Майкл Дж[1]. Наука о гибкости. Литература ббк 75. 0 А52
 Скачать 10.31 Mb. Скачать 10.31 Mb.
|

|
ГЛАВА 6 НЕЙРОФИЗИОЛОГИЯ ГИБКОСТИ: НЕВРАЛЬНАЯ АНАТОМИЯ И ФИЗИОЛОГИЯ Нервная система представляет собой одну из основных коммуникационных систем организма. Поэтому она играет важную роль в определении возможного количества движения. Нервная система состоит из центральной нервной системы — ЦНС (головной и спинной мозг) и периферической нервной системы (черепные и периферические нервы). Упрощенную модель функциональных элементов нервной системы иллюстрирует рис. 6.1. Центральная — Головной мозг  Нервная система Нервная система— Спинной мозг Спинномозговые нервы — Симпатическая Автономная — — Парасимпатическая Рис. 6.1. Структура нервной системы СТРУКТУРНАЯ ОСНОВА: КЛЕТОЧНАЯ НЕЙРОАНАТОМИЯ Структурной и функциональной единицей нервной системы является нейрон (нервная клетка). Чтобы понять сущность нервной системы, необходимо знать, как функционирует нейрон. Нейроны, подобно всем остальным клеткам организма, имеют структурное строение, соответствующее их основной функции — приему и передаче электрических импульсов. Чтобы нервная система функционировала, необходимы три условия. Во-первых, должны быть средства выявления стимулов или изменений в окружающей среде (например растягивание). Эту функцию выполняют структуры, называемые рецепторами. Во-вторых, после приема стимула его необходимо передать, что обеспечивают проводники нервной системы — нейроны. И наконец, реагирующие органы должны пере- 104  Г л а в а б. Нейрофизиология гибкости: невралъная анатомия и физиология Г л а в а б. Нейрофизиология гибкости: невралъная анатомия и физиология  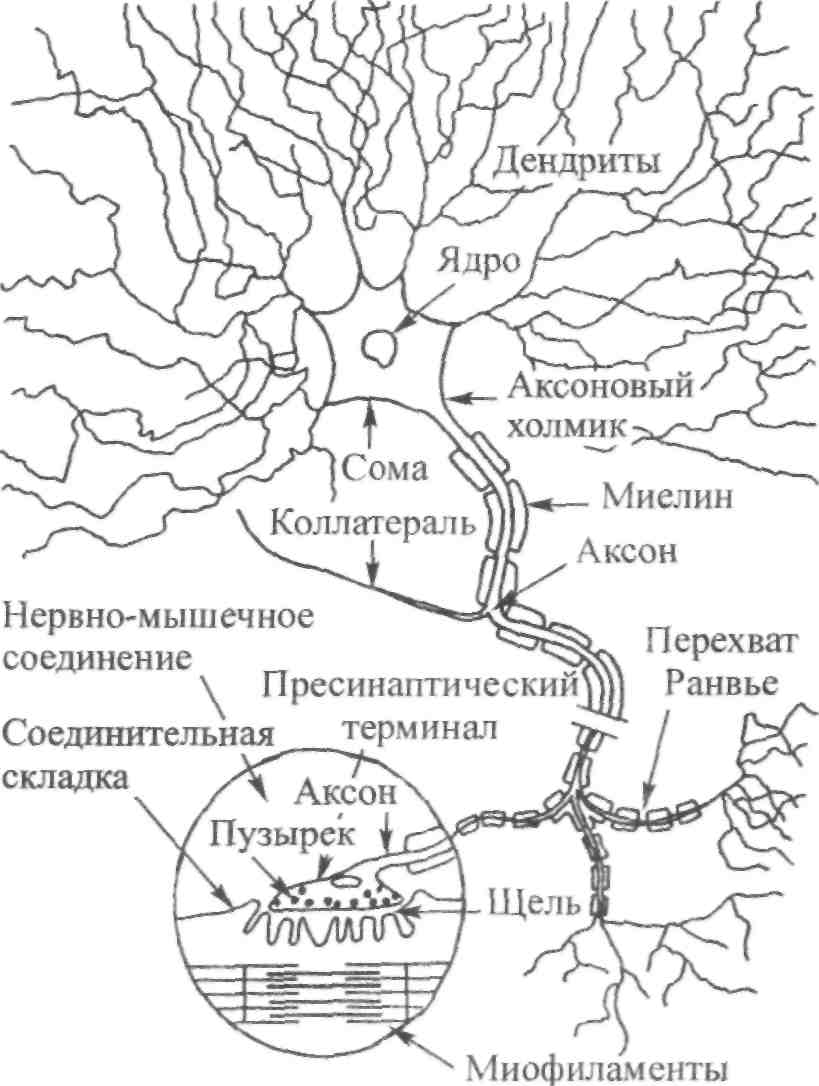 дать соответствующие реакции стимулу. За это ответственны эффекторы (такие, как мышцы и железы). Нейрон. В структурном строении нейрона имеется четыре обособленных морфологических участка: клеточное тело (сома), один или больше дендритов, отдельный аксон и пресинаптический терминал (рис. 6.2). Клеточное тело, или сома, содержит ядро и протоплазму. Протоплазма представляет собой всю живую субстанцию в клетке, окружающую ядро. Ядро отвечает за контроль над всеми процессами, происходящими в клетке. Рис. 6.2. Четыре морфологических участка типичного нейрона: дендриты, сома, аксон и пресинаптические терминалы (Enoka, 1988) Дендрит представляет собой тип нервного волокна, простирающегося из клеточного тела. Дендриты имеют обширные ответвления. Слово дендрит происходит от греческого dendron — дерево. Его функция заключается в приеме и передаче импульсов к клеточному телу, что представляет собой так называемый афферентный процесс. Длинная часть нейрона, идущая от клеточного тела, называется аксоном. Большинство аксонов отводят импульсы от сомы. В то же время они могут передавать импульсы в другом направлении. Аксоны обычно имеют ответвления, которые называются коллатералями. Конечной точкой аксона, включенного в синапс, является пресинаптический терминал. Эта структура является местом соприкасания, обеспечивающим нейрон возможностью передачи сигнала к клетке-мишени. Большие аксоны покрыты жировой изолирующей оболочкой миелина. В периферических нервах миелин производят шванновские клетки. Интервалы между соседними шваннов-скими клетками называются перехватами Ранвье. В этих участках заряженные ионы могут пересекать мембрану аксона, что позволяет регенерировать электрические сигналы. Аксоны образуют так называемую эфферентную систему. Нервы. Пучки нейроновых волокон называются нервами. Они соединяются друг с другом оболочками соединительной ткани (эпиневрий, периневрий и эндоневрий). Почти все нервы являются смешанными (то есть содержат как афферентные, так и эфферентные волокна). Афферентные волокна проводят импульсы к ЦНС. В зависимости от размера и 105    Наука о гибкости Наука о гибкостипроводимости их делят на группы I-IV. Ни одна из функций организма человека не может осуществиться без какого-либо контроля со стороны ЦНС. Даже простая реакция на стимул включает, по крайней мере, спинной мозг. Эфферентные волокна, наоборот, проводят импульсы от ЦНС к эффекторам. ЭЛЕКТРИЧЕСКИЙ ПОТЕНЦИАЛ НЕРВА Функция нерва заключается в передаче сигналов в виде нервных импульсов из одной части тела в другую. Эта передача импульсов представляет собой электрохимический процесс. Мы рассмотрим два вида электрических потенциалов. Потенциал покоя. Нейроны являются поляризованными. Иначе говоря, они имеют неодинаковое количество различных видов ионов внутри мембраны и за ее пределами. Ион представляет собой атом, который приобрел или утратил один или несколько своих электронов и, следовательно, несет положительный или отрицательный заряд. Ионы движутся через жидкость в ответ на электрическое поле или градиент концентрации (разница в концентрации ионов между двумя жидкими участками). Когда нейрон находится в покое, на внешней стороне мембраны находится больше ионов натрия (Na+), чем на внутренней, в то же время ионов калия (К+) содержится больше на внутренней, чем на внешней стороне мембраны; ионы натрия при этом могут транспортироваться наружу через мембрану благодаря транспортному механизму, который называется натрий-калиевым насосом. Чтобы поддержать в таком виде потенциал покоя мембраны, требуется непрерывный расход энергии. Ионы калия могут проходить через мембрану благодаря пассивному транспорту. Таким образом, некоторые из них выйдут из клетки. Их возвращение в клетку обеспечивает натрий-калиевый насос в обмен на Na+, которые выкачиваются наружу. Разница в концентрации ионов по обеим сторонам мембраны нейрона является основой для установления потенциала мембраны. Внутренняя часть клеток, находящихся в покое, является отрицательно заряженной относительно интерстициальной жидкости. Следовательно, нейрон можно рассматривать как своеобразную батарейку с отрицательным терминалом внутри. Разность потенциалов мембраны в покое составляет приблизительно 0,1 В. Производство потенциалов действия. При стимулировании аксона изменяется его потенциал. Стимул изменяет степень проницаемости нев-ральной мембраны. В результате стимулирования степень проницаемости мембраны для ионов натрия (Na+) повышается. Последние быстро диффундируют в нервное волокно. Это направленное вовнутрь движение ионов натрия изменяет остаточный потенциал покоя мембраны, т.е. внешняя часть нервного волокна становится отрицательной относительно внутренней. Следовательно, меняется и полярность нерва. Происходит деполя- 106 Г л а в а 6. Нейрофизиология гибкости: невралъная анатомия и физиология   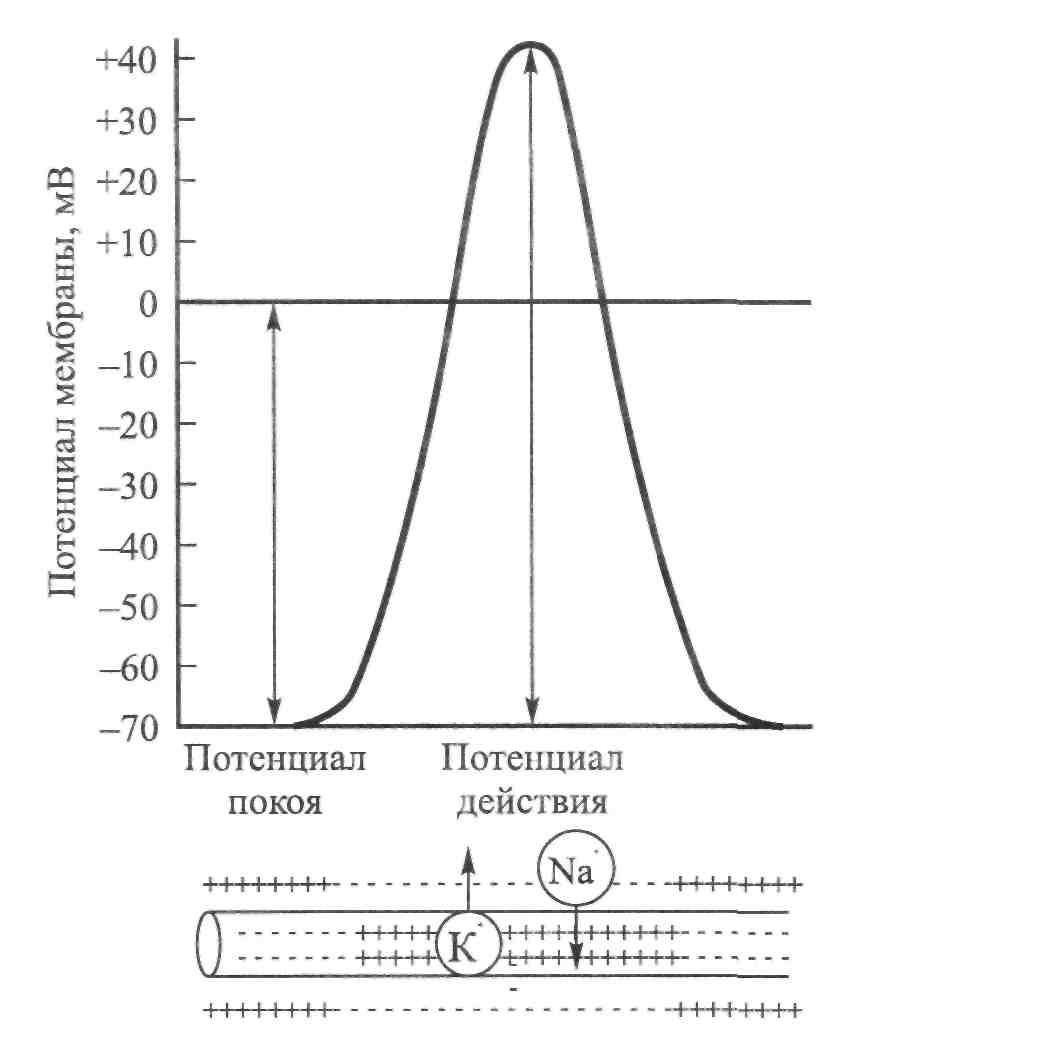 ризация нервного волокна; когда она достигает критического уровня напряжения, образуется потенциал действия. Таким образом, потенциал действия и деполяризация взаимосвязаны, но не являются синонимами. Деполяризация представляет собой поток ионов, инициирующий потенциал действия. При пике деполяризации поступление ионов натрия в клетку замедляется и мембрана становится непроницаемой для натрия. Рис. 6.3. Потенциал действия. Движение ионов через мембрану нервного волокна (внизу) совпадает с изменениями электрического потенциала (вверху). Когда ионы натрия быстро перемещаются в аксон, потенциал мембраны становится положительным. После этого происходит движение ионов калия во внешнюю часть волокна (Alter, 1988) Параллельно происходит изменение степени проницаемости другой клеточной мембраны. После короткого интервала значительно повышается степень проницаемости мембраны для калия (К+). Калий способен легко диффундировать наружу ввиду его высокой концентрации внутри мембраны. В результате ионы калия начинают покидать клетку, унося с собой положительные заряды. Отток ионов калия восстанавливает исходный отрицательный заряд внутри мембраны. Следовательно, разница напряжения мембраны возвращается к своей норме. Этот процесс, называемый реполяризацией, вызывает прекращение потенциала действия (рис. 6.3). Чтобы мембрана восстановила свой исходный потенциал покоя, ионы натрия должны вернуться на внешнюю, а ионы калия — на внутреннюю часть мембраны. Натрий-калиевый насос является активным транспортным механизмом, возвращающим ионы на исходные позиции. Так восстанавливается исходный потенциал мембраны. Закон «все или ничего». Если стимул (например, растяжение) является достаточно мощным, чтобы инициировать нервный импульс в аксоне, он вызывает производство полноценного потенциала действия. Это явление известно как закон «все или ничего», его можно сравнить с процессом стрельбы. Как только сила тяги, направленная на курок, оказывается достаточной, чтобы привести в действие ударный механизм, звучит выстрел. Это критическое значение называется порогом. Увеличение силы тяги не меняет скорость полета пули, поскольку не сила тяги вызывает реакцию (Sage, 1971). Иными словами, либо стимул является достаточно мощным, чтобы стимулировать волокно, либо нет. Усиление стимуляции не приводит к увеличению потенциалов действия. 107 Наука о гибкости Передача информации в ЦНС об интенсивности растягивания. Каким же образом нерв осуществляет дифференциацию различной интенсивности растягивания? Нерв имеет две возможности передачи информации о растягивании с разной интенсивностью. Во-первых, он может одновременно передавать ощущения растягивания различному числу нервных волокон. Поэтому более высокая интенсивность ощущений растягивания может привести в действие большее число чувствительных нервных волокон. Этот процесс называется рекрутированием. Следовательно, интенсивность растягивания можно увеличить, увеличив рекрутирование рецепторов и чувствительных нейронов. Например, слабый стимул растягивания активирует только рецепторы растяжения, имеющие наименьший порог. Однако по мере увеличения интенсивности активируется все большее число менее возбудимых тензорецепторов, что увеличивает число чувствительных единиц. Кроме того, нерв может передавать различное число импульсов за единицу времени в одно и то же волокно. Вследствие этого изменения интенсивности растягивания могут отражаться различной интенсивностью разрядки нервных импульсов в отдельных волокнах: чем выше интенсивность растягивающего стимула, тем выше частота растягивающего импульса. Этот процесс называется интенсивностью кодирования частоты. Итак, чем мощнее растягивающий стимул, тем больше число активных сенсорных нейронов и тем выше частота импульсов в каждом из них. Таким образом, бомбардировка мотонейронов в спинном мозге и в кортикальных центрах головного мозга становится более интенсивной, что ведет к повышенной рефлекторной двигательной активности и усилению ощущений. Адаптация чувствительных нервных окончаний. При воздействии стимула на чувствительное нервное окончание с постоянной силой рецептор, как правило, первоначально реагирует высокой интенсивностью разрядки. Генерируемый потенциал вначале пропорционален интенсивности стимула. Однако во время устойчивого стимулирования он постепенно снижается, вследствие чего интенсивность разрядки уменьшается. Это явление называется адаптацией. Если же воздействие стимула прерывается даже на очень короткий момент, имеет место «взрыв» импульсной активности и процесс повторяется. Тензорецепторы делятся на быстро- и медленноадаптирующиеся. Быстроадаптирующиеся единицы проявляют более быстро снижающуюся интенсивность разрядки при сохранении растягивания (так как в этих единицах потенциал снижается значительно быстрее). С другой стороны, медленноадаптирующиеся единицы проявляют непрерывную интенсивность разрядки при поддерживаемом стимулировании. Они проявляют продолжительно сохраняющиеся потенциалы. Понятия фазные и тонические используются для обозначения быстро- и медленноадаптирующихся рецепторов соответственно (рис. 6.4). Исследования показывают, что быстроадаптирующиеся рецепторы, такие, как тельца Пачини (рецепторы кожи и суставов), ведут себя как ме- 108 Г л а в а 6. Нейрофизиология гибкости: невральная анатомия и физиология  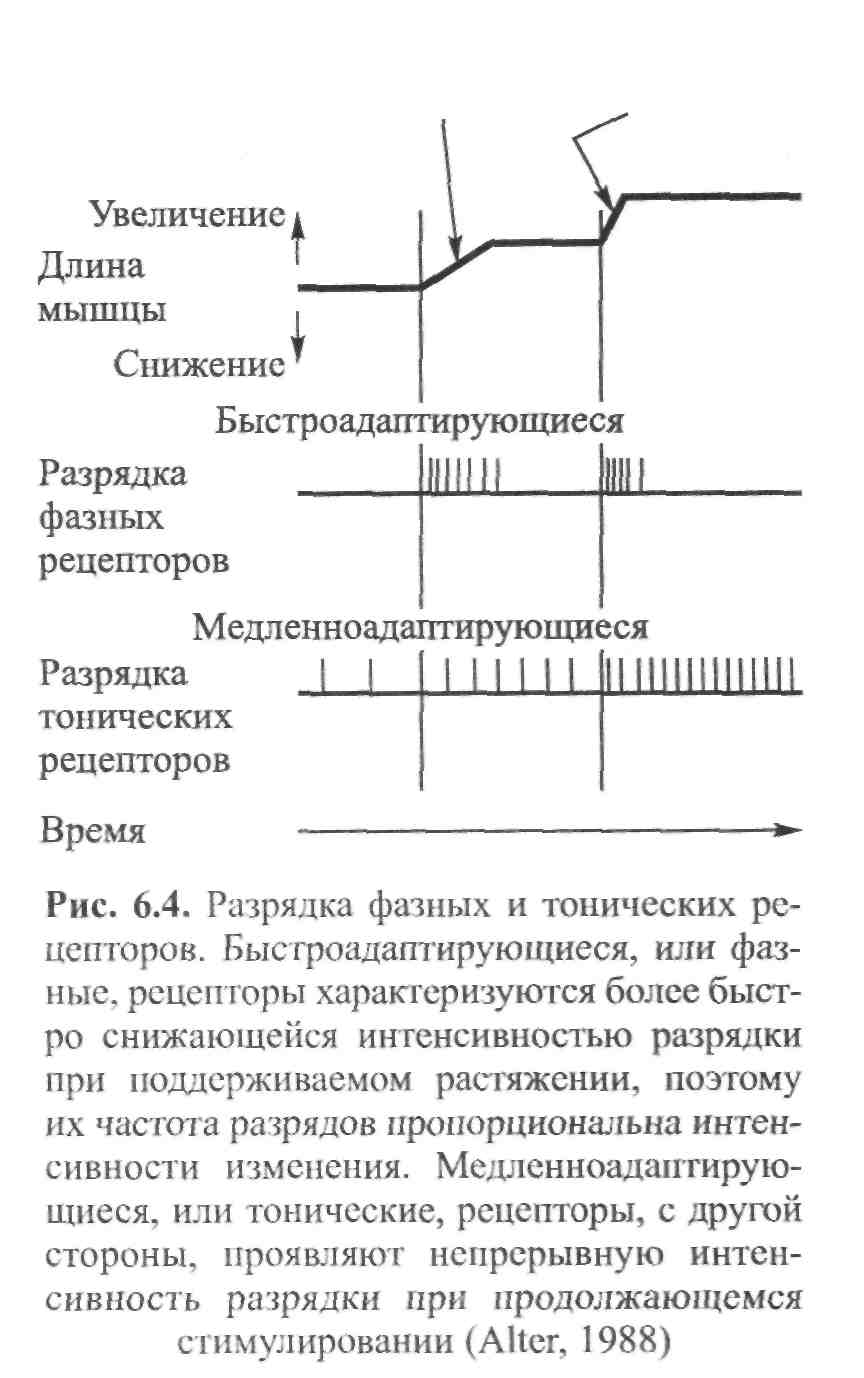 ханические высокочастотные Медленное Быстрое фильтры. Предполагают, что эти растяжение растяжение фильтры состоят из вязких и упругих компонентов. Если усилие воздействует на вязкие компоненты в течение достаточно короткого промежутка времени, оно передается непосредственно в сердцевину рецептора, где происходит нагрузка нервного окончания. Этот динамический компонент быстро снижается, независимо от продолжительности действия стимула. Однако сила, накопленная в растянутых эластичных компонентах, передается затем с заметным снижением в сердцевину рецептора. В результате этого отсутствует стабильный потенциал. С другой стороны, медленно-адаптирующиеся рецепторы, такие, как мышечные волокна (являющиеся рецепторами длины мышцы), не имеют механических высокочастотных фильтров у нервных окончаний. Следовательно, в результате стабильного растягивания образуются стабильные потенциалы. Степень адаптации, которая имеет место, обусловлена, по-видимому, присутствием вязких компонентов в самом веретене. ЧУВСТВИТЕЛЬНЫЕ РЕЦЕПТОРЫ, СВЯЗАННЫЕ С РАСТЯГИВАНИЕМ Три вида рецепторов непосредственно связаны с растягиванием и поддержанием оптимального диапазона движения. Это нервно-мышечные веретена, нервно-сухожильные веретена и суставные механорецепто-ры. Далее мы рассмотрим их структуру, функции и взаимосвязь с растягиванием. Нервно-мышечные веретена. Основными тензорецепторами мышцы являются нервно-мышечные веретена. Это наиболее подробно изученные проприорецепторы — рецепторы, расположенные в мышцах, сухожилиях и преддверии уха. Их рефлексы связаны с локомоцией или положением тела. В разном количестве их можно обнаружить в большинстве скелетных мышц тела. Особенно много нервно-мышечных веретен содержится в небольших мышцах кисти и глаза. Поскольку мышечные 109  Наука о гибкости Наука о гибкостиволокна заключены в веретенообразную капсулу (оболочку соединительной ткани, окутывающую рецептор), их называют интрафузальными волокнами. Экстрафузальные волокна являются обычными сократительными единицами мышцы. Веретена прикрепляются к обоим концам экстра-фузальных волокон и, таким образом, располагаются параллельно им. Следовательно, при растягивании мышцы веретено также подвергается растягиванию. Веретено иптрафузалъныхмышечных волокон Существует два основных типа интрафузальных волокон: волокна ядерного мешочка и волокно ядерной цепочки. Первые содержат в большом количестве саркоплазму и клеточные ядра в расширенной, напоминающей мешок структуре. Эта несократительная структура находится в центральном, или экваториальном, участке интрафузального волокна. Отсюда и название — интрафузальное волокно ядерного мешочка. На дистальных, или полярных, концах волокна ядерного мешочка находятся исчерченные сократительные филаменты, которые возле веретен прикрепляются к эк-страфузальным волокнам. Не так давно было установлено, что существует два подвида волокон ядерного мешочка. Их назвали мешочек1 и мешочек, (Gladden, 1986; рис. 6.5). 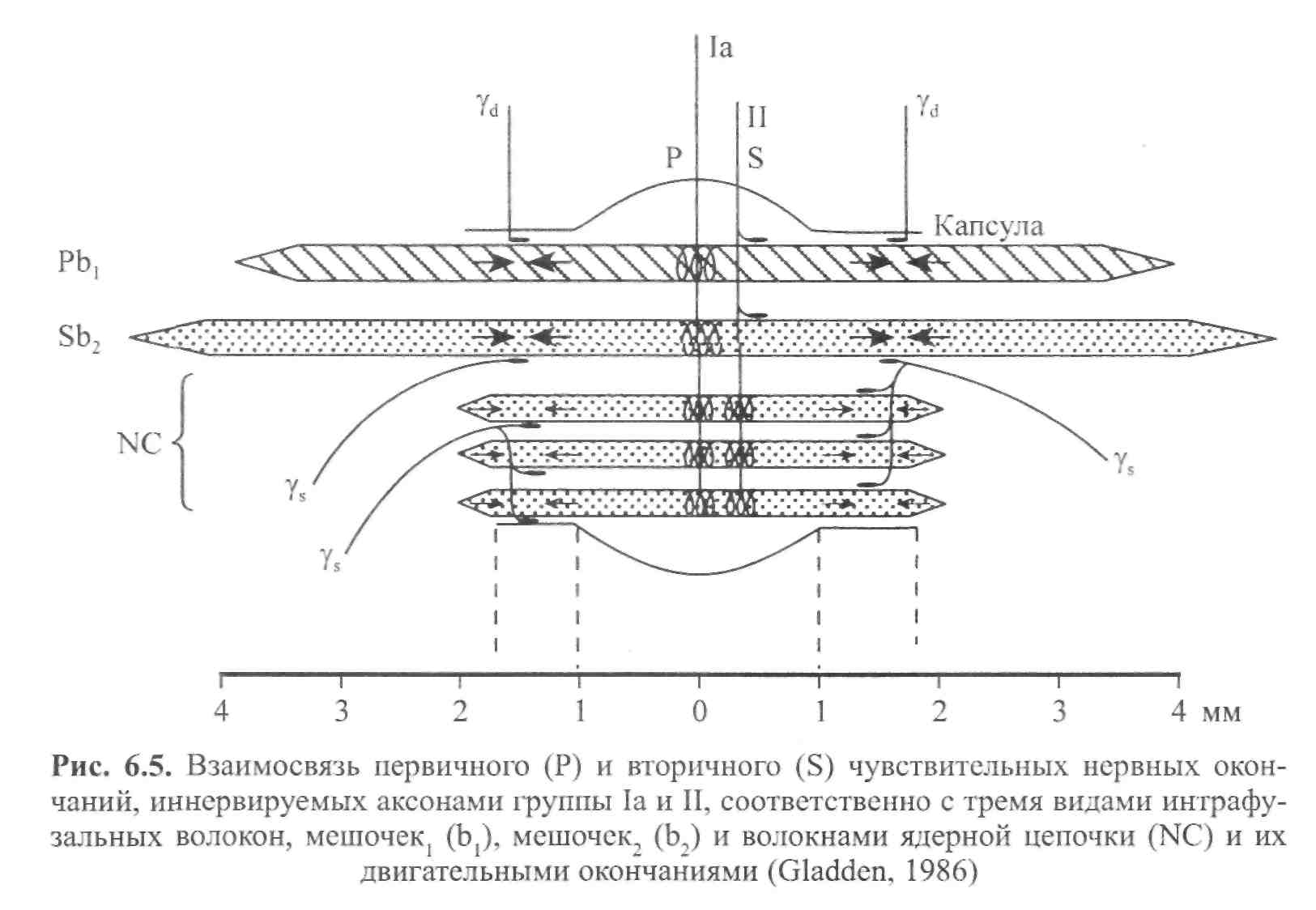 110 Второй тип интрафузального мышечного волокна — волокно ядерной цепочки — более тонкое и короткое, чем волокно ядерного мешочка. Оно содержит всего один ряд ядер, расположенных в виде цепочки по несократительному экваториальному участку. Подобно волокнам ядерного мешоч-     Г л а в а 6 . Нейрофизиология гибкости: невралъная анатомия и физиология Г л а в а 6 . Нейрофизиология гибкости: невралъная анатомия и физиологияка, полярные концы волокна ядерной цепочки также состоят из исчерченных сократительных филаментов. Их концы нередко соединяются с волокнами ядерного мешочка, которые, в свою очередь, прикрепляются к эндо-мизию экстрафузальных волокон. Чувствительные нервы веретен Существует два вида чувствительных нервных (афферентных) окончаний в каждом веретене: первичные и вторичные. Первичные окончания заканчиваются, спиралевидно окружая центральный участок волокна ядерного мешочка и выпуская боковое ответвление к волокну ядерной цепочки. Афферентные аксоны первичных нервных окончаний относятся к волокнам большой группы I. Чтобы отличить эти чувствительные нервные окончания от других, относящихся к этой же группе, большие афференты веретена называют афферентами группы 1а. Первичные нервные окончания имеют очень низкий порог растяжения и поэтому легко возбуждаются. Их реакции могут быть как фазными (динамическими), так и тоническими. Фазная реакция определяет интенсивность или скорость растягивания путем изменения частоты нейронных импульсов во время растягивания. Частота разрядки быстро увеличивается при начальном растягивании. Затем, по достижении новой длины, она снижается до постоянного уровня, соответствующего новой тонической длине. Следовательно, тоническая реакция определяет длину мышцы. Иными словами, первичные нервные окончания определяют длину и скорость растягивания. Вторичные нервные окончания образуют разветвленные, напоминающие цветок окончания. Они содержатся в основном в околоэкваториальном сегменте (вблизи экватора) волокон ядерной цепочки. Аксоны вторичных нервных окончаний относятся к афферентным волокнам группы П. В отличие от первичных нервных окончаний, вторичные нервные окончания определяют только тоническую длину мышцы. Это мотонейроны вызывают соккращение мышечных филаментов в полярных концах интрафузальных мышечных волокон. При сокращении этих концов в центральном экваториальном участке наблюдается пассивное растягивание (там находятся чувствительные нейронные рецепторы). Таким образом, активация гамма-мотонейронов ЦНС может увеличивать количество растяжения, испытываемого чувствительными нервными окончаниями (Banker, 1980). Двигательные нервы веретена Двигательные нервы, иннервирующие каждое интрафузальное мышечное волокно в его полярных участках, называются у-эфферентны-ми волокнами, образующими фузимоторную систему. Различают два вида у-аксонов в зависимости от их воздействия на первичные и вторичные чувствительные нервные окончания. Стимулирование так называемого статического у-аксона (ys) может усиливать чувствительность первично- 111 Наука о гибкости го нервного окончания к длине, не влияя (или незначительно влияя) на восприятие (чувствительность) скорости. С другой стороны, динамические у-аксоны (yd) могут существенно усиливать восприятие скорости первичного нервного окончания, практически не влияя на чувствительность к длине. Функция у-системы заключается в контроле чувствительности веретена к растягиванию. Этот процесс сенситизации веретена при помощи у-эф-ферентов известен как у-смещение (Norback и Demarest, 1981). Активация у-мотонейронов приводит к сокращению или укорачиванию интрафузаль-ных мышечных волокон в их полярных участках. При сокращении интра-фузальных волокон экваториальный участок растягивается. Эта центральная деформация приводит к перетягиванию колец в аннуло спиральную часть первичных окончаний и увеличению интенсивности разрядки аффе-рентов групп 1а и П. Вторая функция у-мотонейронов — поддержание чувствительности веретена во время сокращений мышцы. Когда мышцы укорачиваются, веретено также пассивно укорачивается. Это пассивное сближение двух концов веретена снимает напряжение как с его первичных окончаний (разгрузка веретена), так и со вторичных; такая разгрузка лишает головной мозг информации из веретена относительно изменений мышечной длины. Чтобы предотвратить разгрузку веретена и отрегулировать его чувствительность, у-мотонейроны активируют. Процесс возбуждения нервно-мышечного веретена Процесс возбуждения нервно-мышечного веретена можно охарактеризовать следующим образом. Во-первых, к нервно-мышечному веретену прикладывают минимальный растягивающий стимул. Во-вторых, в чувствительном нейронном окончании происходит изменение степени проницаемости, что приводит к производству генерирующего тока (передача заряда через мембрану нервного терминала). Последний, в свою очередь, вызывает деполяризацию, которая называется генератор потенциала. При несколько большей степени растягивания его амплитуда в окончаниях нервно-мышечного веретена увеличивается. Когда деполяризация достигает порога, возникает проведенный потенциал действия. При дальнейшем увеличении степени растягивания может наблюдаться серия проводимых нервных импульсов. Этапы возбуждения нервно-мышечного веретена приведены ниже:
|