1971, 1976). Степень ослабления зависит от способа растягивания ткани, а также от степени растяжения.
Температура существенно влияет на механическое поведение соединительной ткани в условиях растягивающего напряжения. При повышении температуры ткани степень жесткости уменьшается, а степень растяжимости увеличивается (Laban, 1962; Rigby, 1964). Если температура сухожилия превышает 103°Ф, количество постоянного удлинения в результате данного количества исходного растягивания увеличивается (Laban, 1962; Lehmann, Masock, Warren u Koblanski, 1970). При температуре около 104° Ф происходит термальное изменение микроструктуры коллагена, которое значительно усиливает расслабление вязкости после нагрузки коллагеновои ткани, что обеспечивает более высокую пластическую деформацию при растягивании (Mason и Rigby, 1963). Механизм, лежащий в основе этого термального изменения, пока не известен, однако предполагают, что происходит частичная дестабилизация межмолекулярной связи, усиливающая вязкие свойства текучести коллагеновои ткани (Rigby, 1964).
Если соединительную ткань растягивают при повышенной температуре, условия, в которых ткань может охладиться, в значительной мере могут повлиять на качество удлинения, которое остается после устранения действия растягивающего напряжения. После растяжения разогретой ткани сохраняющаяся растягивающая сила во время охлаждения ткани значительно увеличивает относительную пропорцию пластической деформации по сравнению с разгрузкой ткани при все еще повышенной температуре (Lehmann и др., 1970). Охлаждение ткани до устранения напряжения позволяет коллагеновои микроструктуре больше рестабилизироваться к ее новой длине (Lehmann и др., 1970).
84
     Глава 5- Механические и динамические свойства мягких тканей Глава 5- Механические и динамические свойства мягких тканей
При растягивании соединительной ткани при температурах, находящихся в обычных терапевтических пределах (102-110° Ф), количество структурного ослабления, обусловленное данным количеством удлинения ткани, обратно пропорционально температуре (C.G.Warren и др., 1971, 1976). Это явно связано с прогрессивным увеличением свойств вязкого течения коллагена по мере увеличения температуры. Вполне возможно, что термальная дестабилизация межмолекулярной связи обеспечивает удлинение при меньшем структурном повреждении.
Факторы, влияющие на упруговязкое поведение соединительной ткани, можно обобщить, отметив, что эластичной, или обратимой, деформации больше всего способствует кратковременное растягивание с большой силой при нормальной или несколько пониженной температуре тканей, тогда как пластическому, или постоянному, удлинению в большей степени способствует более длительное растягивание с меньшей силой при повышенной температуре, если только охлаждение ткани осуществляется до устранения напряжения. Кроме того, структурное ослабление, обусловленное остаточной деформацией ткани, оказывается минимальным, когда продолжительное воздействие небольшой силы сочетают с высокими температурами, и максимальным — при использовании больших сил и более низких температур. Эти данные обобщены в табл. 5.1-5.3».
Исследования, проводившиеся другими учеными (Becker, 1979; Glarer, 1980; Light и др., 1984), также показывают, что растягивание при низких-средних уровнях напряжения, является действительно эффективным.
Таблица 5.1. Факторы, влияющие на пропорцию пластичного и эластичного растяжения
Фактор
Эластичное растяжение
Пластичное растяжение
 Количество прикладываемого усилия Высокое усилие Невысокое усилие Продолжительность прикладываемого Небольшая Большая усилия Температура ткани Низкая ВысокаяТаблица 5.2. Факторы, влияющие на упруговязкое поведение соединительной ткани
 Эластичная деформация Эластичная деформация
Вязкая (пластическая) деформация
 Кратковременное растягивание Продолжительное растягивание с большим усилиемс небольшим усилием Нормальная или несколько Повышенная температура, охлаждение пониженная температура тканей осуществляется до устранения напряжения 85

МЫШЦА
Мы достаточно много знаем о различных механических свойствах мышц, так как их интенсивно исследовали многие ученые на протяжении не одного десятилетия. Существует множество причин, ввиду которых необходимо изучать механические свойства мышцы. Прежде всего, таким образом мы можем лучше понять как механические реакции всей мышцы, так и механические свойства сократительных компонентов (Zierler, 1974). В данном контексте необходимость изучения механических свойств мышцы объясняется тем, что таким образом мы можем понять и определить факторы, ограничивающие гибкость, а также лучшие способы ее увеличения.
Соединительные ткани мышц. Мышца состоит из трех независимых компонентов или элементов, которые можно классифицировать либо как эластичные, либо как вязкие. Важность этих компонентов объясняется тем, что они противодействуют деформации и, таким образом, играют важную роль в определении уровня гибкости человека. Эластичные компоненты в ответ на изменение длины производят возвращающую силу. Вязкие компоненты производят усилие в ответ на интенсивность (скорость) и продолжительность изменения длины. Тремя механическими компонентами являются:
Параллельный эластичный компонент (ПЭК).
Серийный эластичный компонент (СЭК).
Сократительный компонент (СК).
Параллельный эластичный компонент
Компонент, отвечающий за пассивное растягивающее напряжение в мышце, называется параллельным эластичным компонентом (ПЭК). Его название объясняется тем, что он располагается параллельно сократительному механизму (рис. 5.10). Если мышцу удалить из тела, то ее длина сок-
86
  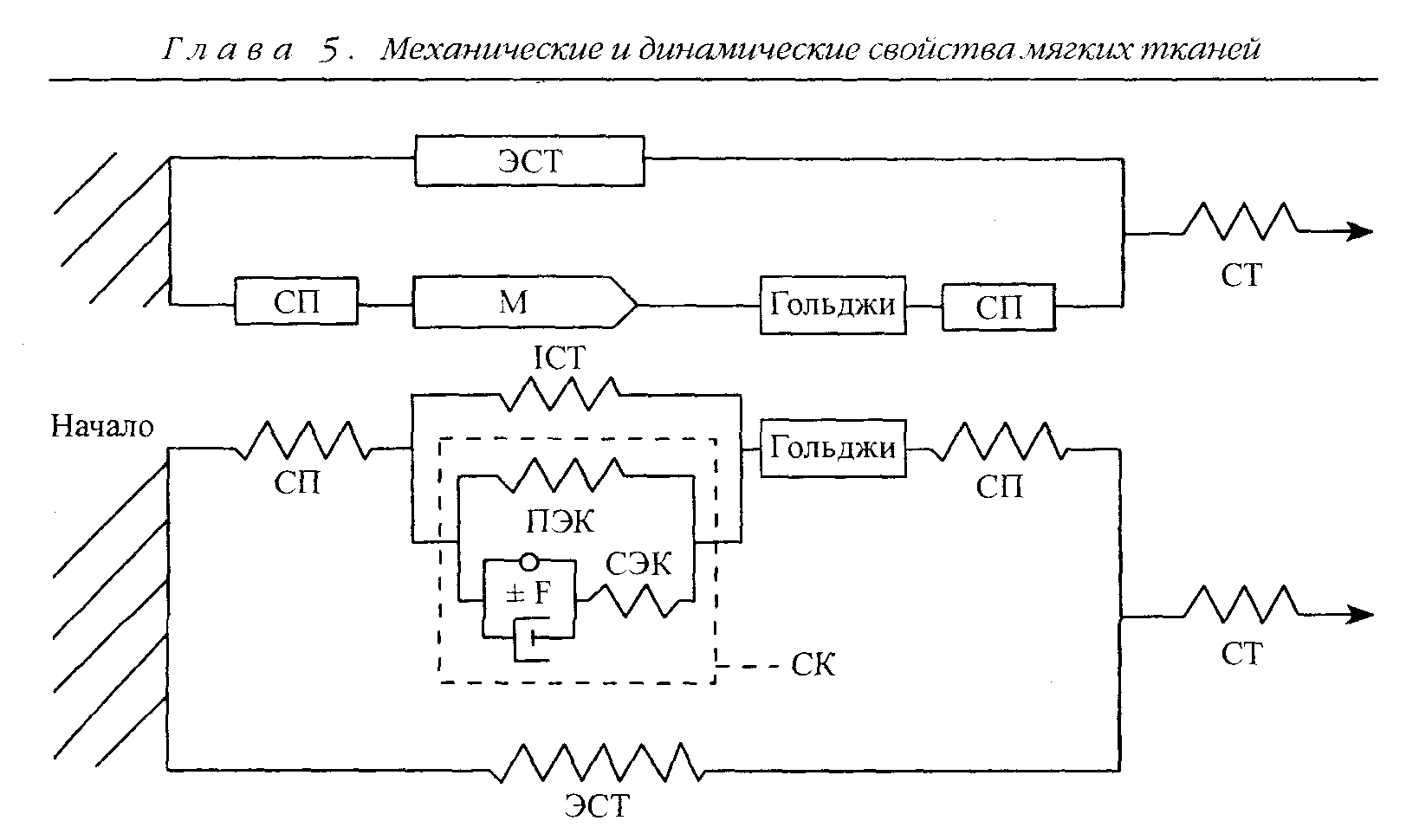
Рис. 5.10. Диаграмма упруговязких компонентов мышечного волокна и окружающей соединительной ткани с нервно-сухожильным веретеном. СП — сухожильные пучки; ЭСТ — экстрасоединительная ткань; М — мышечное волокно; Гольджи — нервно-сухожильное веретено; ПЭК, СК, СЭК, ICT — внутримышечная соединительная ткань; СТ — сухожилие
ратится примерно на 10 % по сравнению с первоначальной (Garamvolgyi, 1971). Такое сокращение длины мышцы не зависит от пассивного сокращения. Длина изолированной, несокращенной мышцы называется ее длиной равновесия; это укорачивание означает, что мышцы находятся под напряжением при нетронутой длине. Длина in situ несокращенной или нерастянутой мышцы называется длиной покоя и обозначается R1 или L0.
Находящаяся в покое мышца эластичная и противодействует удлинению. При длине, уступающей длине равновесия (0,90 L0), напряжение покоя отсутствует и ПЭК слабый. Однако когда нестимулированную мышцу растягивают, она развивает напряжение нелинейным образом. Другими словами, при начальном растягивании производится невысокое напряжение, которое по мере продолжения растягивания значительно увеличивается.
Первоначально считали, что ПЭК состоит, главным образом, из сарколеммы, саркоплазмы и эластичных волокон — эпимизия, перимизия и эн-домизия. Спустя некоторое время Х.Е. Хаксли и Хенсон (1954) выдвинули предположение об S-филаменте, который, как считали, соединял концы ак-тиновых филаментов на обоих концах. Однако уже через год Х.Е. Хаксли (1957) исключил из своей модели мышцы S-филаменты без каких-либо объяснений. Другая трактовка пассивного напряжения покоя была связана с электростатической силой. Например, известно, что объем мышечных волокон остается постоянным даже при растягивании мышцы. Площадь же поперечного сечения (ширина) мышцы должна уменьшиться, как и расстояние между филаметами актина и миозина, по мере их приближения друг к другу. Однако если между филаментами существует электростатическая сила взаимного отталкивания, то, чтобы сблизить филаменты, дол-
87
 Наука о гибкости Наука о гибкости
жна быть выполнена работа. Следовательно, должна существовать сила, способствующая сохранению расположения филаментов. Таким образом, сила, необходимая для сближения филаментов, преодолевающая силу взаимного отталкивания, будет представлена напряжением покоя или «параллельным» сопротивлением растяжению (Davson, 1970; Н.Е. Huxley, 1967). Хотя электростатическая сила может способствовать напряжению покоя при высокой степени растягивания, исследования показывают, что она не может быть доминирующим источником этого процесса.
Как мы уже знаем из главы 3, основным источником эластичности мышц является титин. Это доказательство было получено в результате разрушения филаментов титина во время регистрации степени напряжения мышцы под нагрузкой. В первом исследовании титин преимущественно разрушили радиацией (Horowitz и др., 1986). Результатом оказалось снижение напряжения покоя. Год спустя Горовитц и Подольски (19876) опубликовали данные другого исследования, подтверждающие гипотезу, что эластичные филаменты титина обеспечивают большую часть напряжения покоя в мышце. В другом исследовании (Yoshioka и др., 1986) титин преимущественно разрушали путем контролируемого протеолиза, использования гидролитических (пищеварительных) ферментов. Снова отмечали снижение напряжения покоя. Еще в одном, более позднем исследовании (Funatsu, Higuchi, Ishiwata, 1990) также наблюдали снижение напряжения покоя при деградации титина в результате ферментного сжигания (гезолином плазмы).
Что же происходит, когда саркомер растягивается и отпускается? Как было показано в главе 3, длина актинового тонкого и толстого (миозинового) филаментов во время растягивания не изменяется. Они просто скользят относительно друг друга (теория скольжения филаментов). При растягивании саркомер противодействует силе деформации напряжением покоя. Вначале напряжение покоя среднее. После значительного растягивания оно резко увеличивается и противодействует последующему растяжению (эластичная жесткость). Ответственным за такое поведение является титин. После прекращения растягивания растянутые филаменты титина возвращаются в исходное положение. Таким образом, титин может накапливать потенциальную энергию.
Если титин способствует напряжению покоя, то что можно сказать о небулине? Исследования показывают, что небулин представляет собой набор нерастяжимых филаментов, прикрепленных к одному концу Z-линии, и что его филаметы идут параллельно филаментам титина (Wang, Wright, 1988). Таким образом, небулин может быть потенциальным четвертым фи-ламентом саркомера. Согласно исследованиям, при расщеплении небулина напряжение покоя не снижается (Funatsu, Higuchi, Ishiwata, 1990). Следовательно, небулин не обеспечивает эластичность.
Серийный эластичный компонент
При растягивании мышцы сократительный компонент (т. е. филаменты актина и миозина и их поперечные мостики), ПЭК и СЭК способствуют развитию напряжения. Серийный эластичный компонент (СЭК) полу-
88
 
Глава 5 • Механические и динамические свойства мягких тканей
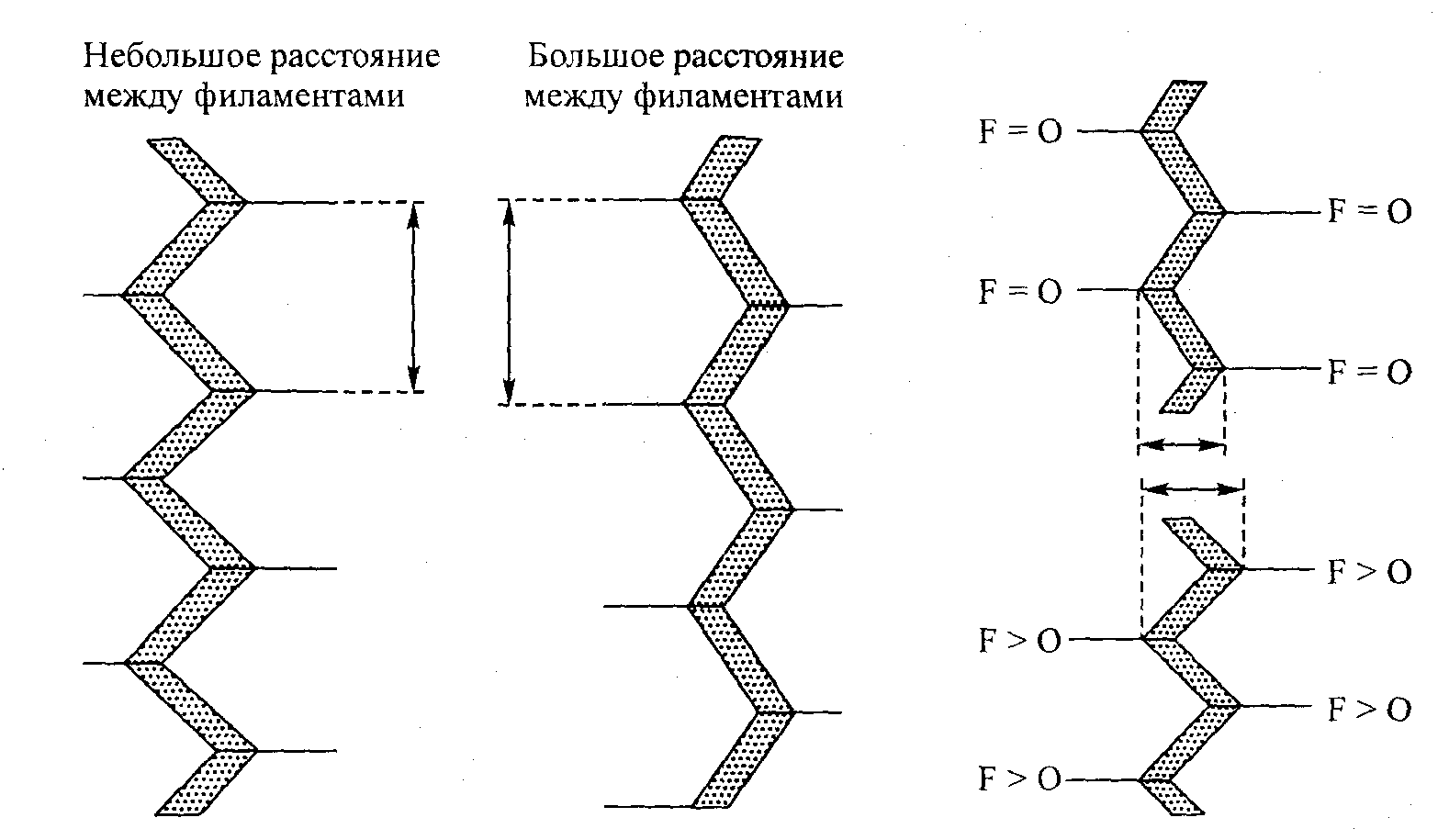
Рис. 5.11. Влияние изменения расстояния между филаментами на структуру Z-линии. Z-линия осуществляет эти изменения, меняя угол между соприкасающимися элементами
Рис. 5.12. Сила, действующая на тонкие филаменты (F), усиливает зигзагообразный паттерн структуры Z-линии, что приводит к изменению эффективной «ширины» последней. Подобные напоминающие гармошку изменения могут обусловить серийную эластичность
чил такое название потому, что эластичные компоненты располагаются непосредственно в соответствии с сократительными компонентами (см. рис. 5.10). СЭК выполняет важную функцию сглаживания резких изменений мышечного напряжения. Одной из главных анатомических частей, образующих СЭК, по мнению специалистов, является сухожилие. Следует отметить, что, по мнению Поллака (1990), Z-линия также может представлять собой определенный источник серийной эластичности саркомера. Эта функция осуществляется тонкими филаментами, которые тянут Z-линию. Например, когда сила, действующая на тонкие филаменты, передается на Z-линию, степень латерального разделения между филаментами снижается (рис. 5.11). Вместе с тем чтобы адаптировать сокращение межфила-ментного пространства, угол изгиба в структуре Z-линии становится более острым (рис. 5.12). Следовательно, Z-линия эффективно утолщается, создавая своеобразную «эластичность».
Сократительный компонент
Способность мышцы увеличивать напряжение называется сократительным компонентом (СК). СК мышцы можно рассматривать как генератор напряжения. Он состоит из филаментов и их поперечных мостиков. Если напряжение пропорционально числу химических связей, установленных между двумя филаментами, то с увеличением степени взаимного перекрытия филаментов большее число участков связывания может взаимо-
89
Наука о гибкости
действовать и большее напряжение может быть развито. Предполагают, что максимальное сократительное напряжение развивается при длине сар-комера, обеспечивающей наиболее полное отдельное перекрытие фила-ментов актина и миозина. При большей длине мышцы число поперечных соединений уменьшается по мере снижения степени перекрытия филамен-тов, что приводит к снижению напряжения. Если продолжать растягивание, развиваемое напряжение может уменьшаться до тех пор, пока оно не превысит напряжение, наблюдаемое в пассивной мышце. Это происходит потому, что при подобной длине филаменты актина и миозина больше не соприкасаются. Таким образом, они в лучшем случае развивают незначительное напряжение.
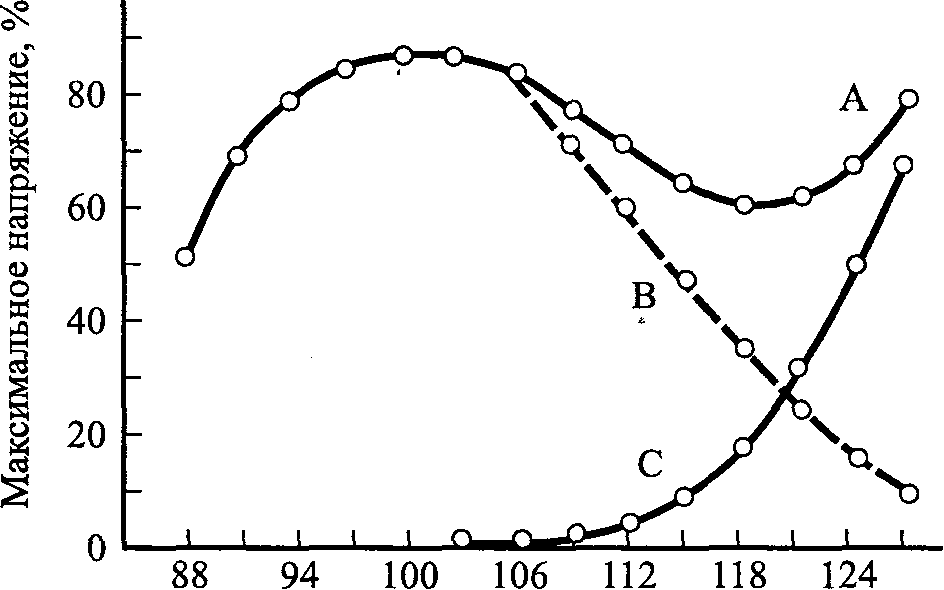
100 -
Общее напряжение активной мышцы во время растягивания. Максимальное общее активное напряжение наблюдается в том случае, когда длина мышцы превышает исходную длину или длину в покое в 1,2-1,3 раза. При большей длине общее активное напряжение снижается до тех пор, пока длина мышцы не превышает ее длину в покое примерно в 1,5 раза; в этом случае производство активного напряжения равно нулю. При длине выше 1,3 L Q количество поперечных соединений снижается, степень перекрытия также сокращается, что приводит к уменьшению напряжения. Более того, хотя ПЭК возрастает в своем производстве пассивного напряжения, этот показатель не соответствует снижению активного напряжения сократительных компонентов. Следовательно, величина общего напряжения снижается. При экстремальной длине мышцы пассивное напряжение, генерируемое СЭК, существенно увеличивается, компенсируя тем самым снижение активного напряжения, что приводит к увеличению общего напряжения. Диаграмма длины-напряжения для пассивного растягивания нестимулированной мышцы показана на нижней кривой С. Кривая А, иллюстрирующая общее изометрическое напряжение при стимулировании мышцы различной длины от максимального растяжения до умеренного сокращения, представляет собой сумму активного сокращения (В) плюс напряжение, обусловленное пассивным растягиванием (С). Активное напряжение (кривая В), обусловленное исключительно мышечным сокращением, можно получить, вычитая величину пассивного напряжения С из величины общего напряжения А. Нормальная длина мышцы в покое 100% (рис. 5.13). Растягивание мыш
цы во время сокраще
ния при большой длине.
Длина в покое, % Ткани при растягивании Рис. 5.13. Диаграмма длины-напряжения общего и развивают напряжение, пассивного напряжения называемое реакцией на 90
     Глава 5 ■ Механические и динамические свойства мягких тканей Глава 5 ■ Механические и динамические свойства мягких тканей
растяжение; оно не зависит от ЦНС и представляет собой механическое свойство растягиваемой ткани. С другой стороны, рефлекс растяжения представляет собой реакцию, обусловленную ЦНС, которая вынуждает растянутую мышцу сокращаться в ответ на стимул растяжения (Gowitzke и Milner, 1988).
Одним из основных аргументов против использования баллистического растягивания является то, что оно вызывает рефлекс растяжения. Однако если выполнять такое растягивание при длине, превышающей 1,5 L0, то рефлекс растяжения не должен привести к увеличению напряжения в СК, поскольку при такой длине филаменты не способны соприкасаться и производить напряжение. Причем это справедливо только в том случае, если все саркомеры мышечного волокна растянуты в одинаковой степени. Но это происходит не всегда. Например, саркомеры, расположенные вблизи сухожилий, растягиваются значительно меньше тех, что находятся в середине мышцы, поэтому они могут проявлять рефлекс напряжения и влиять на степень растяжения. Таким образом, активация рефлекса растяжения, даже если длина мышечного волокна превышает 1,5 L0, по-видимому, приведет к производству дополнительного напряжения СК.
СОСУДИСТАЯ ТКАНЬ
Во время движения и особенно во время растягивания многочисленные структуры тела подвергаются воздействию различных сил. В первую очередь это соединительные ткани (например, сухожилия, связки и фасции) и мышцы. Однако не следует забывать и о двух других категориях структур — сердечно-сосудистой и нервной системах. В следующих разделах мы проанализируем влияние растягивания на эти структуры.
Анатомия сердечно-сосудистой системы. Сердечно-сосудистая система состоит из многообразия кровеносных сосудов, по которым движется кровь через все тело. Существует три основных вида кровеносных сосудов: артерии, вены и капилляры.
Артерия — кровеносный сосуд, по которому кровь идет от сердца. Все артерии (за исключением легочной и ее ответвлений) переносят насыщенную кислородом кровь. Артериальный сосуд, имеющий диаметр меньше 0,5 мм, называется артериолой. В зависимости от превалирующего компонента ткани в стенках артерий их можно разделить на эластичные и мышечные. Эластичными артериями являются такие крупные сосуды, как аорта, сонная, подключичная и подвздошная артерии. Подавляющее большинство артерий относится к категории мышечных. Посредством сужения и расширения артерий осуществляется регуляция кровотока (рис. 5.14).
Вена — кровеносный сосуд, по которому кровь поступает к сердцу. Все вены (за исключением легочной) содержат дезоксигенированную кровь. Небольшие вены называются венудами.. Главное различие между венами и артериями — сравнительная слабость средней оболочки стенок вен. Значительно меньшее содержание в них мышечных и эластичных во-
91
Наука о гибкости

Адвентициальная оболочка
Интима Кровоток
 БОЛЬШИЕ ВЕНЫ БОЛЬШИЕ ВЕНЫ
ВЕНЫ СРЕДНЕГО РАЗМЕРА (диаметр 1-10 мм)
Адвентициальная оболочка
Средняя оболочка
С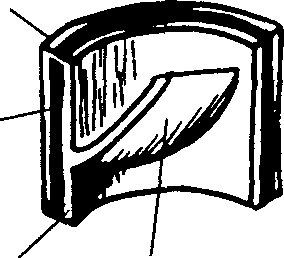 редняя оболочка редняя оболочка
Интима / Капилляры
Клапаны в венах (диаметр >2 мм)
БОЛЬШИЕ АРТЕРИИ (эластичные, проводящие)
А двентициальная оболочка двентициальная оболочка
Средняя ""' ^ Интима оболочка Кровоток
АРТЕРИИ СРЕДНЕГО
РАЗМЕРА (мышечные)
(диаметр 0,3-10 мм)
А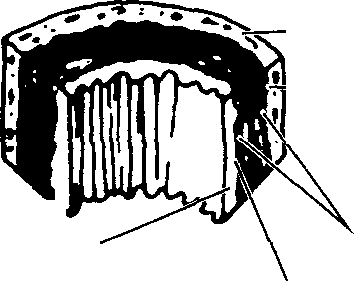 двентициальная двентициальная
- оболочка
_ Внешняя
эластичная
^ AV
Анастомо?
мембрана
Интима
Средняя оболочка Внутренняя эластичная мембрана
Главная артерия
Рис. 5.14. Структурные характеристики основных сегментов кровеносных сосудов
млекопитающих
локон связано с намного более низким давлением венозной крови. Кроме того, в отличие от артерий, большинство вен имеют клапаны.
Капиллярами называются микроскопические кровеносные сосуды, соединяющие артериолы с венулами. Их стенки состоят из одного слоя эндо-телиальных клеток. Средний диаметр капилляров небольшой, от 7 до 9 мкм; длина колеблется от 0,25 до 1 мм. Капилляры выполняют разнообразные функции: транспорт крови со всеми необходимыми ее компонентами и их обмен с окружающими тканями, поддержание нормального давления и кровообращения; кроме того, они являются и резервуаром крови.
Растяжение кровеносных сосудов. Когда кровеносные сосуды подвергаются нагрузке (растяжение кровотоком или влияние мышечного растяжения на кровеносные сосуды), они проявляют типичные упруго-вязкие свойства в ответ на постоянное изменение нагрузки, сопровождающее каждый сердечный цикл.
Влияние длины саркомера скелетной мышцы на общую длину капилляров изучал Эллис с коллегами (1990), использовав для этого длинный разгибатель пальцев крысы. Данные, полученные из шести капилляров, «показали, что четыре (сосуда) оказались растянутыми в такой же степени, как и мышца, один был растянут больше, и один — меньше». Было высказано предположение, что различия в степени растяжимости капилляров могут быть обусловлены различиями в диаметре сосудов, толщине стенок и степени привязывания к соседним мышечным клеткам.
92
  Глава 5 ■ Механические и динамические свойства мягких тканей Глава 5 ■ Механические и динамические свойства мягких тканей


94
 
95
Наука о гибкости
Эпиневрий
Эпиневрий — внешняя фиброзная оболочка нерва из плотной соединительной ткани. Эта наиболее крайняя оболочка окутывает весь нерв и лежит между пучками волокон. Она содержит волокна соединительной ткани, кровеносные сосуды и небольшие нервные волокна, иннервирую-щие сосуды. Компоненты эпиневрия, наиболее заметными из которых являются коллагеновые фибриллы, ориентированы преимущественно продольно. Эпиневрий включает также эластичные волокна.
Периневрии
Периневрии расположен в глубине эпиневрия и отдельно окутывает каждый пучок нервных волокон. Таким образом, каждый пучок окружен периневрием, включающим 3-10 концентрических слоев клеток. Количество слоев зависит от размера нервного пучка и его удаления от ЦНС. Клетки в этих слоях плотно соединяются, превращая периневрии в барьер для прохождения большинства макромолекул. Коллагеновые фибриллы здесь более тонкие, чем в эпиневрий, и содержат меньший процент эластичных волокон.
Эндоневрий
Эндоневрий представляет собой наиболее глубоко расположенную нервную оболочку, окутывающую каждое отдельное нервное волокно. Он состоит из тонкого слоя коллагеновых фибрилл, ориентированных в основном продольно. Диаметр этих фибрилл практически такой же, как фибрилл, находящихся в периневрии.
Растягивание нервов. Исследования поведения периферических нервов, подвергающихся растяжению (растягивающей нагрузке), начали проводить еще в конце XIX ст. Несмотря на это, наши сведения о биомеханических свойствах периферических нервов, а также о пределе растягивания нерва весьма ограничены.
Результаты исследований показывают, что в хронически травмируемых нервах могут изменяться механические свойства, например увеличивается жесткость (Beel, Groswald, Luttges, 1984).
Если нерв подвергать постепенно увеличивающейся растягивающей нагрузке, наблюдается линейная взаимосвязь между нагрузкой и удлинением до определенного момента, когда нерв прекращает вести себя как эластичная структура (Sunderland, 1978, 1991). Основным компонентом, сообщающим эластичность нервному стволу и обусловливающим его предел прочности, является периневрии. Диапазон эластичности составляет 6-20 % по сравнению с длиной в покое.
Если растягивание продолжать после преодоления предела эластичности, прямо пропорциональная взаимосвязь между деформацией и силой исчезает. По мере прикладывания сил большей величины кривая выравни-
96
   Глава5 • Механические и динамические свойства мягких тканей Глава5 • Механические и динамические свойства мягких тканей
вается до уровня максимальной нагрузки или предела прочности, т. е. достижения разрыва.
Данные, касающиеся величины растяжения, вызывающей структурные изменения, являются весьма немногочисленными, а показатели удлинения колеблются от 11 до 100 %. Эти структурные изменения в значительной мере зависят от величины и характера деформирующей силы, а также от продолжительности ее воздействия.
Свойства напряжения — деформации периферического нервного ствола. Сандерленд и Бредли (1961) провели серию экспериментов, изучая явление напряжения-деформации в растянутых периферических нервах человека, подвергавшихся постепенно увеличивающимся нагрузкам вплоть до механического повреждения. У лиц в возрасте 30-50 лет были взяты и сразу же подвергнуты анализу образцы срединного (п=24), локтевого (24), медиального подколенного (13) и латерального подколенного (15) нервов спустя 12 часов после смерти.
Проведенные тесты позволили получить следующую информацию о диапазоне максимальной нагрузки (кг):
срединный 7,3-22,3
локтевой 6,5-15,5
медиальный подколенный 20,6-33,6
латеральный подколенный 11,8-21,4.
Максимальный предел прочности нервного ствола
Нервы не являются гетерогенными структурами и не ведут себя как совершенные цилиндры. Диапазон максимального растягивающего напряжения (кг-мм2) определяли на площади поперечного сечения нервного ствола:
срединный 1,0-3,1
локтевой 1,0-2,2
медиальный подколенный 0,5-1,8
латеральный подколенный 0,8-1,9.
Максимальное удлинение нервного ствола
Если при растягивании нерва не был превышен его предел эластичности, то нерв восстанавливает свою исходную длину. Исследования также показывают, что при устранении нагрузки нерв восстанавливает и свои эластичные свойства. Если же предел эластичности превышен, нерв не восстанавливает свою исходную длину, а оказывается деформированным. Тесты выявляют линейную (эластичную) взаимосвязь между нагрузкой и удлинением в диапазоне удлинения, которую можно представить следующим образом (%):
6-ш
97
Наука о гибкости
срединный 6-20,
локтевой . 8-21,
медиальный подколенный 7-21,
латеральный подколенный 9-22.
Процент удлинения при механическом поврежденииМаксимальное удлинение при пределе эластичности составляет около 20 %. При максимальном удлинении порядка 30 % происходит полный механический разрыв. Удлинение при механическом повреждении нервного ствола как процент от длины в покое составляло (%): срединный 7-30, локтевой 9-26, медиальный подколенный 8-32, латеральный подколенный 10-32. Значение нервного растяжения для специалистов в области медицины. Ридевик с коллегами (1990) выяснили некоторые факторы, имеющие практическое значение для специалистов в области медицины. Во-первых, было установлено, что при механическом повреждении нервный ствол оказывается в целом неповрежденным, несмотря на многочисленные разрывы периневральных оболочек. «Таким образом, чисто визуально невозможно определить структурную целостность нервного ствола». Кроме того, разрывы периневральных оболочек имеют место не в одном участке. Этот факт «указывает на то, что травмы от растяжения периферического нерва могут быть не локальными, а происходить вдоль всей длины нерва» (рис. 5.16). Внутриневральный капиллярный кровоток. Важным последстви ем растягивания нерва является воздействие на внутриневральный капил лярный (микроваскулярный) кровоток (рис. 5.17). При растяжении нерва площадь его поперечного сечения постепенно уменьшается. Это изменение приводит к сжатию, вызывающему дальнейшую де формацию нервного волокна, а также нару шение его кровоснабжения. Важность адекватного кровоснабжения для функции нерва хорошо известна. Поэтому можно ожидать, что растягивание, отрицательно влияющее на внутриневральный капилляр ный кровоток, нарушает нервную функ цию. Исследования, проведенные Лунд- борг (1975), Лундборг и Ридевик (1973) и Рис. 5.16. Поведение пучка и нахо- Огата и Наито (1986), показали нарушение дящихся в нем нервных волокон внуТриневрального капиллярного кровото- нервного ствола, растянутых до п _. „ точки механического повреждения ка ПР И удлинении нерва на 8%. Полная 98
Глава 5 ■ Механические и динамические свойства мягких тканей
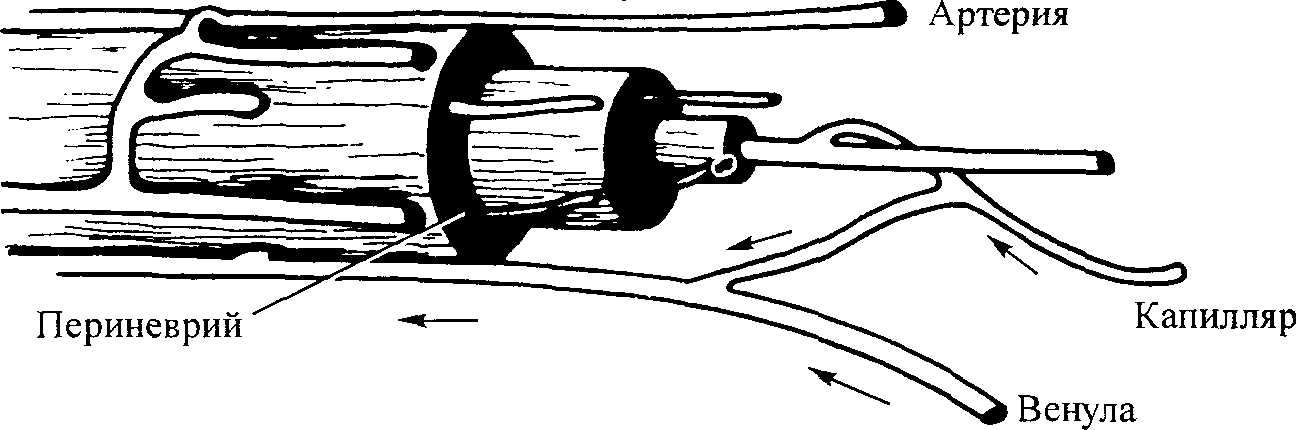   Рис. 5.17. Архитектура микрососудов внутри пучка, определенная в результате исследований периневрия, артериолы, венулы, капилляров. Обратите внимание на капиллярные петли, которые иногда располагаются в плоскостях, перпендикулярных продольной оси нерва. Стрелками отмечено направление кровотока внутриневральная ишемия (снижение кровоснабжения) возникала при удлинении на 15 %. После расслабления, следовавшего за растяжением, кровообращение восстанавливалось. Влияние растягивания на нервную передачу. Другим немаловажным последствием растяжения нервов является нарушение электрической проводимости. Нарушение проводимости наблюдали при растягивании на 6-100 %, в зависимости от вида подопытного животного. Не так давно Уолл с коллегами (1992) высказали предположение, что начальное (раннее) нарушение проводимости обусловлено не ишемией, а механической деформацией. Защитные структуры нервных стволов. Каким образом периферические нервы ног позволяют человеку стоять прямо, наклоняться вперед, не сгибая ноги в коленях, и упираться ладонями в пол? Ведь во время выполнения этих движений ткани, вовлеченные в растягивание, иногда удлиняются до 5 см! Ответ заключается в следующем. Для большинства периферических нервов характерны три особенности, защищающие их от физической деформации: ненатянутость, ход (расположение) нервов относительно суставов и эластичность. Ненатянутость нервного ствола и нервных волоконНервный ствол проходит волнообразно. Такой же волнообразный ход в оболочках эпиневрия характерен и для пучков волокон, а также для каждого нервного волокна внутри пучка. Если напряжение небольшое или отсутствует вообще, нервы сокращаются подобно гармошке (J.W.Smith, 1966). Вследствие этого длина нервного ствола и нервных волокон между любыми двумя фиксированными точками конечности значительно превышает линейное расстояние между этими точками (рис. 5.18). При начальном растягивании волнистость нерва устраняется. По мере продолжения растягивания она исчезает в пучках и, наконец, в отдельных нервных волокнах. Таким образом, только при этом оконча-
7*
99
Наука о гибкости

Пучок.
Нервное_ волокно
Нервный ствол тельно исчезает волнистость и нервные волокна подвергаются напряжению. Если растягивание продолжается, проводимость в нервных волокнах постепенно ухудшается и затем полностью нарушается до тех пор, пока не происходит разрыв нервных волокон внутри пучка. В последнюю очередь структурным повреждениям подвергается пери-неврий.
Рис. 5.18. Диаграмма, иллюстрирующая характерную волнистость нервов, пучков и нервных волокон, которая защищает нервы при их растягивании во время движения конечностей с полной амплитудой
Важность этой волнистой системы трудно переоценить. Как отмечает Сандерленд (1991), «такая волнистость позволяет абсорбировать и нейтрализовать силы тяги, производимые во время движений конечности; таким образом, нервные волокна оказываются постоянно защищенными от перерастяжения». Ход нерва относительно суставовВторым важным свойством, обеспечивающим защиту нервов, является ход, или расположение, нерва относительно суставов. Все нервы, за исключением двух, пересекают сгибательный аспект суставов («внутреннюю часть» сустава, когда он согнут). Поскольку диапазон сгибания сустава намного превышает диапазон выпрямления, нерв, пересекающий сгибательный аспект сустава, остается в расслабленном состоянии в момент сгибания и только немного растягивается при выпрямлении. С другой стороны, нерв, пересекающий выпрямляющий аспект сустава, находится в расслабленном состоянии во время выпрямления и подвергается значительному напряжению во время сгибания. Вполне понятно, что нервы, пересекающие выпрямительный аспект сустава, имеют преимущество с точки зрения воздействия на них сил, генерируемых во время движений конечностей. Исключение составляют локтевой нерв, пересекающий разгибательный аспект локтевого сустава, и седалищный нерв в точке, в которой он пересекает разгибательный аспект тазобедренного сустава. Вследствие этого оба нерва периодически подвергаются чрезмерному напряжению при полном сгибании. Так как движения, включающие сгибание туловища вперед при выпрямленных коленях, встречаются во многих видах спорта и физической деятельности, этот факт, безусловно, заслуживает внимания. Сандерленд (1991) указывает, что в месте пересечения седалищным нервом разгибательной части тазобедренного сустава эпиневральная ткань 100
  Глава 5 • Механические и динамические свойства мягких тканей Глава 5 • Механические и динамические свойства мягких тканей
составляет до 88 % площади поперечного сечения нерва. Он выдвигает предположение, что эта структура, по-видимому, является специальным защитным механизмом.
Эластичность нервных стволов
Третьим свойством, предохраняющим нерв от деформации, является его эластичность. Эластичность — это сопротивление материала растяжению, т. е. свойство, позволяющее ему восстановить свою первоначальную форму или размер. Основным компонентом, обусловливающим эластичность нервного ствола, является периневрий. Как свидетельствуют результаты исследований, диапазон эластичности периферических нервов составляет 6-20 %.
Факторы, ограничивающие эластичность и подвижность нервов. Периферические нервы проявляют такие качества, как прочность, эластичность и подвижность. Однако со временем эти свойства могут изменяться. Следующие факторы способны трансформировать механические характеристики нервных волокон:
адгезия и образование рубцов;
изменения соотношения коллагеновой и эластичной ткани нерва;
деформации;
травмы;
швы.
Влияют ли тренировочные занятия на периферические нервы? На сегодняшний день мы не знаем, как влияют различные виды традиционных режимов и методов растягивания на силу, эластичность и подвижность периферических нервов, поскольку никаких исследований в этом направлении не проводилось.
ФАКТОРЫ,
ВЛИЯЮЩИЕ НА МЕХАНИЧЕСКИЕ
СВОЙСТВА СОЕДИНИТЕЛЬНЫХ
ТКАНЕЙ И МЫШЦ
На поведение соединительных тканей (коллагеновых или эластичных) и мышц, находящихся под воздействием нагрузки, влияет целый ряд факторов, в том числе:
расположение или ориентация волокон;
воздействие различных структур взаимного переплетения коллагеновых молекул в каждой фибрилле;
наличие межфибриллярных субстанций;
количество волокон и фибрилл;
площадь поперечного сечения волокон;
соотношение количества коллагена и эластина;
химический состав тканей;
степень гидратации;
101
Наука о гибкости
степень расслабления сократительных компонентов;
температура ткани до и во время приложения силы;
температура ткани перед устранением действия силы;
количество прикладываемой силы (нагрузка);
продолжительность прикладывания силы (время);
тип прикладываемой силы (баллистическая или статическая).
НЕОБХОДИМОСТЬ ДАЛЬНЕЙШИХ ИССЛЕДОВАНИЙ
Процесс познания является бесконечным. В области механики мягких тканей имеется множество проблем, на которые следует обратить внимание. На две из них указал Г.К.Ли (1980):
Мягкая ткань в основном состоит из жидкости. Движение этой составной части — жидкости — играет важную роль в реакции ткани на деформирующие силы. Кроме того, реакции ткани на деформацию определяются механическими, электрическими и биохимическими характеристиками клеточных и молекулярных составляющих ткани. Предполагают, что эти факторы обусловливают зависимую от интенсивности сущность биомеханической реакции мягких тканей. Проведение исследований в этом направлении является весьма желательным.
В основном реакции мягких тканей в большей или меньшей степени контролируются и координируются нервной системой. Пассивные механические свойства эластичных волокон и биологических мембран изучены достаточно подробно многими учеными. Однако необходимо выяснить сущность взаимодействия мышц с пассивными компонентами мягких тканей.
РЕЗЮМЕ
Все ткани (соединительная, мышечная, нервная и сосудистая) подвергаются прогнозируемым изменениям при воздействии на них какой-либо силы. Под действием чрезмерной растягивающей силы происходит разрыв всех тканей. Степень повреждения определяется такими факторами, как величина силы, скорость ее приложения и продолжительность воздействия.
Исследования показывают, что оптимальное эластичное, или восстанавливающееся, удлинение имеет место при кратковременном растягивании с высоким усилием при нормальной или несколько пониженной температуре ткани. Пластическое, или остаточное, удлинение имеет место при продолжительном растягивании с невысоким усилием при повышенной температуре с последующим охлаждением ткани перед устранением напряжения. Кроме того, минимальное структурное ослабление связано с растягиванием с невысоким усилием в сочетании с повышенной температурой, тогда как максимальное ассоциируется с высоким усилием и более низкой температурой.
102
 Глава 5 • Механические и динамические свойства мягких тканей Глава 5 • Механические и динамические свойства мягких тканей
Таким образом, идеальная программа растягивания, обеспечивающая оптимальное увеличение длины ткани без повреждений, предус-матривает повышение температуры перед растягиванием(путем выпол-
нения физических упражнений или терапевтических процедур, вызывающих повышение внутренней температуры тела), использование усилий небольшой интенсивности, поддержание растягивающего усилия в течение более продолжительного периода времени и охлаждение ткани до нормальной температуры перед устранением растягивающей силы.
Кровеносные сосуды и периферические нервы также обладают способностью растягиваться. Растягивание кровеносных сосудов приводит к снижению кровотока. Чрезмерное растяжение нервов вызывает нарушение функции и постепенно ведет к механическому повреждению.
|
 Скачать 10.31 Mb.
Скачать 10.31 Mb.