Модуль 1 1 Потенціал спокою
 Скачать 1.25 Mb. Скачать 1.25 Mb.
|

|
Види вторинного гальмування: а) Позамежне гальмування - виникає в нейронах центральної нервової системи в тому випадку , якщо потік інформації до тіла нейрона вище його працездатності. Розвивається різке зниження збудливості нейрона. б) Парабіотичне гальмування - виникає при дії сильних і довготривалих подразників (парабіоз в тканинах). Парабіоз - явище межового стану між загибеллю і життям тканини (різко падають всі властивості тканини, основна властивість - фазна зміна лабільності). Якщо парабіотичний фактор продовжує діяти, тканина гине . в) Песимальне гальмування (гальмування Введенського) (лат. - найгірший) - виникає внаслідок зниження функціональної лабільності збудливої структури в разі надмірних за силою або частих подразнень і виявляється пригнічена рефлекторної діяльності. Песимальне гальмування відіграє важливу роль у механізм координації рефлекторної діяльності в ЦНС. Воно не пов'язане з активацією спеціалізованих гальмівних утворів, а виникає внаслідок поступового зниження збудливості, і ті самі імпульси, що раніше викликали рефлекторний акт, тепер його гальмують. Значення процесів гальмування у функціонуванні нервової системи надзвичайно велике: воно обмежує неконтрольоване поширення збудження у ЦНС, забезпечуючи чіткі координовані реакції; відіграє охоронну роль, захищаючи нейрони від ушкодження внаслідок надмірних за силою подразнень, значний внесок у вивчення механізмів центрального гальмування зробили праці таких вчених, як І. М. Сєченов, Ч. Шеррінгтон, Є. Введенський, Дж. Екклс, П. Г. Костюк, П. М. Сєрков. г) Гальмування вслід за збудженням (А. А. Ухтомський) - пригнічення нейронів після порушення. Є результатом того, що слідом за піком потенціалу дії виникає період слідової гіперполяризації, який характеризується зниженням збудливості. Після сильних ритмічних подразнень лапки жаби зникнення подразника призводить до миттєвого розслаблення лапки-піднята кінцівка падає, як батіг. Описана послідовність процесів збудження і гальмування часто зустрічаються в корі великих півкуль при умовно-рефлекторній діяльності. д) Гальмування за принципом негативної індукції. е) Умовне (внутрішнє ) гальмування. 9)Рухові рефлекси спинного мозку, їх рефлекторні дуги, фізіологічне значення Спинний мозок – перший рівень ЦНС. Він має сегментарну будову – кожен сегмент отримує аферентну інервацію від відповідного сегменту тіла людини і до цього сегменту передає еферентну інформацію (еферентно інервує м'язи відповідного сегменту). Аферентна інформація надходить в кожен сегмент волокнами задніх корінців (відростки псевдоуніполярних клітин), еферентна інформація виходить передніми корінцями (відростки мотонейронів передніх рогів та вегетативних ядер бічних рогів). У складі задніх рогів спинного мозку переважають вставні нейрони. Біла речовина спинного мозку представлена волокнами висхідних та низхідних шляхів. Контроль на рівні спинного мозку Рецептори шкіри Вісцерорецептори, ангіорецептори. Вид рецепторів Пропріорецептори (м'язів, сухожилків, суглобів) Про зміну у зовнішньому середовищі Про що передається інформація Про зміну у внутрішньому середовищі Про ступінь розтягнення, скорочення м'яза, про зміну положення кінцівок Контроль на рівні спинного мозку. Вид рецепторів рецептори шкіри На основі аналізу аферентної інформації від пропріорецепторів, рецепторів шкіри, вісцерорецепторів спинний мозок забезпечує регуляцію рухових функцій (тонічних та фазичних). Р  оль спинного мозку у здійсненні тонічних рефлексів можна довестиу досліді з високою перерізкою сідничного нерва у спінальної жаби (у такої жаби зруйновані всі рівні ЦНС окрім спинного мозку). Якщо таку жабу підвісити на штативі, кінцівка зі сторони перерізки провисає і ніби подовжується за рахунок зменшення ступеня згинання в колінному, гомілковому суглобах. Зміна положення кінцівки зі сторони перерізки пов'язані зі зникненням після перерізки м'язового тонусу. Цей дослід доводить: оль спинного мозку у здійсненні тонічних рефлексів можна довестиу досліді з високою перерізкою сідничного нерва у спінальної жаби (у такої жаби зруйновані всі рівні ЦНС окрім спинного мозку). Якщо таку жабу підвісити на штативі, кінцівка зі сторони перерізки провисає і ніби подовжується за рахунок зменшення ступеня згинання в колінному, гомілковому суглобах. Зміна положення кінцівки зі сторони перерізки пов'язані зі зникненням після перерізки м'язового тонусу. Цей дослід доводить:а) Наявність м'язового тонусу у спінальної жаби; б) Рефлекторну природу цього тонусу, оскільки він щезає після перерізки нерва, тобто після пошкодження рефлекторної дуги. В 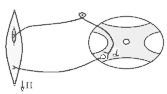 основі формування спінального тонусу лежить міотатичний тонічний рефлекс. Схема дуги цього рефлексу наведена зліва. основі формування спінального тонусу лежить міотатичний тонічний рефлекс. Схема дуги цього рефлексу наведена зліва.У спінальної жаби, підвішаної на штативі, м'язи-згиначі розтягуються силою тяжіння. При цьому збуджуються м'язові веретена (адекватний подразник рецептора – розтягнення центральної частини інтрафузального веретена) моносинаптично активуються -мотонейрони, що іннервують екстрафузальні волокна цього ж м'яза. При перерізці сідничного нерва порушуються провідникові ланки рефлекторної дуги тонус зникає кінцівка розгинається. Т О 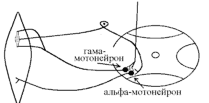 тже, тонус спінальної тварини слабкий, він недостатній для створення антигравітаційної пози. Причиною слабкого тонусу у спінальної тварини є відсутність активуючих впливів на - і на -мотонейрони спинного мозку, надсегментарних утворень вище лежачих рівнів ЦНС (заднього, середнього мозку і т.д.). Активація -мотонейронів призводить до посилення скорочення екстрафузальних волокон м'яза посилення його тонусу. тже, тонус спінальної тварини слабкий, він недостатній для створення антигравітаційної пози. Причиною слабкого тонусу у спінальної тварини є відсутність активуючих впливів на - і на -мотонейрони спинного мозку, надсегментарних утворень вище лежачих рівнів ЦНС (заднього, середнього мозку і т.д.). Активація -мотонейронів призводить до посилення скорочення екстрафузальних волокон м'яза посилення його тонусу.Активація -мотонейронів скорочення скоротливих сегментів інтрафузальних волокон розтяг центральної частини рецептора збудження нервового закінчення передача інформації до -мотонейронів до екстрафузальних волокон м'яза їх скорочення посилення тонусу м'язів. Тобто, в даному випадку посилення тонусу пов'язане з передачею інформації до -мотонейронів по -петлі. Роль спинного мозку в здійсненні фазичних реакцій можна простежити на спінальній жабі при нанесенні їй на шкіру механічних та хімічних подразнень. При цьому можна отримати більшу кількість рефлекторних фазичних реакцій (згинальний, почухувальний, крокувальний і т.д.). Та всі ці реакції однотипні – зводяться до згинання та розгинання кінцівок та тулуба тварини. Рефлекси можуть бути сегментарними (замикатися на рівні одного сегмента локальний рефлекс) і міжсегментарними (нервові сигнали іррадіюють в інші сегменти спинного мозку генералізована рухова реакція). Р 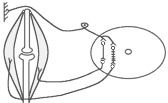 ефлекторна дуга спінального шкірно-м'язового рефлексу (захисний згинальний рефлекс). ефлекторна дуга спінального шкірно-м'язового рефлексу (захисний згинальний рефлекс).В клінічній практиці окрім шкірно-м'язового рефлексу велике значення мають сухожильні рефлекси. Свою назву він отримав у зв'язку з тим, що його можна викликати нанесенням короткого удару неврологічним молоточком по сухожилку м'яза. Наприклад, при дослідженні колінного рефлексу наносять удар по суходилку чотирьохголового м'яза стегна. У відповідь на це кінцівка в колінному суглобі спочатку розгинається (перша фаза рефлексу), а потім згинається (друга фаза). М 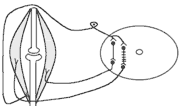 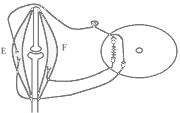 еханізм розвитку першої фази рефлексу заключається в наступному: при ударі по сухожилку м'яз швидко розтягується збудження м'язових веретен моносинаптична активація -мотонейронів, що інервують чотирьохголовий м'яз та гальмування (за участю вставних гальмівних нейронів) центрів м'язів-антагоністів. еханізм розвитку першої фази рефлексу заключається в наступному: при ударі по сухожилку м'яз швидко розтягується збудження м'язових веретен моносинаптична активація -мотонейронів, що інервують чотирьохголовий м'яз та гальмування (за участю вставних гальмівних нейронів) центрів м'язів-антагоністів.Механізм розвитку другої фази такий: при скороченні м'яза розтягується його сухожилки гальмування-мотонейронів, що інервують чотирьохголовий м'яз та активація -мотонейронів, що інервують м'язи-антагоністи. До обох центрів інформація передається через вставні нейрони (збудження та гальмування). 10) Рухові рефлекси заднього мозку, децеребраційна ригідність. Задній мозок – це довгастий мозок та міст. Вони зберігають ознаки сегментарної будови, але мають і надсегментарні утворення. Серед найбільш важливих рухових надсегментарних утворень – присінкові ядра та сітчаста речовина. Вони носять назву надсегментарних утворень, так як впливають на м’язи не прямо, а через мотонейрони сегментарних структур – рухові ядра спинного мозку і черепно-мозкових нервів. Задній мозок отримує і переробляє всю аферентну інформацію, що надходить від спинного мозку, оскільки всі специфічні висхідні шляхи від спинного мозку, входячи в стовбур мозку (задній та середній мозок) віддають коллатералі (гілочки) до ретикулярної формації тут продовжується обробка аферентної інформації. Задній мозок отримує нову аферентну інформацію, яка відіграє роль в регуляції рухових функцій – це інформація від вестибулярних рецепторів. Вестибулярні рецептори (перетинчастий лабіринт вискової кістки) Вестибулорецептори отолітового апарата Вестибулорецептори півкружних каналів Види Присінок (маточка та мішочок) Ампули півкружних каналів Локалізація Адекватний подразник Зміщення ендолімфи відносно волоскових рецепторних клітин Зміщення отолітової мембрани відносно волоскових рецепторних клітин Зміна положення голови в просторі Рух з прискоренням в горизонтальній чи вертикальній площині – лінійне прискорення Рух з кутовим прискоренням (обертові рухи з прискоренням) в трьох площинах Отолітові рецептори – являють собою волоскові клітини, волоски яких занурені в отолітову мембрану – желеподібна мембрана з включенням кристалів CaCО3– отолітів. Зміщення цієї мембрани деформує волоски збудження рецептора (чи його гальмування, в залежності від того, в який бік згинається волосок). Отолітова мембрана зміщується відносно волоскових клітин при двох умовах: 1. Зміщення голови в просторі. 2. Рух з лінійним прискоренням. Вестибулорецептори півколових канальців – волоскові клітини, які локалізуються в ампулах півколових канальців і занурені в ендолімфу цих канальців. Ендолімфа зміщується відносно волоскових клітин і деформує їх при русі з кутовим прискоренням. При цьому збуджуються волоскові клітини канала, який знаходиться в тій площині, в якій проходять обертальні рухи (у відповідності з трьохвимірним простором, в якому ми існуємо, наявні 3 півколових канальці). Від вестибулорецепторів інформація передається в задній мозок по аферентним волокнам VIII пари черепно-мозкових нервів (далі ЧМН) – вестибуло-кохлеарного нерва (точніше – по його вестибулярній частині). В задньому мозку розміщені 4 вестибулярні ядра (медіальне, латеральне, верхнє та нижнє). В регуляції рухових функцій у людини найбільше значення має латеральне вестибулярне ядро Дейтерса, від якого починається вестибулоспінальний шлях (частина МНС). Вивчити роль заднього мозку в регуляції рухових функцій можна на тваринах з децеребрацією – перерізкою стовбура мозку між середнім та заднім мозком (точніше – нижче червоних ядер середнього мозку). При порівнянні можливостей регуляції рухових функцій такої тварини і спінальної тварини, можна оцінити роль в регуляції цих функцій саме заднього мозку. Пі 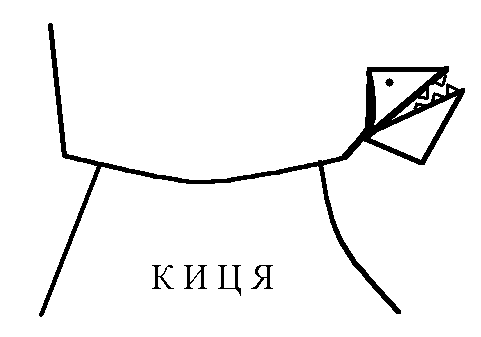 сля операції децеребрації у тварини виникає децеребраційна ригідність, яка у кішки проявляється у слідуючому: передні і задні кінцівки розігнуті; голова закинута назад за рахунок розгинання шиї; хвіст піднятий; тулуб розігнутий. Ця поза кішки пов’язана з гіпертонусом розгиначів. Позу децеребраційної тварини (децеребраційна ригідність) можна охарактеризувати як посилену позу антигравітації. Згадаємо, що спинний мозок забезпечував формування м’язового тонусу рефлекторної природи, але не забезпечував формування пози антигравітації із-за малої сили тонусу. сля операції децеребрації у тварини виникає децеребраційна ригідність, яка у кішки проявляється у слідуючому: передні і задні кінцівки розігнуті; голова закинута назад за рахунок розгинання шиї; хвіст піднятий; тулуб розігнутий. Ця поза кішки пов’язана з гіпертонусом розгиначів. Позу децеребраційної тварини (децеребраційна ригідність) можна охарактеризувати як посилену позу антигравітації. Згадаємо, що спинний мозок забезпечував формування м’язового тонусу рефлекторної природи, але не забезпечував формування пози антигравітації із-за малої сили тонусу.В 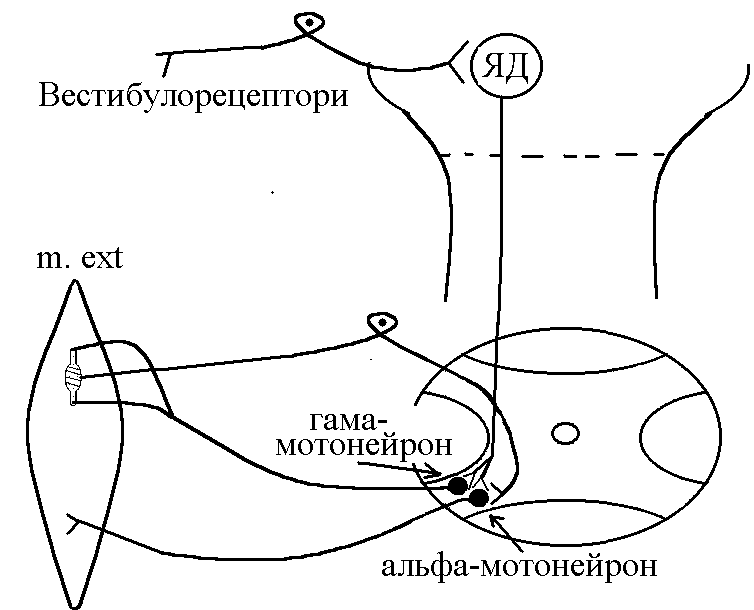 основі формування децеребраційної регідності лежать вестибулоспінальні впливи. Присінкові ядра Дейтерса тонічно активні, так як вестибулорецептори адаптуються дуже повільно. Від цих ядер по присінково-спинномозковим шляхам, що не перехрещуються, інформація поступає до мотонейронів, які інервують м’язи тулуба та проксимальних відділів кінцівок. При цьому - та -мотонейрони екстензорів активуються, а - та -мотонейрони флексорів, навпаки – гальмуються. основі формування децеребраційної регідності лежать вестибулоспінальні впливи. Присінкові ядра Дейтерса тонічно активні, так як вестибулорецептори адаптуються дуже повільно. Від цих ядер по присінково-спинномозковим шляхам, що не перехрещуються, інформація поступає до мотонейронів, які інервують м’язи тулуба та проксимальних відділів кінцівок. При цьому - та -мотонейрони екстензорів активуються, а - та -мотонейрони флексорів, навпаки – гальмуються.Активація -мотонейронів екстензорів приводить до безпосередньої активації екстрафузальних волокон цих м’язів збільшення їх тонуса. Активація -мотонейронів приводить до скорочення скоротливих сегментів інтрафузальних волокон скорочення цих сегментів розтягнення ядерної сумки інтрафузального волокна збудження рецептора передача інформації по -петлі до -мотонейронів посилення їх активності збільшення тонусу м’язів екстензорів. Протилежні зміни виникають у флексорній групі м’язів. Тому коротко можна сказати, що в основі розвитку децеребраційної ригідності лежать вестибулоспінальні впливи, завдяки яким відбувається: -- та -посилення міотатичних тонічних рефлексів екстензорів; - - та -послаблення міотатичних тонічних рефлексів флексорів. Шийні рефлекси положення – виникають у відповідь на зміну нормального положення голови відносно тулуба. При цьому подразнюються пропріорецептори шиї передача інформації в шийні сегменти спинного мозку по пропріоспінальним низхідним шляхам до мотонейронів спинного мозку перерозподіл м’язового тонусу, який змінює положення тулуба відповідно до положення голови: - якщо голова нахиляється чи повертається в бік, то зі сторони нахилу збільшується тонус розгиначів, а з протилежної сторони – тонус згиначів. Такий перерозподіл тонуса попереджає падіння, тобто втрату пози; - якщо голова нахиляється вперед, тонус згиначів збільшується на передніх кінцівках кішки, тонус розгиначів – на задніх (кицька п’є молоко чи жере Віскас); - якщо голова закидається назад, тонус розгиначів збільшується на передніх кінцівках кішки, тонус згиначів – на задніх (кицька дивиться на папугу й готується до стрибка). Присінкові (вестибулярні) рефлекси положення – виникають при зміні положення голови в просторі. При нахилах чи поворотах голови збуджуються отолітові вестиблорецептори інформація передається до вестибулярних ядер Дейтерса до мотонейронів спинного мозку та до м’язів. В результаті зі сторони нахилу (поворота) збільшується тонус розгиначів, а з протилежної сторони – тонус згиначів збереження пози. Статичні рефлекси положення Шийні Присінкові Пропріорецептори шиї – у відповідь на зміну положення голови відносносно тулуба Присінкові отолітові рецептори у відповідь на зміну положення голови у просторі З яких рецепторів починається Рівень замикання Задній мозок Спинний мозок Спрямовані на підтримання рівноважного положення тіла в просторі (на підтримання пози). Значення Перерозподіл тонусу за рахунок шийних рефлексів не можна отримати у спінальної жаби через слабкість тонуса, хоча рівень їх замикання – спинний мозок. Задній мозок підсилює ці рефлекси. Шийні та присінкові рефлекси положення у вигляді перерозподілу тонуса можна отримати у децереброваної тварини. Але на фоні ригідності ступінь цього перерозподілу недостатній для утримання антигравітаційної пози: децеребрована тварина може стояти, як лялька, але ще не може втримувати рівновагу при умовах, що вмикають шийні та присінкові статичні рефлекси положення. Таким чином, задній мозок: 1. Забезпечує створення посиленої антигравітаційної пози – пози децеребраційної ригідності. 2. Забезпечує здійснення статичних позових рефлексів (шийних та присінкових), що спрямовані на збереження рівноважної пози при зміні положення голови відносно тулуба (шийні) чи при зміні положення голови в просторі (присінкові), але ступінь виразності цих рефлексів на фоні посиленого тонуса недостатній для збереження пози. 3. Не забезпечує збереження пози при русі (немає стато-кінетичних рефлексів), відновлення порушеної пози (немає випрямляючих рефлексів). 4. Фазичні рефлекси на фоні посиленого тонуса не викликаються. Ретикулярна формація стовбура мозку, характер її висхідних та низхідних впливів. Сітчаста речовина (ретикулярна формація – РФ) – скупчення нервових клітин, які мають велику кількість дендритів, що галузяться та довгі аксони, які формують низхідні (ретикулоспінальні) та висхідні шляхи сітчастої речовини. Аферентних шляхів РФ дуже багато, вона отримує інформацію від: - усіх специфічних висхідних еферентних шляхів, які віддають колатералі до ядер РФ, заходячи в стовбур мозка; - від іних утворень ЦНС, а саме:
Еферентні зв’язки РФ можна поділити на висхідні та низхідні. Висхідні зв’язки РФ та висхідні впливи: - через неспецифічні ядра таламуса передає інформацію всім відділам головного мозку та має неспецифічний активізуючий вплив на клітини. Цей вплив забезпечує бодьорий стан організму та визначає рівень активності клітин кори. РФ має окремі структури (гіпногенні зони), котрі зменшують ступінь (рівень) активності клітин кори та зумовлюють стан сну; - по ретикулогіпоталамічним шляхам передає інформацію від різних рецепторів в гіпоталамус. Характер низхідних ретикулоспінальних впливів: - від медіального ядра РФ заднього мозку починається ретикулоспінальний (входить до складу МНС) – специфічно активує мотонейрони флексорів та гальмує мотонейрони екстензорів тулуба та проксимальних відділів кінцівок; - від інших ядер РФ беруть початок шляхи, які неспецифічно активують чи гальмують спінальні мотонейрони і таким чином визначають ступінь їхньої активності; - від вегетативних центрів РФ (дихальний, гемодинамічний) по ретикулоспінальним шляхам передається інформація до симпатичних рухових нейронів спинного мозку при регуляції відповідних функцій організму. |