Практикум по систематике. Министерство образования и науки российской федерации
 Скачать 23.75 Mb. Скачать 23.75 Mb.
|

|
Вопросы для самоконтроля
III. ЦАРСТВО РАСТЕНИЯ – PLANTAE, PHYTOBIONTA, или VEGETABILIA Растения отличаются от водорослей многими признаками, они являются многоклеточными автотрофными организмами, приспособленными к обитанию в воздушно-наземной среде. В процессе длительной эволюции у растений выработалась общая структура, выражающаяся в их морфологической расчлененности на листостебельный побег и корневую систему и в сложном анатомическом строении их органов. У растений возникают специальные органы поглощения минеральных растворов из субстрата корни и корневые волоски у спорофита и ризоиды у гаметофита. В хлоренхиме листьев осуществляется фотосинтез. Проводящие ткани связывают два важнейших концевых аппарата – корневые волоски и листья, а механические ткани обеспечивают устойчивое положение растения в почве и в воздухе. Стебель своим ветвлением и листорасположением обеспечивает наилучшее размещение листьев в пространстве, чем достигается наиболее полное использование световой энергии, а ветвлением корня – эффект размещения огромной всасывающей поверхности корневых волосков в почве. Для растений характерно закономерное чередование двух поколений – гаметофита (полового), несущего половые органы с гаметами и спорофита (бесполого), несущего спорангии со спорами. При этом зигота превращается в многоклеточный зародыш, из которого развивается спорофит. Спорофит является преобладающим «взрослым» растением. Спорофит у подавляющего большинства растений расчленен на побеги (стебель и листья) и корни. Основу клеточной стенки растений составляют полисахариды: целлюлоза, гемицеллюлозы и пектиновые вещества. Растения имеют пигменты – хлорофиллы а и b и каротиноиды. Основной запасной продукт - крахмал, который образуется на лейкопластах. Размножение вегетативное, бесполое (спорами) и половое В царстве растений выделяются несколько отделов, которые объединены в три группы: бессосудистые споровые – отдел моховидные (Bryophyta), сосудистые споровые – плауновидные (Lycopodiophyta), хвощевидные (Equiseto-phyta), папоротниковидные (Polypodiophyta), семенные - голосеменные (Gymnospermae, или Pinophyta) и покрытосеменные (Angiospermae, или Magnoliophyta). Кроме того, к царству растений относятся вымершие отделы – риниевидные (Rhyniophyta), зостерофилловидные (Zosterophyllophyta) и небольшой своеобразный отдел псилотовидные (Psilotophyta), которые мы не рассматриваем. Безсосудистые споровые растения III.1. Отдел Моховидные - Bryophyta Оборудование и материалы: микроскоп, лупа, предметные и покровные стекла, иглы препаровальные, живые и гербарий мхов: кукушкина льна и других видов зеленых мхов; сфагновые мхи, постоянные микропрепараты поперечных срезов листа и стебля, продольного среза коробочки спорогония кукушкина льна и сфагнума. Моховидные – бессосудистые растения, у них нет развитой проводящей системы (трахеид, сосудов, ситовидных трубок). В царстве растений моховидные занимают особое место. Во-первых, они типичные обитатели суши с приспособленными к воздушной среде гаметангиями и спорангиями и с четким чередованием поколений. Во-вторых, в жизненном цикле у моховидных по сравнению с остальными растениями все наоборот – преобладает не спорофит, а гаметофит. И стебель, и листья характерны не для спорофитов, а для гаметофитов. Гаметофит представляет собой растение в виде побега, расчлененного на стебель и листья, или же листовидного таллома. Спорофит назывется спорогоном и представляет собой коробочку с ножкой, расширенной в нижней части в стопу-гаусторию, с помощью которой он, присасывется к гаметофиту, живет за его счет, так как от него получает воду и необходимую пищу. В период созревания он может иметь зеленую окраску и фотосинтезировать. Моховидные – небольшие многолетние (за редким исключением) вечнозеленые растения, у которых отсутствуют корни. К субстрату они прикрепляются ризодами, представляющими собой сильно вытянутые в длину одиночные клетки у примитивных представителей (печеночников) и нити из одного ряда клеток, разделенных косыми перегородками, у более совершенных. Поглощение и испарение у моховидных осуществляется всей поверхностью гаметофита. У некоторых мхов в стебле есть проводящий пучок из узких и длинных клеток – гидроидов и лептоидов, выполняющих функцию сосудов и ситовидных трубок. Мхи могут быть одно-, и двудомными. На гаметофитах образуются антеридии и архегонии обычного для растений строения. Процесс оплодотворения возможно только при наличии воды. При отсутствии воды половой процесс не происходит. По этой причине в жизни мхов большую роль играет вегетативное размножение. Большинство мхов вегетативно размножаются с помощью специальных органов – выводковых почек, листьев, частей листьев, веточек, которые образуются на гаметофите. Эти органы могут длительное время находиться в состоянии покоя и переноситься на большие расстояния водой и ветром. Моховидные способны оживать после полного высыхания, любая часть может дать начало новому растению. Как этап вегетативного размножения можно рассматривать прорастание многих почек на протонеме – нитевидной или пластинчатой стадии гаметофита, развивающегося из споры. В результате из одной проросшей споры образуется много гаметофитов. Расселение мхов происходит при помощи спор. Споры образуются в спорангиях на бесполом поколении мхов – спорогонии. Половой процесс у мхов происходит в половых органах. Из зиготы вырастает ножка и коробочка со спорами – спорогоний, в котором образуются споры. Форма коробочки, способы ее вскрывания, механизм разбрасывания спор во многом определяют систематику листостебельных мхов. Моховидные широко распространены в природе, они произрастают в различных местообитаниях, но везде, как правило, предпочитают наиболее увлажненные условия. Они играю существенную роль в растительном покрове, особенно тундры, болот и лесов. Отдел моховидные подразделяют на 3 класса: Анто-церотовые (Anthocerotopsida), печеночники (Hepatico-psida) и листостебельные мхи (Bryopsida). Будут рассмотрены только два последних. III.1.1. Класс Печеночники, или Печеночные мхи – Marchantiopsida, или Hepaticopsida Печеночники имеют пластинчатые или слоевищные гаметофиты, характеризующиеся дорзивентральным строением, т.е. различающиеся верхней и нижней сторонами. Верхняя сторона отличается от нижней стороны, наличием ризоидов. Ризоиды одноклеточные. В клетках слоевища имеются особые образования – масляные тельца. Печеночники разделяются на слоевищные – подкласс маршанциевые и листостебельные – подкласс юнгерманиевые, причем последние преобладают по числу видов и распространены преимущественно в тропиках и субтропиках. В средней полосе чаще всего встречаются слоевищные печеночники, одним из представителей которых является маршанция многообразная (Marchantiapolymorpha) (рис. 15), обитающая в сырых местах, по краям болот на торфяной почве, по стенкам канав, у родников и т.д. Несмотря на широкое распространение маршанции в природе, они не играют доминирующей роли в растительном покрове. Задание
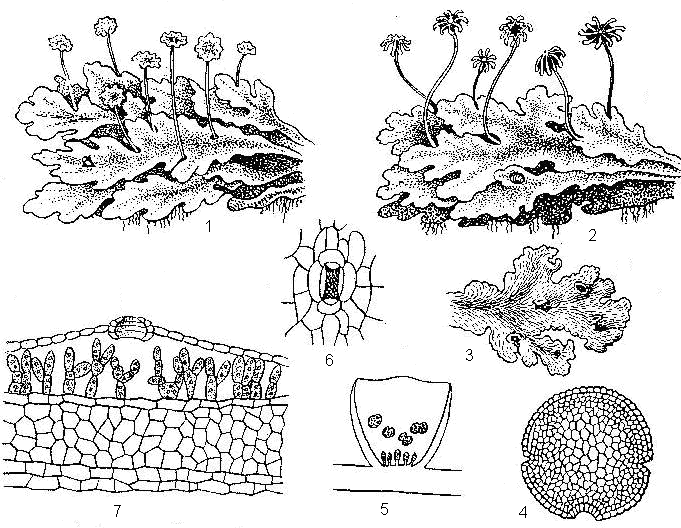 Рис. 18. Маршанция: 1 – слоевище с антеридиальными подставками; 2 - слоевище с архегониальными подставками; 3 – слоевище с выводковыми корзиночками; 4 – выводковая почка; 5 – разрез корзиночки с выводлковыми почками; 6 – устьице (вид сверху); 7 – анатомическое строение слоевища; верхний эпидермис с устьицем, ассимиляционная камера Маршанция представляет собой стелющуюся, дихотомически ветвящегося лентовидного таллома длиной 5-20 см с углублением на вершине. В углублении находится точка роста, за счет деятельности ее клеток происходит нарастание слоевища по обе стороны от выемки и возникают новые ветви. В средней части слоевища проходит дихотомически ветвящаяся жилка. Нижняя сторона таллома сплошь покрыт буроватыми ризоидами. Старые части слоевища постепенно отмирают, когда отмирание доходит до места ветвления, каждая ветвь продолжает расти дальше как самостоятельная особь. Таким образом, происходит примитивное вегетативное размножение маршанции. При рассмотрении верхней поверхности таллома в лупу отмечают мелкий сетчатый рисунок разграничивающий всю его поверхность на мелкие, слегка выпуклые ромбические, или пяти-, шестиугольные участки. В центре каждого участка выделяется пятно с точкой в центре. Пятна – это устьица, темные точки в них – устьичные отверстия. На нижней стороне слоевища находятся многочисленные ризоиды двух типов – простые и язычковые. Язычковые ризоиды располагаются рядами вдоль таллома, длиной до 2 см, они мертвые и поглощают влагу по принципу капиллярности. Переплетаясь друг с другом, язычковые ризоиды образуют шнуровидные тяжи, которые как фитиль, поглощают воду. У основания таллома они углубляются в почву. Рядом с язычковыми ризоидами располагаются бесцветные или фиолетовые однослойные мелкие листовидные чешуйки, называемые амфигастриями. Они защищают точку роста и прижимают ризоиды к таллому. Простые ризоиды, располагающиеся по обеим сторонам от жилок живые, растут вниз и углубляются в почву, всасывают воду и прижимают точку роста ближе к почве. На поперечном срезе микропрепарата слоевища рассматривают анатомическое строение маршанции. При малом увеличении видно, что верхняя часть среза зеленая, нижняя, более мощная, почти бесцветная. Сверху таллом покрыт эпидермой, под которой располагаются воздушные камеры ромбовидной формы. Камеры разделяются однослойными бесцветными стенками. Клетки эпидермы и камер содержат мало хлорофилловых зерен. Внутри камер находятся однорядные нити, слабо связанные друг с другом клетки - ассимиляторы, богатые хлорофилловыми зернами. Полость камеры сообщается с окружающей средой через незакрывающиеся устичьное отверстие. Таким образом, каждой ромбовидной фигурке на верхней стороне слоевища соответствует воздушная камера с устьицем в центре. Маршанция – двудомное растение, архегонии и архегонии развиваются на особых подставках, формируются на разных особях и напоминают миниатюрные грибочки со шляпками, рассеченными на лопасти. Антеридии имеют форму 8-лучевого диска, с углублениями. При созревании сперматозоиды в тонкой пленке воды благодаря хемотаксису движутся по направлению к женскому таллому. По образующемуся капиллярному пространству вода вместе со сперматозоидами снизу поднимается к архегониям. После оплодотворения яйцеклетка дает зиготу. Дальше из зиготы образуется в спорогон на короткой ножке. Внутри в клетках материнской ткани происходит редукционное деление и формируются одноклеточные гаплоидные споры с элатерами. Попадая на влажную почву, спора прорастает и дает небольшой предросток (протонему), разрастающийся путем деления клеток в пластинку – зачаток молодого слоевища. На верхней стороне таллома маршанции находят специальные органы вегетативного размножения – выводковые почки. Они развиваются в особых вместилищах - корзиночках. Рассматривая в бинокуляр выводковую корзиночку, обращают внимание на ее форму и на находящиеся в ней выводковые почки, вырастающие из эпидермальных клеток дна корзиночки в виде зеленых пластинок. Затем извлекают препаровальной иглой из корзиночки несколько выводковых почек и рассматривать их в микроскоп. Выводковые почки располагаются на ножках. Рядом с почками имеются слизевые клетки, вырабатывающие слизь. Она выталкивает выводковые почки наружу. Зарисовывают слоевища с антеридиальными и с архегониальными подставками и с выводковыми почками, и анатомическое строение слоевища. Ш.1.2. Класс Листостебельные мхи – Bryopsida, или Musci Листостебельные мхи характеризуются наличием зеленых побегов радиальной симметрии, расчлененного на стебель и листья, корень отсутствует. У зеленых и андреевых мхов листья многослойные, у сфагнума они однослойные и дифференцированы на два типа специализированных клеток. Ризоиды многоклеточные, часто ветвящиеся. Архегонии и антеридии собраны группами на верхушках главных побегов или на коротких боковых ветвях. Иногда они окружены специализированными окрашенными филлоидами. Между гаметангиями располагаются нитевидные или булавовидные парафизы. После оплодотворения яйцеклетки развивается спорогон, имеющий ножку, на верхушке которой образуется коробочка. Коробочка спорогония состоит из урночки и крышечки, развивается на гаметофите. Внутри урночки формируются спорангии со спорами. Для всех представителей листостебельных мхов характерна особая стадия развития гаметофита, развивающаяся из споры – протонема, резко отличающаяся от взрослого гаметофита. Подавляющее большинство видов мхов приурочено к сырым местам и играют большую фитоценотическую роль в растительном покрове. Класс делят на 3 подкласса – сфагновые, андреевые и бриевые мхи. III.1.2.1.Подкласс Сфагновые мхи – Sphagnidae В этом подклассе один род сфагн (Sphagnum), или сфагнум (рис. 19), насчитывающий 300 видов растений бледно-зеленого цвета, за что их называют белыми мхами. Сфагнум является характерным ландшафтным растением и торфообразователем. Он образует обширные массивы в тундре, на верховых болотах и зарастающих озерах, в сырых лесах и на низинных лугах. Высушенные верхние растущие части побегов видов рода сфагнум (Sphagnum) из сем. сфагновых – Sphagnaceae используют как лекарственное средство. Задание
Выделяют из дерновины одно растение сфагнума и рассматривают его, пользуясь лупой. Главный стебель, боковые и верхушечные ветви густо покрыты очень мелкими листьями. Одни боковые ветви оттопырены, другие свешиваются, прилипая к стеблю. Короткие ветви верхушки образуют густую пушистую головку желтой, бурой, коричневой или красноватой окраски. Сфагновые мхи стоят вертикально и неопределенно долго нарастают; нижние, старые части мха постепенно отмирают и буреют. Для сфагнума характерно разное строение стеблевых и веточных листьев. С боковой ветви препаровальной иглой соскабливают листья, изготовляют препарат и рассмат-ривают при малом и большом увеличениях микроскопа. Веточные листья округло-треугольной формы, остроконечные, прикрепляются к стеблю широким основанием. Они однослойные, без жилок. Лист состоит из двух типов клеток: слегка изогнутые живые хлорофиллоносные клетки; между ними находятся по одной и более крупной, мертвой, бесцветной, водоносной (гиалиновой) клетке, с кольчатыми утолщениями и порами на поверхности. Таким образом, в структуре листа сфагнума совмещаются две ткани – ассимиляционная и водоносоная. Листья разных видов сфагнума различаются деталями строения водоносных и хлорофиллоносных клеток. 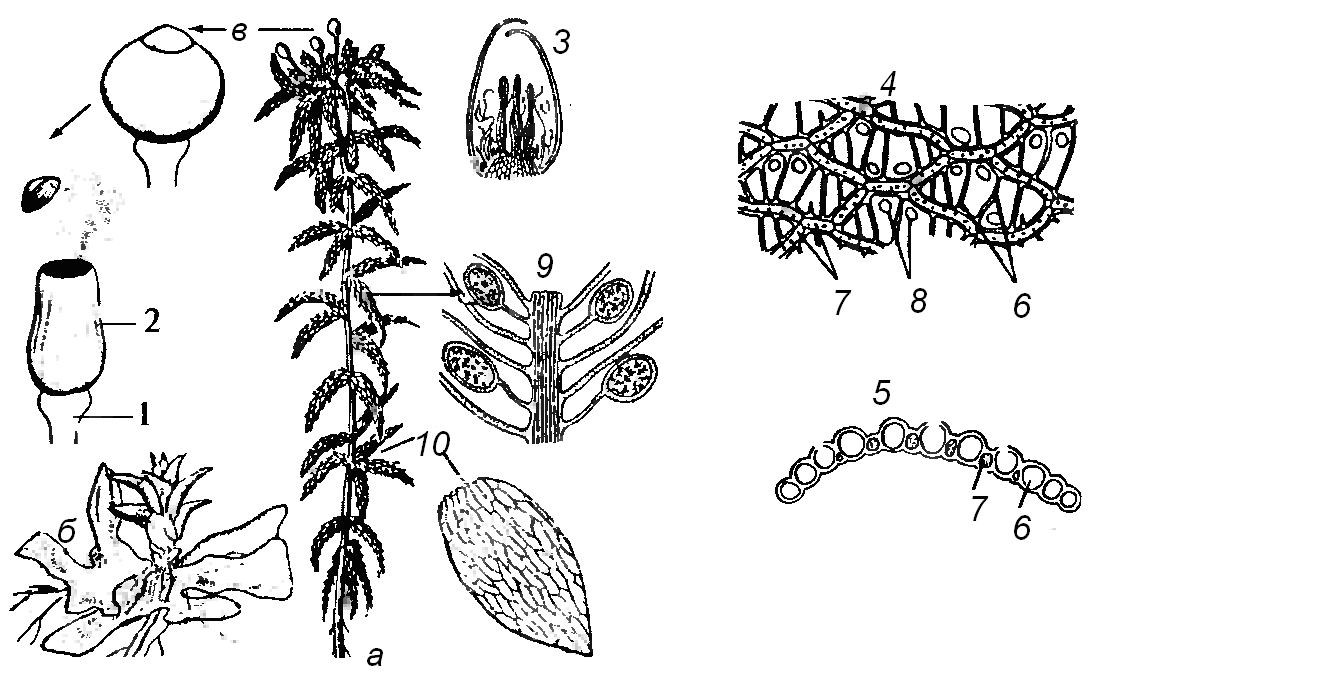 Рис. 19. Сфагнум: А – общий вид гаметофита со спорангиями; б – протонема; в – спорогоны; 1 – ножка спорогона; 2 – коробочк5а спорогона; 3 – архегоний; 4, 5, 10 – листочки гаметофита (вид под микроскопом с поверхности, поперечный разрез, внешний вид); 6 – водоносные клетки листа; 7 – фотосинтезирующие клетки; 8 – поры; 9 – антеридии Изготавливают препарат стебля сфагнума. Стебель зажимают в бузину и режут с помощью микротома. Стебель сфагнума покрыт корой или эпидермой, состоящей из нескольких (2–5) слоев клеток гиалодермы. Её крупные паренхимные тонкостенные клетки по типу гиалиновых клеток листа. Они имеют гиалиновые утолщения и сквозные отверстия на всех стенках клетки, поэтому вода может распространяться в соседние клетки во всех направлениях. К центру от гиалодермы располагается механическая ткань, состоящая из очень узких прозенхимных клеток с сильно утолщенными стенками бурого цвета и занимающая незначительную часть стебля. В центре находится паренхима сердцевины. Сфагнум является гаметофитом, однодомным или двудомным, у разных видов. Органы размножения образуются близ вершины, на ветвях головки. Антеридиальные веточки отличаются от вегетативных цветом, а также большей толщиной и более плотным расположением листочков, в пазухах которых на длинных ножках сидят овальные антеридии. Укороченные архегониальные веточки похожи на почки. На самой верхушке их, под прикрытием нежных кроющих листьев, сидят группами архегонии. Спорогоны созревают в середине лета. Шарообразный зрелый темно-коричневый спорогон открывается опадающей крышечкой. Делают срез, проходящий через коробочку и через ее короткую толстую ножку, вдающуюся в вырост веточки и приготовят микропрепарат. Этот вырост называют ложной ножкой. Значительную часть полости коробочки занимает крупноклеточный бугорок – колонка. На ней сидит куполообразный спорангий. Споры сфагнума крупные, округло-тетраэдрические, покрытые толстой желтой экзиной. Из спор сфагнума вырастают предростки (протонема) в виде зеленых лопастных пластинок с ризоидами. Из почек, появляющихся на протонеме, вырастают новые растения сфагнума. Протонема является начальной фазой гаметофита. Проделать опыт, показывающий водопоглощающие свойства сфагнума. Для этого поставить сухое растение сфагнума в стакан с водой. Через некоторое время побег пропитается водой, и верхушка его наклонится через край стакана, а вскоре с нее начнут стекать капли воды. Зарисовывают внешний вид сфагнума и участок листа с хлорофиллоносными и водоносоными клеткми. III.1.2.2. Подкласс Зеленые мхи – Bryidae Зеленые мхи отличаются от сфагновых отсутствием водоносных клеток в листьях и на поверхности стебля, наличием многоклеточных ризоидов, анатомическим строением стебля и более сложным строением спорогона, коробочка которого сидит на длинной ножке. Кукушкин лен(Polytrichumcommune)(рис. 20) – самый крупный из зеленых мхов; он широко распространен в сыроватых лесах и на болотах, где обитает рядом со сфагнумом, занимая относительно высокие участки микрорельефа. Почти чистые заросли кукушкина льна образуют в лесах высокие, пышные темно-зеленые подушки. Задание
Используя гербарий, рассматривают побеги мха и отмечают, что не все побеги несут спорогон. Он образуется только на верхушках женских гаметофитов. Следовательно, кукушкин лен – двудомное растение. Длинные, прямые и довольно крепкие стебли кукушкина льна густо покрыты сравнительно длинными, жестковатыми листьями. Более старые листья, отмирая, буреют и отваливаются. На подземной безлистной части стебля развиваются ризоиды – многоклеточные тонкие бесцветные нити. Отделяют от стебля несколько листьев, состоящих из линейной остроконечной пластинки с острыми шипиками по краям и широкого пленчатого влагалища, почти обхватывающего стебель. Листья сидят тесной спиралью, влагалища их находят одно на другое так, что стебель покрывается ими со всех сторон. Далее исследуют микроскопическое строение листа. На морфологической верхней стороне листа располагаются ассимиляционные пластинки. На нижней поверхности листа находится эпидермис. На вершине мужского растения находятся широкие красновато-желтые листья, образующие розетку, в центре которой между парафизами сидят антеридии на толстых многоклеточных ножках. Перемещение сперматозоидов на женские растения происходит обычно во время дождя. Мужские растения после опадения антеридиев продолжают рост, оставляя на стебле листочки розетки.  Рис. 20. Схема развития кукушкина льна: 1 – мужской и женский гаметофиты; 2 – антеридий со сперматозоидами; 3 – сперматозоид; 4 – архегоний с яйцеклеткой; 5 – развитие зиготы из оплодотворенной яйцеклетки; 6 – развивающийся спорофит; 7 – зрелые спорофиты на женских гаметофитах: а – ножка; б – коробочка; в – крышечка; д – колонка; е – спорогенные клетки спорангия и образование из них спор (мейоз); 8 – споры (+ и –); 9 – прорастание спор; 10 – протонемы мужская и женская На женских растениях на конце побегов располагаются спорогоны. Зрелый спорогон состоит из коробочки, прикрытой волокнистым колпачком, и длинной ножки, основание которой, глубоко внедряется в верхушку женского растения и является гаусторией. Рассматривают постоянный препарат продольного разреза коробочки кукушкина льна. Коробочка состоит из урночки, в которой созревают споры и крышечки с заостренной верхушкой. В сухую погоду через щели перистома споры вытряхиваются из коробочки и подхватываются течениями воздуха, и таким образом рассеиваются. На сырой почве споры прорастают в предросток (протонему). Микроскопически протонема зеленых мхов похожа на нитчатые зеленые водоросли. Ветви протонемы, лишенные хлорофилла, являются ризоидами. На протонеме возникают почки, из которых вырастают взрослые растения. Зарисовывают внешний вид кукушкина льна и спорогон. СОСУДИСТЫЕ СПОРОВЫЕ РАСТЕНИЯ К этой группе относятся растения с проводящей системой, которые в отличие от семенных растений расселяются спорами. Общая черта развития этих растений заключается в том, что в жизненном цикле этих растений чередуются хорошо развитый спорофит и маленький просто устроенный гаметофит – заросток. Спорофиты преобладают по размерам и сложности строения над гаметофитами. У большинства сосудистых споровых растений имеются хорошо развитые вегетативные органы: стебли, листья, корни и проводящие ткани. III.2. Отдел Плауновидные – Lycopodiophyta Плауновидные – многолетние, вечнозеленые растения с простыми мелкими или чешуевидными спирально расположенными листьями, характеризующиеся дихотомическим ветвлением. Стебель хорошо развит. Побеги и корни характеризуются самым древним – дихотомическим типом ветвления. Для плауновидных характерно отсутствие главного корня, все корни занимают на побегах боковое положение. Спорангии плауновидных формируются на верхней стороне спорофиллов, которые образуют специализированные спороносные побеги – стробилы в виде колосков. Отдел плауновидные разделяется на 2 класса - плауновидные (Lycopodiopsida) равноспоровые и полушниковые (Jsoetopsida) разноспоровые. III. 2.1. Класс Плауновые – Lycopodiopsida Оборудование и материалы: микроскопы, постоянные микропрепараты продольных срезов спороносных колосков, гербарные образцы плауна булавовидного, п. сплюснутого, п. годичного, засушенные спороносные колоски плауна со спорами. Плауновые встречаются в сыроватых и тенистых хвойных и смешанных лесах, но некоторые виды, например плаун сплюснутый (Lycopodiumcomplanatum) (рис. 22), произрастает в сухих сосновых лесах. В России плауновидные представлены 14 видами, в Чувашии – 4 вида. Собранные в июле – сентябре и высушенные споры вечнозеленых многолетних травянистых растений плауна булавовидного (Lycopodiumclavatum), п. годичного (Lycopodiumannotinum) (рис. 22), п. сплюснутого (Lycopodiumcompla-natum) из сем. плауновых (Lycopodiaceae) используют в качестве лекарственного средства. Наиболее распространенным и типичным представителем плауновых является плаун булавовидный (Lycopodiumclavatum). Задание
С особенностями плауновых знакомятся на примере плауна булавовидного (Lycopodiumclavatum) (рис. 21), который является типичным представителем и наиболее распространенным в хвойных лесах. При морфологическом описании плаунов обращают внимание на следующие признаки: собраны ли спорофиллы в колосок (стробил) или спорофиллы не образуют колосок; одиночные колоски собраны по 2 или больше; стебли и ветви цилиндрические или сплюснутые и их характер ветвистости; форму спорофиллов и др. признаки. Рассматривают растение и отмечают, что у плауна булавовидного стебли ползучие, длинные, неравномернодихотомически ветвящиеся, с восходящими густо олиственными побегами высотой 10 – 30 см, заканчивающимися 2 – 4 стробилами – «спороносными колосками». На ползучем стебле плауна располагаются редкие тонкие и прочные корни. Стебель и корни ветвятся дихотомически. Стебель густо покрыт листьями. Отделяют препаровальной иглой лист и рассматривают его при помощи стереоскопического микроскопа. Листья плауна булавовидного сидячие линейно-цельнокрайние, заканчиваются длинной шиловидной верхушкой. Вдоль листа проходит средняя жилка. Зарисовывают лист. Далее изучают спороносные колоски, венчающие вертикальные побеги. Они расположены на довольно длинных ножках по 2. Колосок цилиндрической формы, состоит из оси, на которой плотно расположены спорофиллы. Каждый спорофилл представляет собой чешуевидный треугольный лист с заостренным и загнутым кверху концом. Выделяют один спорофилл и рассматривают на верхней стороне его почковидный спорангий, сидящий на короткой ножке. Спорангий вскрывается продольной щелью. Зарисовывают часть колоска и спорофилл со спорангием. Строение колоска также рассматривают, пользуясь постоянным препаратом его продольного среза.  Рис. 21. Строение и схема цикла развития плауна булавовидного: а – спорофит; б – гаметофит; 1 – ползучий побег с придаточными корнями; 2 – восходящие побеги; 3 – ножка спороносных колосков; 4 – листочки: а - восходящего побега; б – ножки спороносных колосков; 5 – спороносмные колоски; 6 – споролистики: в – вид с брюшной, г – спинной сторон; 7 – спорангии; 8 – споры; 9 – прорастающая спора; 10 – архегоний; 11 – антеридий; 12 – оплодотворение; 13 – оплодотворенная яйцеклетка; 14 – развитие нового спорофита на гаметофите Дальше приступают к изучению спор. Раздавливают спорангий на предметном стекле, рассматривают споры в микроскоп и зарисовывают. Все споры одинаковые, мелкие и имеют округлотетраэдрическую форму со сферическим основанием и бледно-желтого цвета. Поверхность спор неровная, с выступающими сетчатыми утолщениями оболочки, в которых скапливается воздух, не позволяющий спорам смачиваться водой и тонуть в ней, хотя они и тяжелее воды. В клетке споры, кроме протопласта и ядра, имеются пластиды и капельки масла (до 50%). Внесенный осторожно в пламя порошок горит тихо и ровно, брошенный сверху в пламя сгорает мгновенно, с яркой вспышкой, бездымно. Из спор плаунов вырастают бесцветные обоеполые заростки, развитие которых происходит в почве и только при наличии микоризного гриба. Продолжительность жизни заростков плаунов – 12 – 15 лет и более. После оплодотворения развивается зародыш, при дальнейшем развитии преобразующийся в молодое растение. Плаун годичный отличается от плауна булавовидного более жесткими оттопыренными листьями и одиночными сидячими спороносными колосками. Плаун сплюснутый (рис. 19) имеет веерообразно расположенные дихотомически разветвленные веточки с чешуевидными прижатыми листьями и 3 – 4 стробила, сидящих на коротких ножках. 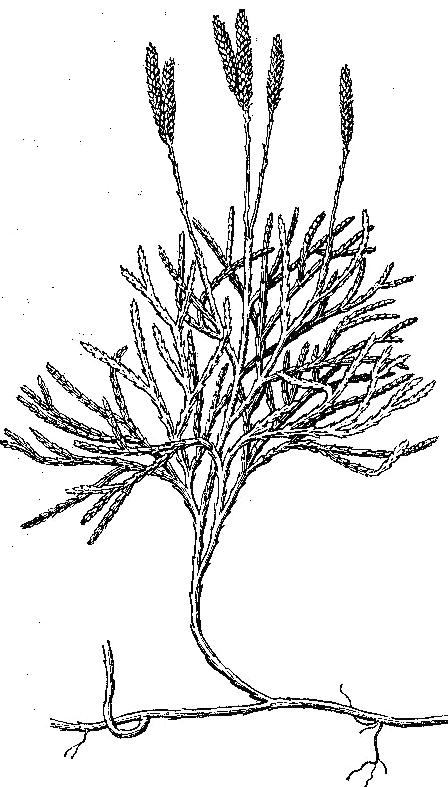 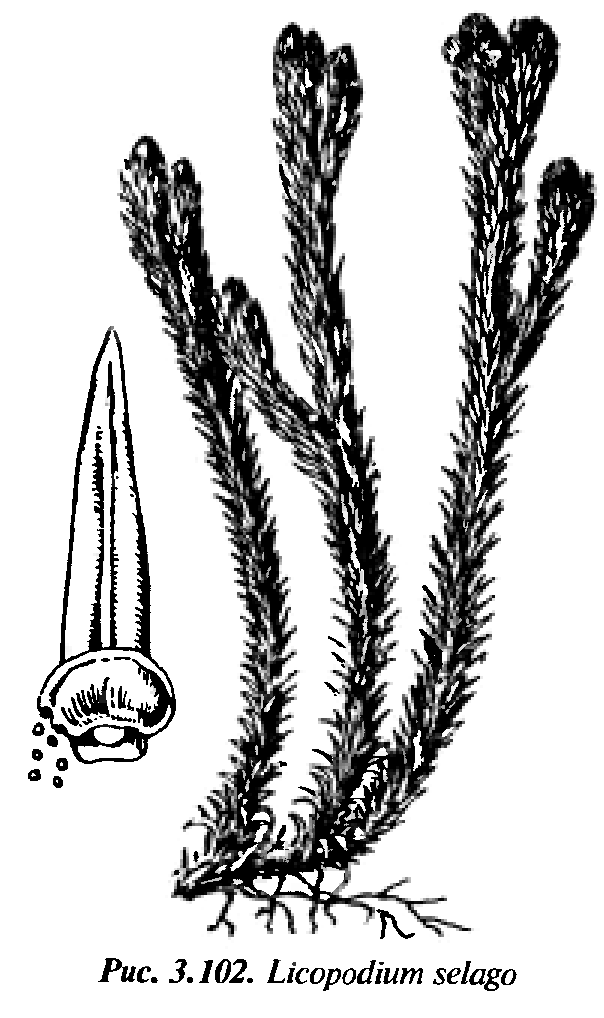 а б Рис. 22. Плауны сплюснутый (а) и годовалый (б) III.3. Отдел Хвощевидные, или Членистые – Equisetophyta, или Sphenophyta Оборудование и материалы: живые и гербарные образцы хвоща полевого, хвоща лугового, хвоща лесного, хвоща зимующего; заспиртованные или засушенные спороносные колоски этих растений; постоянные микропрепараты продольных срезов спороносных колосков; микроскопы. У хвощевидных очень характерная специфика строения их побегов. Побеги хвощей расчленены на четко выраженные узлы и междоузлия, легко разламывающиеся в узлах на членики. Членистость обусловлена мутовчатым листорасположением и наличием в нижних частях междоузлий интеркалярной меристемы, по которой происходит разламывание на членики. Листья зеленые или бесцветные, сильно редуцированные, чешуевидные срослись в общее пленчатое влагалище. Оно плотно охватывает стебель и защищает меристематические ткани. Ассимиляционную функцию выполняют зеленые стебли, поверхность которых увеличивается ребристостью, стенки клеток кожицы пропитаны кремнеземом. Подземная часть хвощей представлен сильно развитым корневищем, в узлах которого формируются придаточные корни. У некоторых видов боковые ветви корневищ превращаются в клубни, которые служат местом отложения запасных продуктов, а также органом вегетативного размножения. Для репродуктивных органов хвощевидных характерно наличие стробилов в виде колосков. Спороносный колосок состоит из спорангиофоров, не похожих на листья. Спорангиофоры, т. е. органы, несущие спорангии, имеют щитковидную форму. Кроме обычных двух оболочек, споры имеют ленточки - элатеры, закрученные вокруг споры. Хвощевидные – равноспоровые растения. У разных видов хвоща по-разному дифференцированы спороносные и вегетативные побеги. У хвоща приречного (Equisetumfluviatile) они не различаются, и спорангии образуются поздно на верхушках побегов. До этого невозможно определить, на каком побеге будут закладываться спороносные колоски. У хвоща полевого (E. arvense) спороносные и вегетативные побеги резко отличаются (рис. 23, б). Розово-бурые неветвящиеся спороносные побеги появляются рано весной и развиваются за счет запасов питательных веществ в клубеньках корневища. После созревания спор и усыхания спороносных побегов появляются вегетативные побеги. У хвоща лесного (E. sylvaticum), хвоща лугового (E. pratense) (рис. 23, а) одновременно появляются вегетативные и спороносные побеги, и после рассеивания спор спороносные побеги функционируют как вегетативные.  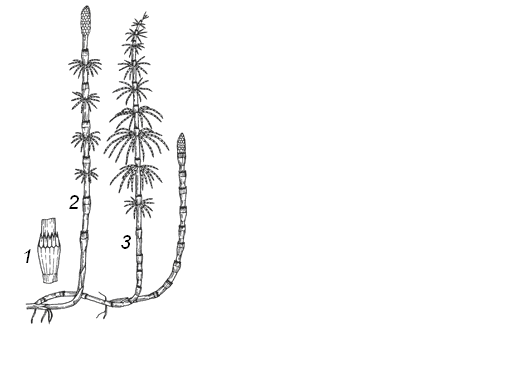 а б Рис. 23. Хвощи: а – хвощ лесной; б – хвощ луговой; 1 – листовое влагалище; 2 – спороносный побег; 3 – вегетативный побег У хвощей сочетаются ксероморфные и гигроморфные признаки. Как у ксерофитов, у хвощей редуцированы листья – они мелкие и незеленые. Устойчивость к засухе обеспечивается сложно устроенными устьицами, регулирующими испарение, и развитой механической тканью. Эпидерма сте- блей хвощей содержит кремнезем. Заростки развиваются в течение нескольких недель, и после перекрестного оплодотворения (у обоеполых заростков – за счет неодновременного созревания антеридиев и архегониев) образуют молодые спорофиты. III.3.1. Класс хвощевые - Equisetopsida Современные хвощовые представлены только одним порядком (Equisetales), одним семейством хвощевых (Equisetaceae) и одним родом хвощ (Equisetum) с 25 космополитными видами. В России насчитывается 12 видов, а также ряд гибридов между ними, в Чувашии – 6 видов. При определении и описании хвощей используют следующие основные морфологические признаки: а) стебли растений одинаковые или спороносные стебли отличаются от стерильных (вегетативных); б) спороносные и стерильные стебли появляются одновременно или спороносные стебли появляются рано весной; в) форма спороносного колоска: тупой на конце или заостренный; г) форма стеблевого влагалища, а также его размер и число зубцов; д) степень ветвистости стебля, его форма роста и высота, наличие или отсутствие полости, а также продолжительность жизни (однолетний или многолетний). Задание
ния.
Делают морфологическое описание хвоща полевого, наиболее широко распространенногое растения из хвощей. Хвощ полевой (рис. 23, б; 24) – многолетнее растение с членистым ветвистым корневищем, проникающим в почву до 100 – 120 см. На корневище имеются клубни, достигающие величины лесного ореха, содержащие запасной крахмал. Они возникают как укороченные боковые побеги. От узлов корневища отходят пучки тонких черных корней. 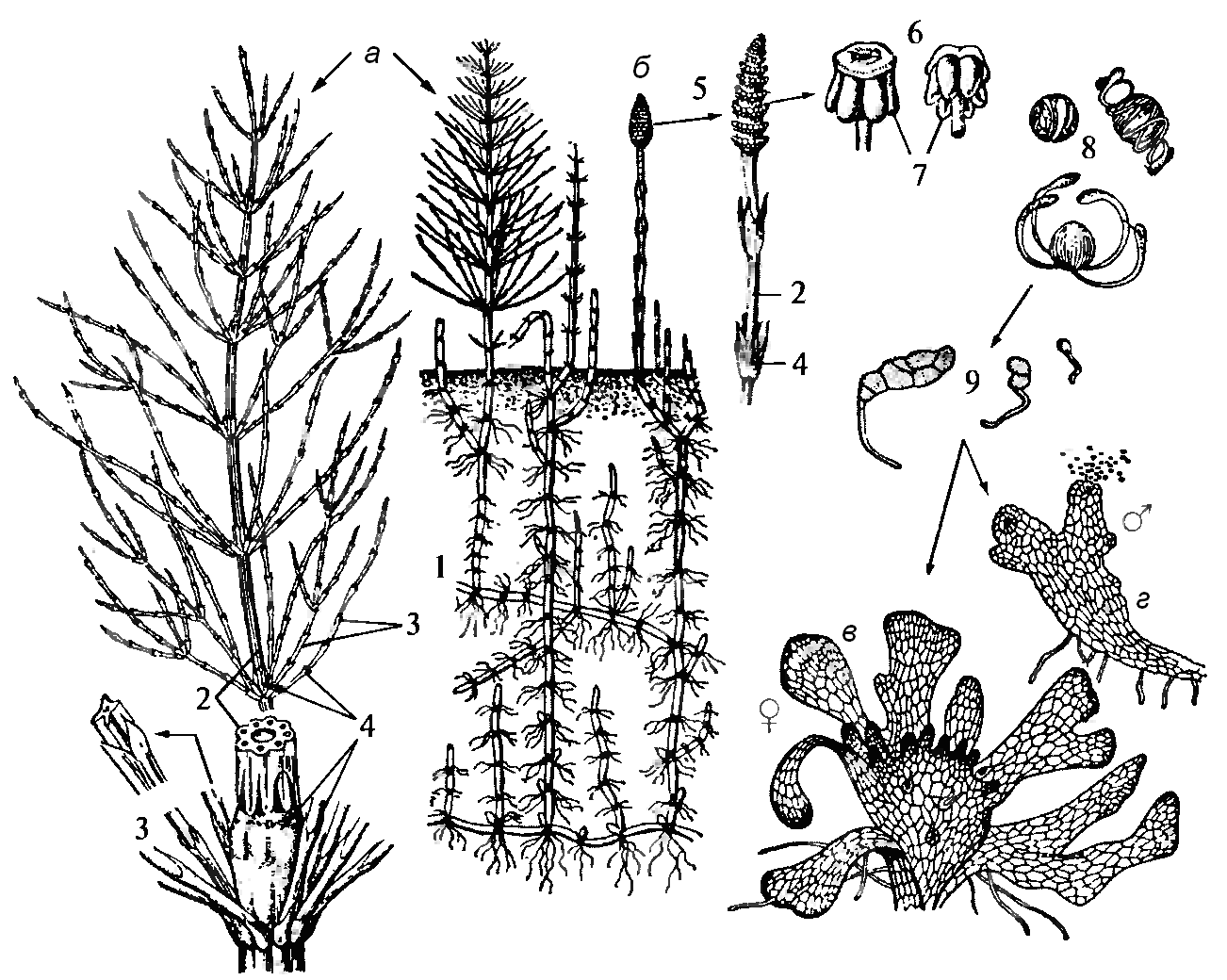 Рис. 24. Строение и цикл развития хвоща полевого: а – вегетативный побег; б – спороносный побег; в – женский гаметофит; г мужской гаметофит; 1 – корневище с клубеньками; 2 – стебельт; 3 – боковые веточки; 5 – спороносный колосок; 6 – спорангиофор; 7 – спорангии; 8 – споры с элатерами; 9 – прорастающие споры Наземные побеги, восходящие от корневища, двоякие: ранневесенние – розовато-бурые, без хлорофилла, спороносные, после созревания спор отмирающие; и поздне-весенние – стерильные, мутовчато-разветвленные, зеленые, вегетирующие. Рассматривают сначала строение зеленого, ветвистого летнего побега. Отделяют от главного стебля один его членик, состоящий из длинного междоузлия и узла. Членик отрывается легче всего над самым узлом, где находится неокрепшая ткань зоны роста междоузлия. На нижнем конце членика открывается крупная полость междоузлия, отгороженная на другом конце сплошной перегородкой узла от соседнего междоузлия. От узла вверх направлена мутовка мелких острых бурых листьев, сросшихся основаниями в трубку, которая наподобие острозубчатого воротничка охватывает нижнюю часть междоузлия, образуя цилиндрическое влагалище. Стерильные побеги бороздчатые, ветвистые, зеленые, ассимилирующие. Функции ассимиляции выполняют также многочисленные зеленые боковые ветви, сидящие мутовками. Они закладываются в пазухах листьев и, развиваясь прорывают влагалище. Рассматривают в лупу боковую ветку хвоща и сравнивают ее с главным стеблем и отмечают, что их строение одинаковое, членистое. В лупу хорошо видно, что поверхность междоузлий продольнобороздчатая. Число бороздок, чередующихся с параллельными ребрышками, равно числу листьев, т. е. зубцов влагалища. Прослеживают направление ребрышек при переходе с одного междоузлия к другому и отмечают, что ребрышки на одном междоузлии продолжают в бороздки на другом, и наоборот. Хвощ полевой размножается вегетативно, развивая подземные побеги. В поле при механической обработке почвы подземные побеги хвоща разрываются на части, которые продолжают развиваться как отдельные растения, усиливая засоренность поля. Размножение при помощи спор связано с появлением весенних побегов. Они также членистые, не зеленые, а розовато-бурые, не ветвящиеся, со спороносными колосками на верхушках. Весенний побег имеет колосок и его подставку. Рассматривают мутовки сросшихся листьев, лишенных хлорофилла и образующих влагалища у оснований междоузлий. На подставках влагалища вздутые и более крупные, чем на летних побегах. При помощи стереоскопического микроскопа рассматривают живые или заспиртованные спороносные колоски. Колосок состоит из правильных шестигранных щитков, расположенных горизонтальными мутовками. Извлекают пинцетом из колоска один щиток. Щиток имеет ножки прикрепляющей его к стержню колоска. Вокруг ножки по краю свисают мешковидные спорангии, расположенные по нижнему краю щитка, раскрывающиеся щелью на внутренней, обращенной к ножке стороне. Щиток с ножкой является спорофиллом хвоща. Зарисовывают общий вид колоска и спорангиофор и делают обозначения. Строение колоска можно изучить и на постоянном препарате. Затем изготавливают препарат спор. Для этого надо засушенным колоском постучать по предметному стеклу, чтобы высыпалось немного спор. Не закрывая их покровным стеклом и не добавляя воды, рассматривают в микроскоп. Затем, слегка подышав на споры, вновь рассматривают под микроскопом. Хвощи – равноспоровые растения. Споры их имеют не две, а три оболочки. Кроме экзины и и интины, они несут еще одну наружную оболочку – перину, из которой при созревании спор образуются две спиральные ленты – элатеры с небольшим расширением на концах. Элатеры способствуют рассеиванию спор и обеспечивают распространение их целыми группами. Зарисовывают 2 – 3 споры. При определении хвоща полевого указать на признаки, отличающие его от близких видов, используя таблицу 2.. Далее составляют схему жизненного цикла хвоща. Таблица 2
III.4. Отдел Папоротниковидные или Папоротники – Polypodiophyta Папоротники насчитывают около 300 родов и более 10 тысяч видов, из которых около 160 видов встречается в России, в Чувашии – 16 видов. В нашей флоре произрастают только многолетние травянистые папоротники. Они широко распространены по всему земному шару и встречаются в самых разнообразных местообитаниях. Для папоротниковидных характерны: крупнолистность (макрофилия), отсутствие стробилов (спороносного колоска), отсутствие камбия, в местах отхождения сосудистых пучков из стебля в листья (вайи) имеются листовые прорывы. В жизненном цикле папоротников преобладает спорофит – многолетнее травянистое растение с подземным корневищем. У папоротников по массе преобладают листья, которые и составляют наиболее заметную часть спорофита. У большинства видов папоротников листья совмещают две функции – фотосинтеза и спороношения. Листья – вайи крупные, в почке сложены улиткообразно, нарастают верхушкой и несут спорангии. Молодые вайи покрыты волосками и чешуйками. Большинство лесных папоротников умеренной зоны имеют мясистые, сифоностелические корневища, на которых каждый год образуются новые пучки листьев. Листья чаще всего перистые. Размножение папоротников осуществляется в основном за счет спор. Большинство видов папоротников – равноспоровые растения, спорангии находятся на нижней стороне листа, и они приурочены к главной или боковым жилкам, реже (у орляка) – близ края листа. Спорангии собраны в группы – сорусы, и их созревание начинается от центра группы. Групповое положение спорангиев позволяет обеспечить их лучшее снабжение элементами питания за счет крупного проводящего пучка, подходящего к сорусу. Сорусы защищены особыми выростами листа – покрывальцами, или индузиями. Расположение спорангиев на нижней стороне листа оказывается биологически выгодным: 1) обеспечивается надежная защита спорангиев во время их созревания и одновременно не снижается интенсивность фотосинтеза; 2) обеспечивается более равномерное рассеивание спор; 3) создается возможность большего продуцирования спорангиев на всей поверхности листа, чем только по краю. Споры папоротников имеют многослойную оболочку и хорошо переживают неблагоприятные условия, длительное время сохраняя способность к прорастанию. Для их прорастания необходимы влага, положительные температуры, определенная кислотность почв, интенсивность и качество света, специфичное для каждого вида. Споры равноспоровых папоротников дают обоеполым гаметофитам (заросткам). Они могут быть автотрофными зелеными или гетеротрофными. Автотрофные гаметофиты большинство папоротников пластинчатые с ризоидами и имеют сердцевидную форму, размером около 5 мм. Несмотря на обоеполость таких гаметофитов, самооплодотворение случается редко, так как антеридии появляются раньше. Они располагаются в области ризоидов, а архегонии – вблизи выемки на переднем конце заростка. Разница во времени формирования антеридиев и архегониев способствует перекрестному оплодотворению, в итоге чего появляются более жизнеспособный спорофит. Отдел папоротниковидные делят на 9 классов, только представители пяти классов, из которых только представители пяти существуют в настоящее время. Задание 1. Рассмотреть щитовник мужской, кочедыжник женский, страусник обыкновенный, орляк по гербарным экземплярам и сравнить их между собой. 2. Изучить щитовник мужской. Составить краткое морфологическое описание. Зарисовать вайю. 3. Рассмотреть под микроскопом поперечный срез соруса. Схематично зарисовать участок вайи с сорусом и спорангий, обозначить их части 4. Исследовать и зарисовать заросток. 5. Определить рассмотренные растения и указать на важнейшие отличия его от наиболее близких видов. Составить схему жизненного цикла папоротника. С особенностями папоротниковидных знакомятся на примере щитовника мужского (Dryopterisfilix-mas) (рис. 25), одного из самых широко распространенных папоротников в смешанных и широколиственных лесов. Собранные осенью или ранней весной, очищенные от корней и отмерших листьев, с оставленными основаниями черешков, высушенные корневища дикорастущего многолетнего травянистого растения щитовника мужского из сем. аспидиевых (Aspidiaceae), используют в качестве лекарственного сырья. Рассматривают гербарный образец щитовника мужского. Это многолетнее споровое растение с косо растущим коротким корневищем с хорошо выраженным дорзовентральным строением, верхняя поверхность несет черешки отмерших вай, нижняя – тонкие придаточные корни. Цвет корневища и покрывающих его черешков снаружи черно-бурый От верхушки корневища отходит пучок зеленых листьев – вай, основания черешков покрыты одноцветными, светло-бурыми пленчатыми чешуйками, несущими по краю сдвоенные выступы-зубчики. Пластинка вайи в очертании эллиптически-продолговатая, дважды перисторассеченная. Сегменты первого порядка расположены поочередно, заостренные, сегменты второго порядка с зубчатым краем и тупой верхушкой. На нижней поверхности вай, вдоль жилок сегментов второго порядка, расположены сорусы (пучки спорангиев), прикрытые почковидными покрывальцами - индузиями. Зарисовывают общий вид вайи и один из сегментов с сорусами. 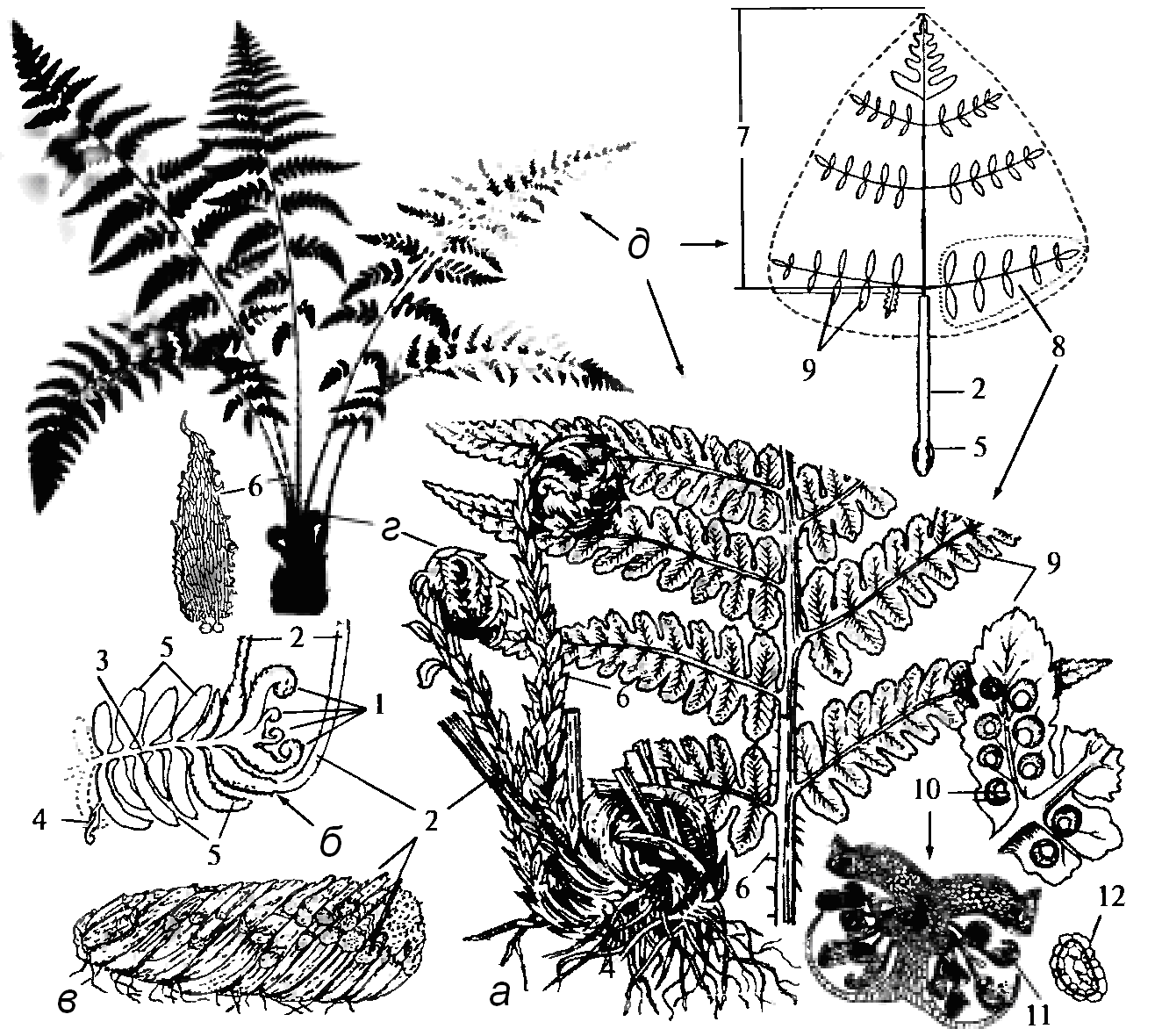 Рис. 25. Щитовник мужской: а, б, в – корневище: внешний вид, схема строения, сухое сырье; г – молодые листья; д – зрелые вайи (внешний вид, схема строения, врагмент нижней части); 1 - зачаточные листочки верхушечной почки корневища; 2 – черешок; 3 – осевая часть корневища; 4 – придаточные корни; 5 – утолщенные основания вай; 6 – пленчатые чешуйки черешка; 7 – рахис; 8 – сегменты (листочки) первого порпядка; 9 – сегменты (листочки) второго порядка; 10 – сорусы (внешний вид, разрез); 11 – спорангии; 12 – споры Затем закладывают сегмент вайи, заранее размоченный в теплой воде в сердцевину бузины, делают несколько поперечных срезов через сорус на микротоме и изготавливают препарат. Можно воспользоваться и готовым, постоянным препаратом. Рассматривают срез соруса при малом и при большом увеличениях микроскопа. В центре соруса находится массивный вырост листа – плацента, которая, расширяясь, переходит в тонкий зонтиковидный индузий, слегка вдавленный в середине. Под индузием находится группа спорангиев, прикрепленных к плаценте довольно длинными ножками. Если разрез прошел через середину плаценты, то можно видеть проводящий пучок в продольном разрезе, идущий из листа в покрывальце. Зарисовывают поперечный разрез вайи с сорусом. Рассматривают отдельно спорангий, который имеет форму чечевицы. Стенка спорангия многоклеточная, однослойная. По ребру спорангия выделяется ряд клеток с неравномерным утолщением стенок, узкой полосой опоясывающий его. Это механическое кольцо. Детали строения покрывальца, спорангиев и кольца различны у разных видов папоротников и являются важными систематическими признаками при их определении. Споры имеют почковидно-овальную форму и толстую бугорчатую стенку. Споры в спорангиях папоротника, образуются из материнских клеток путем мейоза. Спора является первой клеткой нового гаметофита. Зарисовывают спорангий с механическим кольцом и несколько спор. Из споры, попавшей в благоприятные условия, развивается маленький, эфемерный, самостоятельно вегетирующий заросток – гаметофит (рис. 26). Он обоеполый, до 4-5 мм в поперечнике, с сердцевидной выемкой. Под микроскопом хорошо видно, что на нижней стороне, вблизи от выемки, архегонии с брюшком, погруженными в ткань заростка, а в ризоидальной части – округлые антеридии. После оплодотворения из зиготы, находящейся в брюшке архегония, развивается зародыш, состоящий из зачаточных органов нового спорофита папоротника – стебля, корня и листа. Молодой спорофит папоротника укореняется в том месте, где находился заросток и происходило оплодотворение. Рассматривают общий вид заростка сначала в лупу, а затем под микроскопом. Зарисовывают заросток, делают обозначения и составляют схему жизненного цикла папоротника. Рассматривают отличительные признаки других видов папоротников. Щитовник игольчатый (Щ. шиповатый) – Dryopteriscarhtusiana (D. spinulosa) имеет более мелкие корневища,  Рис. 26. Цикл развития папоротника. чешуйки по краю усажены головчатыми волосками. Листья дважды и трижды перисторассеченные, в очертании узко-треугольной формы, краевые зубцы вытянуты в мягкую иголочку. Встречается часто в хвойных лесах. У щитовника австрийского (Dryopterisaustiaca(Ddilatata) листья трижды и четырежды перисторассеченные, в очертании треугольные, самый нижний сегмент второго порядка, направленный к основанию листа, значительно длиннее других. Основания черешков покрыты бурыми чешуйками с широкой продольной темной полоской. Встречается редко в хвойных лесах. Корневища кочедыжника женского (женский папоротник) – Athyriumfilix-femina(рис. 27, б)почти вертикальные,усажены основаниями черешков, имеющими трехгранную форму и почти черную окраску, чешуйки цельнокрайние.   а б Рис. 27. Папоротники: а – страусник обыкновенный; б – кочедыжник женский Листья нежные трижды перисторассеченные, с мелкими зубчатыми сегментами, на коротких черешках. Сорусы отличаются продолговатой формой. Страусник обыкновенный (страусопер) (рис. 27, а) – Matteucciastruthiopteris из сем. Onocleaceae имеет листья похожие на мужского щитовника, но не несут сорусов и растут, образуя воронку, в центре которой находятся несколько коротких, светло-зеленых, а при созревании спор – бурых спороносных листьев. Корневище толстое, вертикальное, овальное в очертании. Чешуйки, покрывающие основания черешков трегранные, темно-бурые, цельнокрайние. | ||||||||||||||||||||||||||||||||||||||||||||||