Модуль 1 1 Потенціал спокою
 Скачать 1.25 Mb. Скачать 1.25 Mb.
|

|
5)Механізми проведення збудження через нервово-м’язовий синапс. Медіатор, мембранні циторецептори та блокатори нервово-м’язових синапсів. Н 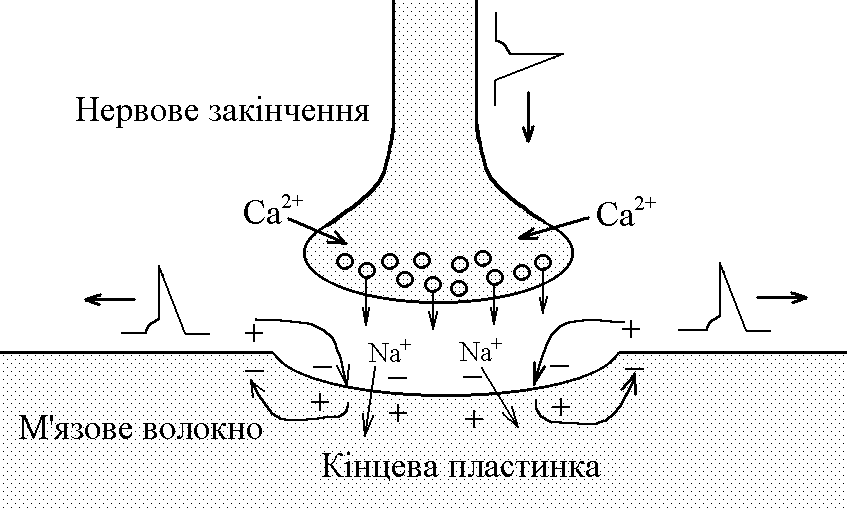 ервово-м’язовий синапс утворений нервовим закінченням аксона -мотонейронів та кінцевою пластинкою – частина мембрани м’язового волокна, яка контактує з нервовим закінченням. Розділяє пресинаптичну мембрану нервового волкна та кінцеву пластинку синаптична щілина. ервово-м’язовий синапс утворений нервовим закінченням аксона -мотонейронів та кінцевою пластинкою – частина мембрани м’язового волокна, яка контактує з нервовим закінченням. Розділяє пресинаптичну мембрану нервового волкна та кінцеву пластинку синаптична щілина.Механізм передачі збудження через нервово-м’язовий синапс полягає в тому, що ПД іде по мембрані нервового волокна поширюється по пресинаптичній мембрані, при цьому відкриваються кальцієві канали пресинаптичної мембрани вхід іонів Са+ всередину нервового закінчення взаємодія з везикулами (міхурці, в яких є медіатор ацетилхолін) рух везикул до пресинаптичної мембрани злиття везикул з пресинаптичною мембраною вихід медіатора ацетилхоліну в синаптичну щілину дифузія ацетилхоліну до мембрани кінцевої пластинки взаємодія з мембранними циторецепторами (Н-холінорецептори) відкриття хемочутливих натрієвих каналів вхід іонів Na+ в м’язове волокно через кінцеву пластинку розвиток деполяризації кінцевої пластинки, що має назву потенціалу кінцевої пластинки (ПКП). ПКП – один із видів місцевого збудження.Я 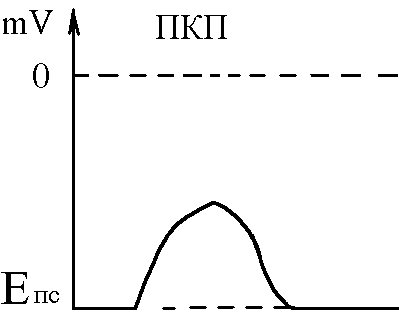 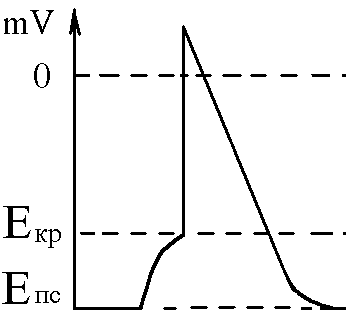 к будь-яке з місцевих збуджень ПКП поширюється на сусідні ділянки постсинаптичної мембрани за допомогою місцевих струмів. Ці струми в незбуджених ділянках мембрани мають вихідний напрям, тобто викликають деполяризацію. к будь-яке з місцевих збуджень ПКП поширюється на сусідні ділянки постсинаптичної мембрани за допомогою місцевих струмів. Ці струми в незбуджених ділянках мембрани мають вихідний напрям, тобто викликають деполяризацію.Cила цих струмів чисельно відповідає амплітуді ПКП, а вона складає 45-50 мВ. Поріг деполяризації постсинаптичної мембрани м’язового волокна складає 40 мВ (величина ПС = –90 мВ, величина Екр = –50 мВ. Отже, сила цих струмів надпорогова. Час дії цих струмів дорівнює тривалості ПКП і також є надпороговим. Швидкість збільшення сили відповідає швидкості розвитку ПКП й також є надпороговою. Отже, місцеві струми в незбуждених ділянках постсинаптичної мембрани викликають деполяризацію до Екр розвиток ПД, які будуть поширюватися від місця свого виникнення вздовж усієї довжини мембрани м’язового волокна (і через електромеханічне спряження будуть викликати скорочення м’яза). Закономірності проведення збудження через нервово-м’язовий синапс: 1. Однобічне проведення. 2. Уповільнене проведення (час від виникнення ПД на пресинаптичній мембрані до виникнення ПД на постсинаптичній мембрані складає близько 0,5 мсек. 3. Проведення збудження через синапс супроводжується швидким розвитком втоми (у зв’язку з виснаженням запасів медіатора в нервовому закінченні); Ці особливості (1-3) характерні для будь-якого хімічного синапса і пов’язані з хімічним характером передачі інформації через синапс. 4. Збудження передається через нервово-м’язовий синапс без трансформації ритму: 1 ПД на пресипаптичній мембрані викликає 1 ПКП на мембрані кінцевої пластинки, а це викликає 1 ПД на мембрані м’язового волокна. Блокаторами нервово-м’язової передачі є курареподібні речовини, паприклад, диплацин, що утворюють стійкі з’єднання з Н-холінорецепторами та не дають змогиацетилхоліну взаємодіяти з ними. Медіатор ацетилхолін взаємодіє з рецепторами протягом короткого – часу, після чого комплекс медіатор-циторецептор розпадається і медіатор руйнується ферментом ацетилхолінестеразою, активність якого в синапсі дуже висока. 6)Механізми скорочення та розслаблення скелетних м'язів Т 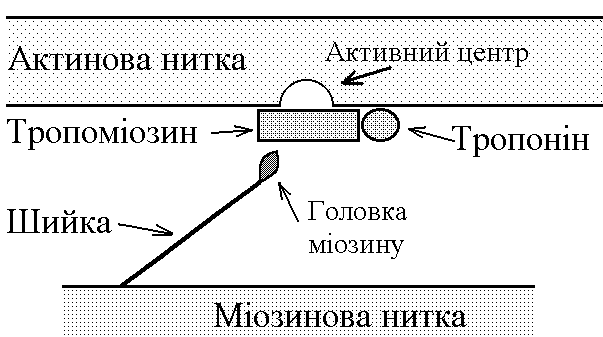 ермін “спряження збудження із скороченням” означає взаєзв’язок збудження в скелетних м’язах (виникнення та поширення ПД по мембрані волокна) та його скорочення, тобто актоміозинової взаємодії. ермін “спряження збудження із скороченням” означає взаєзв’язок збудження в скелетних м’язах (виникнення та поширення ПД по мембрані волокна) та його скорочення, тобто актоміозинової взаємодії.В стані спокою взаємодії актинових та міозинових протофібрил немає тому, що активні центри актину заблоковані регуляторними білками тропоніном та тропоміозином. Знімають блокаду іони Са2+, концентрація яких в саркоплазмі в стані спокою низька 10–8 ммоль/л. ПД, що виникають в постсинаптичній мембрані поширюються вздовж всієї довжини мембрани м’язового волокна, у тому числі і по мембрані Т-трубочок (це вгинання зовнішньої мембрани м’язових клітин всередину волокна). Б 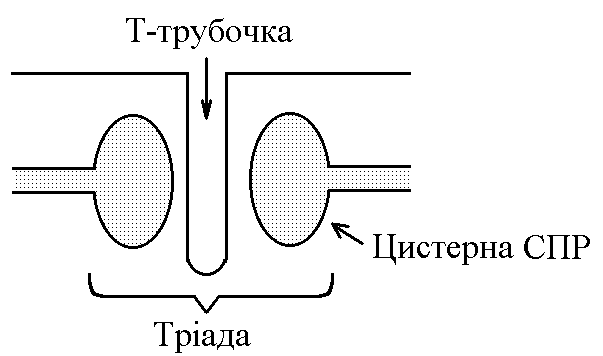 езпосередньо поруч з Т-трубочками розташовані цистерни саркоплазматичного ретикулуму (СПР – частина аграпулярної епдоплазматичної сітки). Т-трубочки з двома поруч розташованими цистернами утворюють тріади. СПР крім цистернмаютьпоздовжні трубочки, в яких локалізуються кальцієві насоси – вони активно транспортують іони – Са2+із саркоплазми в СПР і в його цистернах накопичується велика кількість іонів Са2+. При русі ПД по мембрані Т-трубочок в мембрані цистерн СПР відкриваються кальцієві канали іони Са2+ по градієнту концентрації виходять з цистерн СПР у саркоплазму підвищення концентрації іонів Са2+ в саркоплазмі міоцита з 10–8 до 10–5 ммоль/лдифузія іонів Са2+ до протофібрилвзаємодія з регуляторним білком тропоніномзміна третинної конформації тропоніну та тропоміозину відкриття активних центрів актину взаємодія головок міозину з активними центрами актину (м’язове скорочення). езпосередньо поруч з Т-трубочками розташовані цистерни саркоплазматичного ретикулуму (СПР – частина аграпулярної епдоплазматичної сітки). Т-трубочки з двома поруч розташованими цистернами утворюють тріади. СПР крім цистернмаютьпоздовжні трубочки, в яких локалізуються кальцієві насоси – вони активно транспортують іони – Са2+із саркоплазми в СПР і в його цистернах накопичується велика кількість іонів Са2+. При русі ПД по мембрані Т-трубочок в мембрані цистерн СПР відкриваються кальцієві канали іони Са2+ по градієнту концентрації виходять з цистерн СПР у саркоплазму підвищення концентрації іонів Са2+ в саркоплазмі міоцита з 10–8 до 10–5 ммоль/лдифузія іонів Са2+ до протофібрилвзаємодія з регуляторним білком тропоніномзміна третинної конформації тропоніну та тропоміозину відкриття активних центрів актину взаємодія головок міозину з активними центрами актину (м’язове скорочення).В основі м’язового скорочення лежить ковзання актинових протофібрил відносно міозинових “теорія ковзання”. С  аркомер – сруктурно-функціональний елемент скорочувального апарату скелетних м’язів. Вони утворені пучками міофібрил, які відокремлені один від одного перпендикулярними смугами – Z-лініями. До Z-ліній прикріпляються одним своїм кінцем тонкі актинові нитки. Інші кінці актинових ниток спрямовані до центру саркомера і входять у проміжки між товстими міозиновими нитками. Частина саркомера, яка примикає до Z лінії та утворена тільки актиновими протофібрилами, має назву І-дисків (ізотропних); слідом за ними розташовані А-диски (анізотропні) – частина саркомера, де має місце взаємне перекриття актинових та міозинових протофібрил. При вкороченні м’яза, в, ході його скорочення, коротшає довжина протофібрил вкорочення довжини кожного саркомера. Але при цьому довжина анізотропних дисків не зменшується, а зменшується довжина ізотропних дисків. Це є наслідком ковзання актинових протофібрил відносно міозинових в напрямку до центру саркомера. аркомер – сруктурно-функціональний елемент скорочувального апарату скелетних м’язів. Вони утворені пучками міофібрил, які відокремлені один від одного перпендикулярними смугами – Z-лініями. До Z-ліній прикріпляються одним своїм кінцем тонкі актинові нитки. Інші кінці актинових ниток спрямовані до центру саркомера і входять у проміжки між товстими міозиновими нитками. Частина саркомера, яка примикає до Z лінії та утворена тільки актиновими протофібрилами, має назву І-дисків (ізотропних); слідом за ними розташовані А-диски (анізотропні) – частина саркомера, де має місце взаємне перекриття актинових та міозинових протофібрил. При вкороченні м’яза, в, ході його скорочення, коротшає довжина протофібрил вкорочення довжини кожного саркомера. Але при цьому довжина анізотропних дисків не зменшується, а зменшується довжина ізотропних дисків. Це є наслідком ковзання актинових протофібрил відносно міозинових в напрямку до центру саркомера.П 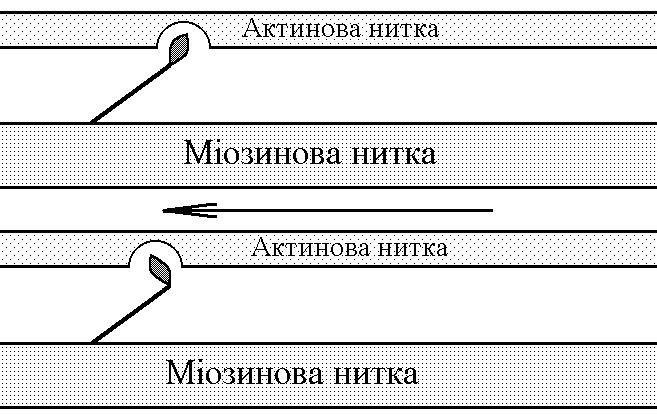 ричиною руху актинових ниток відносно міозинових (їх ковзання) є “нахил” голівки після її приєднання до активного центру актину. Велика кількість голівок, що входять до складу актоміозинових містків, одночасно роблять “нахил” нитка актину переміщується до центру саркомера вздовж нитки міозину. За рахунок одного такого “нахилу” саркомер (м’яз) може вкоротитися на 1% своєї вихідної довжини. Ступінь вкорочення м’яза може досягати 50% такі “нахили” голівок під час одного м’язового скорочення мають повторитися 50 разів. Це можливо, якщо актоміозинові містки після “нахилу” розпадаються (дисоціюють) голівка міозину взаємодіє з наступним активним центром актину наступний “нахил” розпад містку і т.д. На “нахил” однієї головки міозину витрачається енергія однієї молекули АТФ. Для того, щоб відбувся розпад актоміозинового містка необхідно, щоб до голівки міозину приєдналася молекула АТФ (але енергія АТФ при цьому не витрачається). ричиною руху актинових ниток відносно міозинових (їх ковзання) є “нахил” голівки після її приєднання до активного центру актину. Велика кількість голівок, що входять до складу актоміозинових містків, одночасно роблять “нахил” нитка актину переміщується до центру саркомера вздовж нитки міозину. За рахунок одного такого “нахилу” саркомер (м’яз) може вкоротитися на 1% своєї вихідної довжини. Ступінь вкорочення м’яза може досягати 50% такі “нахили” голівок під час одного м’язового скорочення мають повторитися 50 разів. Це можливо, якщо актоміозинові містки після “нахилу” розпадаються (дисоціюють) голівка міозину взаємодіє з наступним активним центром актину наступний “нахил” розпад містку і т.д. На “нахил” однієї головки міозину витрачається енергія однієї молекули АТФ. Для того, щоб відбувся розпад актоміозинового містка необхідно, щоб до голівки міозину приєдналася молекула АТФ (але енергія АТФ при цьому не витрачається).Для розслаблення скелетного м’язу необхідно, щоб зменшилась концентрація іонів Са2+в саркоплазмі. Це досягається завдяки дії кальцієвих насосів повздовжніх трубочок СПР активний транспорт іонів Са2+ в СПР зменшення його концентрації в саркоплазмі до рівня 10–8 ммоль/л від’єднання іонів Са2+ від регуляторного білка тропоніну відновлення вихідної третинної структури тропоніну та тропоміозину блокада активних центрів актину розслаблення м’язу. Таким чином, під час м’язового скорочення енергія АТФ витрачається: 1. На роботу натрій-калієвого насосу зовнішньої мембрани м’язового волокна (цей насос підтримує градієнт концентрації іонів К+ та Na+ поза та всередині клітини, що необхідно для підтримки на необхідному рівні ПС та для розвитку ПД на мембрані). 2. На “нахил” головки міозину. 3. На роботу кальцієвого насосу СПР розслаблення м’язу. 7)Види м’язових скорочень: одинокі та тетанічні; ізотонічні та ізометричні. В залежності від режимів навантаження виділяють наступні види м’язового скорочення. 1 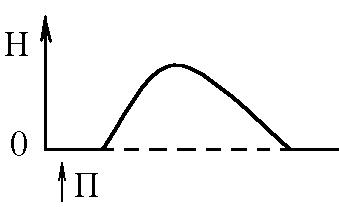 . Ізометричного скорочення – скорочення при незмінній довжинні м’яза.Довжина м’яза при скороченні залишається сталою, але змінюється її напруження (Н). В експерименті таке скорочення можна отримати, якщо ізольований м’яз закріпити з двох сторін та стимулювати електричним струмом. В умовах цілісного організму ізометричне скорочення буває, коли людина намагається, але не може підняти вантаж. . Ізометричного скорочення – скорочення при незмінній довжинні м’яза.Довжина м’яза при скороченні залишається сталою, але змінюється її напруження (Н). В експерименті таке скорочення можна отримати, якщо ізольований м’яз закріпити з двох сторін та стимулювати електричним струмом. В умовах цілісного організму ізометричне скорочення буває, коли людина намагається, але не може підняти вантаж.2 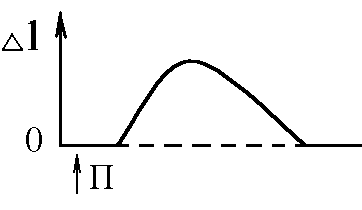 . Ізотонічне скорочення – скорочення при незмінному напруженні м’яза. Напруження м’яза при його скороченні залишається сталим, а довжина змінюється (Δl). В експерименті таке скорочення можна отримати, якщо один кінець ізольованого м’яза закріпити, а інший залишити вільним та стимулювати. В умовах цілісного організму скорочення м’язів максимально наближується до ізотонічного, якщо відбувається згинання ненавантажених кінцівок. . Ізотонічне скорочення – скорочення при незмінному напруженні м’яза. Напруження м’яза при його скороченні залишається сталим, а довжина змінюється (Δl). В експерименті таке скорочення можна отримати, якщо один кінець ізольованого м’яза закріпити, а інший залишити вільним та стимулювати. В умовах цілісного організму скорочення м’язів максимально наближується до ізотонічного, якщо відбувається згинання ненавантажених кінцівок. 3. Змішане (ауксотонічне) скорочення – скорочення, при якому змінюється довжина та напруження м’яза. Такий тип м’язових скорочень найчастіше зустрічається в умовах цілісного організму. В залежності від частоти стимуляції виділяють наступні види м’язового скорочення: 1. Одиночне – спостерігається у відповідь на одиничний стимул чи на їх серію (якщо інтервал між подразниками більший чи рівний тривалості одиночного м’язового скорочення). Виділяють такі періоди м’язового скорочення: - 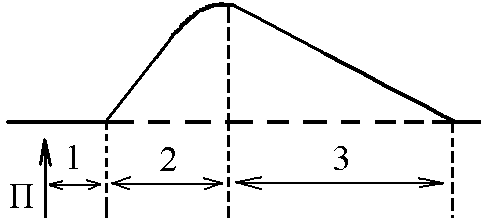 латентий період (1) – інтервал часу від дії подразника (П) до початку скорочення; латентий період (1) – інтервал часу від дії подразника (П) до початку скорочення;- період скорочення (2) – під час цього періоду довжина м’яза зменшується; - період розслаблення (3) – під час цього періоду м’яз розслаблюється, його довжина збільшуєтся до вихідного рівня. 2. Тетанічне – спостерігається в умовах сумації одиночних м’язових скорочень (одне скорочення накладається на інше). Така сумація можлива, якщо м’яз подразнюється серією стимулів і кожен наступний стимул діє тоді, коли попереднє одиночне скорочення ще не закінчилось (тобто, інтервал між стимулами має бути меншим тривалості одиночного м’язового скорочення). Виділяють наступні види тетанічних скорочень: Зубчастий тетанус – виникає на базі одиночних скорочень, якщо кожен наступний стимул попадає на період розслаблення одиночого м’язового скорочення. Отже, інтервал між стимулами в серії має бути менший тривалості одиночного м’язового скорочення, але більший, чим cума тривалості латентного періоду і періоду вкорочення одиночного скорочення. Гладкий тетанус – виникає на базі одиночних скорочень, якщо кожен наступний стимул попадає на латентний період або період вкорочення одиночного м’язового скорочення. Отже, інтервал між стимулами в серії повинний бути меншимчи рівним сумі тривалості латентного періоду одиночного скорочення, але більше тривалості латентного періоду одиночного скорочення. Нижче зображений запис реєстрації скорочень м’яза при збільшенні частоти її стимуляції. Видно, що при збільшенні частоти стимуляції на фазі одиночних скорочень виникають тетанічні, спочатку зубчастий, а потім гладкий тетанус. Тетанічні скорочення відрізняється від одиночного наступними параметрами: 1. Тривалість тетанічного скорочення більша, ніж одиночного. 2. Амплітуда (сила) тетанічного скорочення більша, ніж одиночного. Амплітуда гладкого тетануса більша, ніж зубчастого. 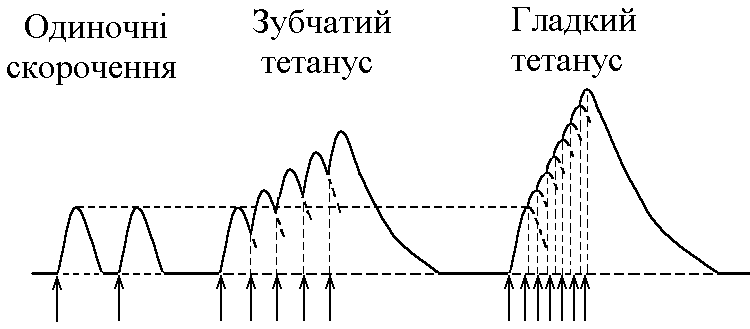 Стрілочками показано момент дії подразника на м’язове волокно. Асинхронний тетанус. Асинхронний тетанус представляє вид скорочень, який притаманний скелетним м’язам при їх природній діяльності в умовах цілісного організму. Кожен скелетний м’яз складається з великої кількості окремих м’язових волокон. При природній діяльності м’язів в умовах цілісного організму частота, з яким до них надходять ПД по аксонах мотонейронов недостатня для того, щоб у кожному окремому волокні м’яза розвивався гладкий тетанус. У той же час м’язи людини здатні до тривалого злитого скорочення. Таке скорочення є наслідком асинхронного (неодночасного) скорочення окремих м’язових волокон м’яза. Кожне волокно скорочується в режимі одиночних скорочень, але в межах м’яза відбувається (додавання) скорочень окремих волокон. У підсумку м’яз скорочується злитно та довготривало. Це і є асинхронний тетанус. Його тривалість може бути дуже великою, а його амплітуда не буває такою значною, як амплітуда гладкого тетанусу, який можна одержати в експерименті. 8)Центральне гальмування розподіляється за локалізацією на пре- та постсинаптичне; за характером поляризації (зарядом мембрани) - на гіпер- та деполяризаційне; за будовою гальмівних нейронних ланцюгів - на реципрокне, або поєднане, зворотне та латеральне. Розрізняють дві форми гальмування: 1.Первинне - для його виникнення необхідна наявність спеціальних гальмівних структур (гальмівних нейронів і гальмівних синапсів). Гальмування в даному випадку виникає без попереднього збудження. Може бути двох видів: а) Пресинаптичне гальмування, як свідчить назва, локалізується в пресинаптичних елементах і пов'язане з пригніченням проведення нервових імпульсів у аксональних (пресинаптичних) закінченнях. У основі такого гальмування лежить розвиток тривалої деполяризации терміналі аксона і, як наслідок, блокування проведення збудження. Воно особливо ефективне для обробки інформації, яка надходить до нейрона різними пресинаптичними шляхами, оскільки в цьому разі збудження, що надходить через один із входів, може бути вибірково зменшене або навіть повністю пригнічене, якщо немає інших впливів на інші синаптичні входи. Припускають, що пресинаптичне гальмування пригнічує сигнали, біологічне значення яких у даний момент є незначним, і пропускає інформацію, що є найважливішою для організму. Механізм: До збудливого аксону підходить вставний гальмівний аксон, який виділяє гальмівний медіатор ГАМК. Цей медіатор діє на постсинаптичну мембрану, яка є мембраною збудливого аксона, і викликає в ній деполяризацію. Ця деполяризація гальмує вхід Са2+ з синаптичної щілини в закінчення збудливого аксона і таким чином призводить до зниження виділення збудливого медіатора в синаптичну щілину гальмуючи реакцію. Пресинаптичне гальмування досягає максимуму через 15-20 мс і триває близько 150 мс, тобто набагато довше, ніж постсинаптичні гальмування. Пресинаптичне гальмування блокується судомними отрутами - бікуліном і пікротоксином, які є конкурентними антагоністами ГАМК. б) Постсинаптичне гальмування (ГПСП) обумовлене виділенням пресинаптичним закінченням аксона гальмівного медіатора (гамма-аміномасляна кислота (ГАМК) чи гліцин), який знижує або гальмує збудливість мембран соми і дендритів нервової клітини, з якою він контактує. Гальмівні медіатори виділяються спеціальними гальмовими нейронами - клітинами Реншоу (в спинному мозку) і корзинчатими клітинами(в проміжному мозку), зірчасті клітини кори великого мозку та ін . Клітини Реншоу забезпечують розвиток гальмування в мотонейронах м'язів - антагоністів. Вони також забезпечують ще одне (антидромное) гальмування, оберігаючи мотонейрони від перезбудження. Корзинчаті клітинирегулюють потоки імпульсів збудення, що йдуть до центрів проміжного мозку і кори півкуль. Вони викликають синхронне гальмування цілої групи нейронів діэнцефальных центрів, регулюючи в такий спосіб ритм активності кори. Постсинаптичне гальмування локальне. Розвивається воно градуально, здатне до сумації, не залишає після себе рефрактерності. Є більш оперативним, чітко адресованим та універсальним гальмівним механізмом. За своєю суттю це «центральне гальмування», яке було описане у свій час Ch. S. Sherrington (1906). Дослідженням П. Г. Костюка ( 1977) довело, що постсинаптичне гальмування пов'язане з первинною гіперполяризацією мембрани соми нейрона, в основі якої лежить підвищення проникності постсинаптичної мембрани для К+. Внаслідок гіперполяризації рівень мембранного потенціалу віддаляється від критичного (порогового) рівня. Тобто відбувається його збільшення - гіперполяризація . Це призводить до гальмування нейрона. Такий вид гальмування називаєтьсягіперполярізаційним. Амплітуда і полярність ГПСП залежать від вихідного рівня мембранного потенціалу самого нейрона. Механізм цього явища пов'язаний з Сl+. З початком розвитку ГПСП Сl- входить в клітину. Коли в клітині стає його більше, ніж зовні, гліцин конформує мембрану і через її відкриті отвори Сl- виходить з клітини. У ній зменшується кількість негативних зарядів, розвивається деполяризація. Такий вид гальмування називається деполярізаційним. Постсинаптичне гальмування є найпоширенішим у ЦНС. В основі цього виду гальмування лежить гальмівний постсинаптичний тенціал (ГПСП), який є локальною мембрани, що виникає внаслідок підвищення її проникності до СІ- або К+. Залежно від структури гальмівного нейронного ланцюжка, розрізняють такі форми постсинаптичного гальмування: реципрокне, зворотнє та латеральне. Пряме постсинаптичне гальмування виникає у постсинаптичному нейроні внаслідок гіперполяризації його мембрани під час розвитку ГПСП під впливом ПД будь-якого гальмівного нейрона. Зворотне постсинаптичне гальмування здійснюється у нейронному ланцюзі, який складається з рухового нейрона та вставного гальмівного нейрона (клітини Реншоу). Імпульси від збудженого рухового нейрона «тільки прямують через аксон, а й поширюються через його колатералі, що відгалужуються від нього, і активують вставний нейрон — клітину Ренто. Цей гальмівний нейрон спричинює пригнічення розрядів рухового нейрона. Отже, з цих двох нейронів утворюється контур з негативним зворотним зв'язком, який дає змогу стабілізувати частоту розрядів рухового нейрона, обмежуючи надмірну імпульсацію до ефекторного органа. Латеральне гальмування. Гальмівний ланцюг нейронів характеризується тим, що вставні гальмівні нейрони впливають не тільки на збуджену клітину, але й на сусідні нейрони, в яких збудження є слабшим або зовсім відсутнє. Таке гальмування називається латеральним, позаяк ділянка гальмування, яка утворюється, міститься збоку (латерально) від збудженого нейрона. Реципрокне (поєднане) гальмування характеризується тим, що у тому випадку, коли при активації аферентів збуджуються, наприклад, мотонейрони м'язів-згиначів, то одночасно (на цьому боці) гальмуються мотонейрони м'язів-розгиначів, які діють на цей же суглоб. Відбувається це тому, що аференти від м'язових веретен утворюють збуджуючі синапси на мотонейронах м'язів–агоністів, а через посередництво вставного гальмівного нейрона - гальмівні синапси на мотонейронах м'язів-антагоністів. З фізіологічної точки зору таке гальмування дуже вигідне, оскільки полегшує рух суглоба (працює «автоматично», без додаткового довільного або мимовільного контролю). Постсинаптичне гальмування переважно легко знімається при введенні стрихніну, який конкурує з гальмівним медіатором (гліцином) на постсинаптичній мембрані. Правцевий токсин також пригнічує постсинаптичне гальмування, порушуючи вивільнення медіатора з гальмівних пресинаптичних закінчень. Тому введення стрихніну або правцевого токсину супроводжується судомою, яка виникає внаслідок різкого посилення процесу збудження в ЦНС, зокрема, мотонейронів. 2. Вторинне- не потребує наявності гальмівних структур. Гальмування розвивається в результаті змін функціональної активності збудливих нейронів. |