Модуль 1 1 Потенціал спокою
 Скачать 1.25 Mb. Скачать 1.25 Mb.
|

|
11) Рухові рефлекси середнього мозку, їх фізіологічне значення. Середній мозок (СрМ) за участі сітчастої речовини опрацьовує аферентну інформацію, яка поступає в спинний та задній мозок. Нова інформація поступає в СрМ від зорових та слухових рецепторів. Її опрацювання відбувається в ядрах передніх (від зорових рецепторів) та нижніх (від слухових рецепторів) горбків. На основі опрацьовання інформації від усіх цих рецепторів СрМ здійснює контроль за станом зовнішнього та внутрішнього середовища організма. Важливими надсегментарними руховими ядрами СрМ є: 1) червоні ядра – від них інформація від нейронів спинного мозку передається по шляхах що перехрещуються (руброспінальні шляхи – елемент ЛНС); 2) ретикулярна формація; 3) чорна субстанція – знаходиться в середньому мозку, але функціонує як єдине ціле з базальними ядрами. Руброспінальні впливи характеризуються активацією - та -мотонейронів флексорів та гальмуванням - та -мотонейронів екстензорів. Ці впливи протилежні вестибулоспінальним. Завдяки руброспінальним впливам тонус м’язів мезенцефальної тварини більш рівномірніший, ніж у децереброваної (тонус флексорів та екстензорів більш збалансований). Тому, поза мезенцефальної тварини нормальна. Тобто, посилена антигравітаційна поза децереброваної тварини перетворюється в нормальну у мезенцефальної завдяки руброспінальним впливам. За участю рухових ядер СрМ здійснюються позо-тонічні рефлекси двох видів: 1.Випрямні – забезпечують відновлення порушеної пози. У тварин, що мають шию, випрямні рефлекси мають 2 фази: а) Рефлекс випрямлення голови – він починається з подразнення отолітових вестибулорецепторів у відповідь на порушення нормального положення голови в просторі (наприклад при падінні). Рівень замикання цього рефлексу – СрМ. Для його здійснення необхідна участь червоних ядер та РФ СрМ. По низхідних шляхах від цих структур інформація передається до мотонейронів спинного мозку перерозподіл тонусу м’язів шиї голова набуває нормальне положення в просторі (тім’ячком доверху). б) Рефлекс випрямлення тулуба – починається з подразнення пропріорецепторів шиї, що спричиняється рефлексом випрямлення голови, так як при цьому відновлюється нормальне положення голови у просторі, але порушується її нормальне положення відносно тулуба. Вхід інформації від пропріорецепторів шиї здійснюється на рівні спинного мозку підйом до надсегментарних структур СрМ обробка цієї інформації передача до мотонейронів спинного мозку до м’язів, які забезпечують відновлення нормального положення тулуба відносно голови у просторі. 2. Стато-кінетичні – забезпечують збереження пози рівноваги при русі з прискоренням. При русі з лінійним прискоренням збуджуються отолітові рецептори, а при русі з кутовим прискоренням – рецептори півколових канальців. Вхід цієї інформації в ЦНС здійснюється на рівні заднього мозку підйом до структур СрМ обробка та передача низхідними шляхами до мотонейронів спинного мозку перерозподіл тонусу м’язів, який забезпечує збереження рівноваги під час руху з прискоренням. Важливим компонентом стато-кінетичних рефлексів є присінково-окорухові рефлекси ністагм очей більш продовжена фіксація предметів в полі зору під час руху краща орієнтація в просторі більш надійне збереження пози при русі. Ще раз підкреслимо, що рівень замикання випрямних та стато-кінетичних рефлексів – СрМ. Тому, їх можливо отримати у мезенцефальної тварини і неможливо у децереброваної чи спінальної. За рахунок обробки інформації від зорових та слухових рецепторів (за участю ядер чотирьох горбків) рухові ядра СрМ забезпечують здійснення орієнтаційних рефлексів – у відповідь на сильні звукові та світлові подразники мезенцефальна тварина повертає голову в бік подразника, насторожує вуха, в неї дещо посилюється тонус флексорів – тварина ніби готується до відповіді на цей подразник. Роль СрМ в здійсненні фазичних реакцій залежить від рівня організації тварини: - мезенцефальні земноводні (жаба) здатні до локомоцій – руху у просторі (стрибають, плавають); - кішки та собаки не здатні до локомоцій, але здійснюють локомоторні синергії – злагоджені рухи кінцівок, що нагадують акт ходьби: їх інтегрують структури СрМ разом з структурами заднього та спинного мозку. У людини СрМ не забезпечує формування нормальної антигравітаційної пози в спокої та підтримання її при русі. Для здійснення всіх цих функцій у людини необхідна участь кори великих півкуль (принцип кортикалізації функцій). 12)Мозочок, його функції, симптоми ураження. Мозочок (Мз) відіграє дуже важливу роль в координації рухів і виконує цю функцію на основі своїх аферентних та еферентних зв’язків. По аферентним волокнам сигнали поступають в кору мозочка, яка має складну будову та забезпечує тонку обробку поступившої інформації. Причому, кінцеву обробку інформації здійснюють грушоподібні клітини Пуркіньє – по відростках цих клітин (по їх аксонах) здійснюється еферентний вихід обробленої інформації з кори Мз. Далі, інформація здебільшого прямує до ядер Мз по аксонах їх нейронів здійснюється вихід інформації за межі Мз. Аферентні зв’язки Мз: 1. Від рецепторів шкіри та пропріорецепторів Мз отримує інформацію по спіно-церебелярних шляхах; ця інформація забезпечує можливість контролю за рухом який здійснюється. 2. Від вестибулорецепторів (через вестибулярні ядра) – контроль за збереженням рівноваги при русі. 3. Від всіх рухових ядер стовбуру (ретикулярна формація, краєві ядра). 4. Від базальних ядер. 5. Від моторної і асоціативної кори (від моторної кори надходить копія команди до виконання руху, яка відправляється до мотонейронів; від асоціативної кори – інформація про ціль руху). 6. Від зорових та слухових рецепторів – контроль за рухами, які відбуваються. Еферентні зв’язки Мз: 1. З руховими ядрами стовбуру (ретикулярна формація, вестибулярні ядра, червоні ядра), через які Мз здійснює вплив на мотонейрони і на м’язи. 2. З базальними ядрами. 3. З асоціативною та моторною корою 4. З гіпоталамусом. Таким чином, зв’язки Мз обширні і нерідко є двосторонніми (з корою, з базальними ядрами, з руховими ядрами стовбуру). Мз отримує інформацію про характер руху від рецепторів і при необхідності може швидко корегувати ці рухи та їх програму. При цьому різні відділи Мз здійснюють корекцію різних рухів. Др 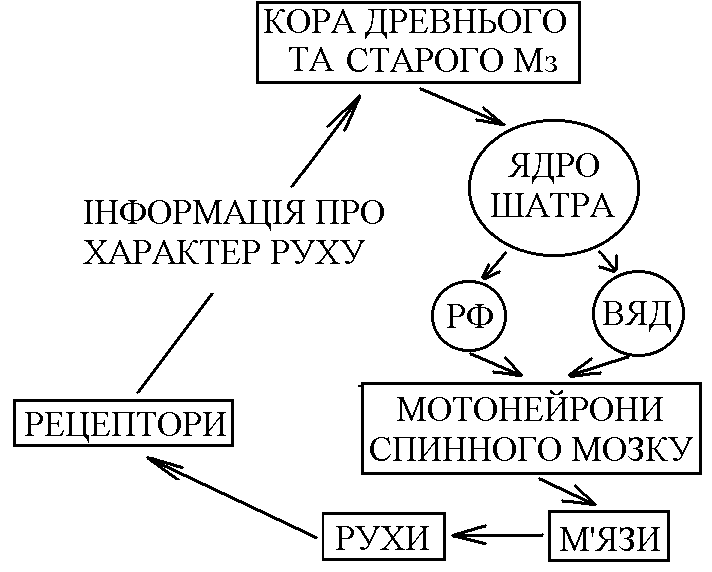 евній та старий Мз (флокуло-нодулярна система та хробак) – отримують інформацію від вестибулорецепторів, пропріорецепторів та шкірних рецепторів. Кора цих відділів передає інформацію до ядра шатра і далі до вестибулярного ядра до медіального ретикулярного ядра до мотонейронів, що іннервують м’язи тулуба та кінцівок. евній та старий Мз (флокуло-нодулярна система та хробак) – отримують інформацію від вестибулорецепторів, пропріорецепторів та шкірних рецепторів. Кора цих відділів передає інформацію до ядра шатра і далі до вестибулярного ядра до медіального ретикулярного ядра до мотонейронів, що іннервують м’язи тулуба та кінцівок.Таким чином, кора древнього та старого Мз, отримуючи інформацію від пропріорецепторів, рецепторів шкіри про характер руху; від вестибулорецепторів – про збереження рівноваги, і при необхідності вносить корективи в грубі позо-тонічні рухи, чому передує передача інформації по шляхах МНС, які регулюють стан м’язів тулуба і проксимальних відділів кінцівок. Ця корекція дуже важлива для підтримання рівноваги (збереження пози) при русі. Пр 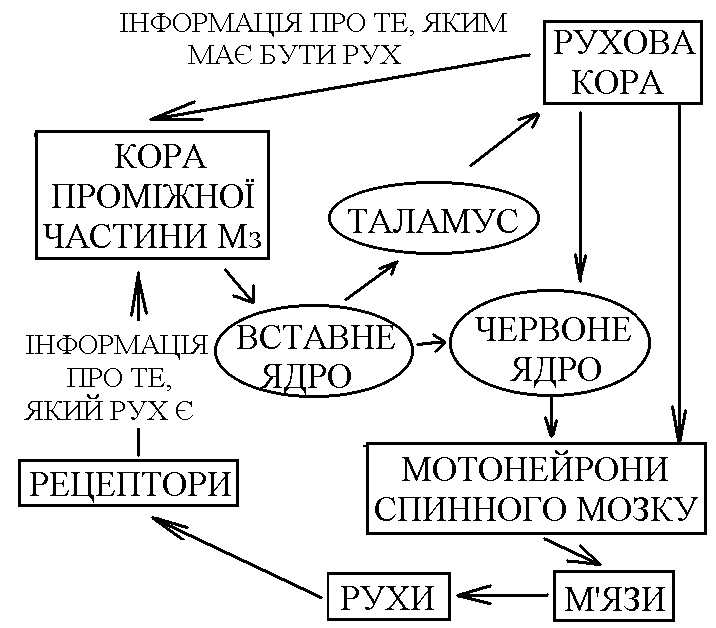 оміжна частина кори Мз, як і кора древнього та старого Мз, отримує інформацію про те, який рух здійснюється. Від рухової кори сюди поступає інформація про те, який рух повинен відбуватися (копія рухової програми). Кора проміжної частини Мз працює як блок порівняння – порівнює інформацію про те, який рух повинен відбуватися, і про те, який рух проходить. При неспівпаданні цієї інформації Мз може вносити такі корективи: оміжна частина кори Мз, як і кора древнього та старого Мз, отримує інформацію про те, який рух здійснюється. Від рухової кори сюди поступає інформація про те, який рух повинен відбуватися (копія рухової програми). Кора проміжної частини Мз працює як блок порівняння – порівнює інформацію про те, який рух повинен відбуватися, і про те, який рух проходить. При неспівпаданні цієї інформації Мз може вносити такі корективи:а) в рух, який здійснюється (виходячи на мотонейрони спинного мозку через стовбурові рухові ядра); б) в рухову програму, передаючи інформацію по церебело-таламо-кортикальному шляху в рухову кору. Вихід до рухової кори і до стовбурових рухових ядер із кори проміжної частини Мз здійснюється через вставне ядро. За рахунок корекції рухів та програми Мз добивається їх повного співпадання (їх відповідності один одному). Кора півкуль Мз пов’язана з базальними гангліями і з асоціативною корою. Завдяки цим зв’язкам вони формують програми рухів і Мз через зубчасте ядропередає їх в рухову кору для виконання. Основні симптоми ураження Мз у людини. Мехпнізми їх виникнення. 1. Дистонія – порушення тонусу м’язів, що спричиняється порушенням впливу Мз на стовбурові рухові ядра (ретикулярна формація, вестибулярні ядра) на мотонейрони спинного мозку на м’язи тулуба та проксимальних відділів кінцівок. 2. Дизеквілібрія – перушення рівноваги. Пов’язана з випаданням коригуючих впливів Мз на вестибулярні ядра Дейтерса. 3. Атаксія – невпорядкованість рухів. Проявляється у вигляді: а) дисметрія – порушення амплітуди рухів; б) порушення направленості рухів. В основі атаксії лежать порушення координації м’язів агоністів та антагоністів. 4. Асинергія – порушення злагодженості рухів. В основі її лежить порушення координації скорочення м’язів-агоністів. 5. Астенія – швидка втомлюваність. Вона пов’язана з виконанням великої кількості “зайвих” рухів для досягнення мети. 6. Адіадохокінез – порушення синхронного виконання рухів, іх послідовності та швидкості. Дисметрія – порушення рівномірності і амплітуди рухів. Дисартрія – порушення артикуляції мови. 13)Синапс – это морфофункциональное образование ЦНС, которое обеспечивает передачу сигнала с нейрона на другой нейрон или с нейрона на эффекторную клетку (мышечное волокно, секреторную клетку). Все синапсы ЦНС можно классифицировать следующим образом. 1. По локализации: центральные (головной и спинной мозг) и периферические (нервно-мышечный, нейросекреторный синапс вегетативной нервной системы). Центральные синапсы можно в свою очередь разделить на аксо-аксональные, аксо-дендритические (дендритные), аксо-соматические, дендро-дендритические, дендро-соматические и т.п. Согласно Г. Шенсрду, различают реципрокные синапсы, последовательные синапсы и синаптические гломерулы (различным способом соединенные через синапсы клетки). 2. По развитию в онтогенезе: стабильные (например, синапсы дуг безусловного рефлекса) и динамичные, появляющиеся в процессе индивидуального развития. 3. По конечному эффекту: тормозные и возбуждающие. 4. По механизму передачи сигнала: электрические, химические, смешанные. 5. Химические синапсы можно классифицировать: а) по форме контакта – терминальные (колбообразное соединение) и преходящие (варикозное расширение аксона); б) по природе медиатора – холинергические (медиатор – ацетилхолин, АХ), адренергическис (медиатор – норадреналин, НА), дофаминергические (дофамин), ГАМК-ергические (медиатор – гаммааминомасляная кислота), глицинергические, глутаматергические, аспартатсргические, пептидергические (медиатор – пептиды, например, вещество Р), пуринергические (медиатор – АТФ). Электрические синапсы. В настоящее время признают, что в ЦНС имеются электрические синапсы. С точки зрения морфологии электрический синапс представляет собой щелевидное образование (размеры щели до 2 нм) с ионными мостиками-каналами между двумя контактирующими клетками. Петли тока, в частности при наличии потенциала действия (ПД), почти беспрепятственно перескакивают через такой щелевидный контакт и возбуждают, т.е. индуцируют генерацию ПД второй клетки. В целом, такие синапсы (они называются эфапсами) обеспечивают очень быструю передачу возбуждения. Но в то же время с помощью этих синапсов нельзя обеспечить одностороннее проведение, т. к. большая часть таких синапсов обладает двусторонней проводимостью. Кроме того, с их помощью нельзя заставить эффекторную клетку (клетку, которая управляется через данный синапс) тормозить свою активность. Аналогом электрического синапса в гладких мышцах и в сердечной мышце являются щелевые контакты типа нексуса. Химические синапсы. По строению химические синапсы представляют собой окончания аксона (терминальные синапсы) или его варикозную часть (проходящие синапсы), которая заполнена химическим веществом – медиатором. В синапсе различают пресинаптический элемент, который ограничен пресинаптической мембраной, постсинаптический элемент, который ограничен постсипаптической мембраной, а также внесинаптическую область и синаптическую щель, величина которой составляет в среднем 50 нм. В литературе существует большое разнообразие в названиях синапсов. Например, синаптическая бляшка – это синапс между нейронами, концевая пластинка – это постсинаптическая мембрана мионеврального синапса, моторная бляшка – это пресинаптичсское окончание аксона на мышечном волокне. Общие принципы работы химического синапса Любой химический синапс, независимо от природы медиатора и хеморецептора, активируется под влиянием потенциала действия, прибегающего к пресинапсу от тела нейрона. В результате – происходит деполяризация пресинаптической мембраны, что повышает проницаемость кальциевых каналов пресинаптической мембраны и приводит к увеличению входа в пресинапс ионов кальция. В ответ на это происходит высвобождение квантов (выход из пресинапса) – 100–200 порций (квантов) медиатора. Выйдя в синаптическую щель, медиатор взаимодействует со специфическим рецептором постсинаптической мембраны, что вызывает изменение ионной проницаемости. В синапсах, в которых осуществляется возбуждение постсинантической структуры, обычно происходит повышение проницаемости для ионов натрия, что вызывает деполяризацию постсинаптической мембраны. Эта деполяризация получила определенное название: возбуждающий постсинаптический потенциал (ВПСП). Если его величина достаточно велика и достигает критического уровня деполяризации, то генерируется ПД. В тормозных синапсах в результате взаимодействия медиатора с рецепторами, наоборот, происходит гиперполяризация (за счет, например, увеличения проницаемости для ионов калия и хлора). Это называется тормозным постсинаптическим потенциалом (ТПСГ1). В гиперполяризоваином состоянии клетка снижает свою возбудимость и благодаря этому прекращает отвечать на внешние раздражители или (если она обладала свойством автоматии) уменьшает спонтанную активность. После каждого цикла проведения импульса медиатор разрушается, например, ацетилхолин разрушается ацстилхолинэстеразой, норадреналин разрушается моноаминоксидазой (МАО) или катсхол-0-метилтрансферазой (КОМТ), либо идет обратный захват в преси-наптическую структуру. В одних случаях захватывается неразрушенный медиатор (например, норадреналип), в других – его осколки (например, холин из ацетилхолина). Синтез медиатора совершается в пресинаптическом элементе, куда попадают исходные продукты для синтеза и ферменты, необходимые в синтезе. Ферменты образуются в соме нейрона и по аксону, примерно со скоростью 6 мм/сутки, попадают в пресинапс, где используются в процессе синтеза медиатора. Угнетение активности этих ферментов фармакологическим путем может привести к истощению запасов медиатора в синапсе и, следовательно, к снижению его функциональной способности. Свойства химических синапсов 1. Односторонняя проводимость – одно из важнейших свойств химического синапса. Асимметрия – морфологическая и функциональная – является предпосылкой для существования односторонней проводимости. 2. Наличие синаптической задержки: для того, чтобы в ответ на генерацию ПД в области пресинапса выделился медиатор и произошло изменение постсинаптического потенциала (ВИСИ или ТПСП), требуется определенное время (синаптическая задержка). В среднем оно равно 0,2–0,5 мс. 3. Благодаря синаптическому процессу нервная клетка, управляющая данным постсинаптичсским элементом (эффектором), может оказывать возбуждающее воздействие или, наоборот, тормозное (это определяется конкретным синапсом). 4. В синапсах существует явление отрицательной обратной связи – антидромный эффект. Речь идет о том, что выделяемый в синаптическую щель медиатор может регулировать выделение следующей порции медиатора из этого же пресинаптического элемента путем воздействия на специфические рецепторы пресинаптичсской мембраны. 5. Эффективность передачи в синапсе зависит от интервала следования сигналов через синапс. Если этот интервал до некоторых пор уменьшать (учащать подачу импульса по аксону), то на каждый последующий ПД ответ постсинаитической мембраны (величина ВПСП или ТПСП) будет возрастать (до некоторого предела). Это явление облегчает передачу в синапсе, усиливает ответ постсинаптического элемента (объекта управления) на очередной раздражитель; оно получило название «облегчение» или «потенциация». В основе его лежит накопление кальция внутри пресинапса. Если частота следования сигнала через синапс очень большая, то из-за того, что медиатор не успевает разрушиться или удалиться из синаптической щели, возникает стойкая деполяризация или катодическая депрессия – снижение эффективности синаптической передачи. Это явление называется депрессией. Если через синапс проходит много импульсов, то в конечном итоге постсинаптическая мембрана может уменьшить ответ на выделение очередной порции медиатора. Это называется явлением дссепситизации – утратой чувствительности. В определенной мере десеиситизация похожа па процесс рефрактерности (утрата возбудимости). Синапсы подвержены процессу утомления. Возможно, что в основе утомления (временного падения функциональных возможностей синапса) лежат: а) истощение запасов медиатора, б) затруднение выделения медиатора, в) явление дссенситизации. Т. о., утомление – это интегральный показатель. В мозге имеется ряд медиаторов, вызывающих возбуждение нейрона: норадреналин (его продуцируют адренергические нейроны), дофамин (дофаминергические нейроны), серото-нин, пептиды (пептидергические), глутаминовая кислота, аспарагиновая кислота и т.д. Во всех этих случаях выделяющийся медиатор взаимодействует со специфическим рецептором, в результате чего меняется проницаемость для ионов натрия, калия или хлора, и в итоге развивается деполяризация (ВПСП). Если она достигает критического уровня деполяризации, то возникает ПД (возбуждение нейрона). Тормозные синапсы образованы специальными тормозными нейронами (точнее, их аксонами). Медиатором могут быть глицин, гамма-аминомасляная кислота (ГАМК) и ряд других веществ. Обычно глицин вырабатывается в синапсах, с помощью которых осуществляется постсинаптическое торможение. При взаимодействии глицина как медиатора с глициновыми рецепторами нейрона возникает гиперполяризация нейрона (ТПСП) и, как следствие, – снижение возбудимости нейрона вплоть до полной его рефрактсрности. В результате этого возбуждающие воздействия, оказываемые через другие аксоны, становятся малоэффективными или неэффективными. Нейрон выключается из работы полностью. |