Общая физиология цнс
 Скачать 0.64 Mb. Скачать 0.64 Mb.
|

|
11. Гипоталамус. Характеристика основных ядерных групп. Участие гипоталамуса в регуляции вегетативных функций и в формировании эмоций и мотиваций. Нейронная организация. Гипоталамус образован группой небольших ядер, расположенных у основания мозга, вблизи гипофиза. Клеточные ядра, образующие гипоталамус представляют собой высшие подкорковые центры вегетативной нервной системы и все> жизненно важных функций организма. В эволюционном плане гипоталамус являете* очень древним образованием. Он хорошо развит уже у наиболее примитивных представителей позвоночных. Организация гипоталамуса сохраняет сходные черты на разных ступенях эволюции, что обусловлено известным постоянством его функций; Скопление нейронных образований, образующих гипоталамус, может быть подразделено на преоптическую, переднюю, среднюю, наружную изаднюю группы ядер. В пре оптическую область входят пёривентрикулярное, медиальное и латеральное преоптические ядра. В группу ядер переднего гипоталамуса относят супраоптическое, супрахиазматическое и паравентрикулярное ядра. Средний гипоталамус составляет вентромедиаль- ное и дорсомедиальное ядра. К. наружной группе ядер относятся латеральное гипота- ламическое ядро и ядро серого бугра. Наконец, в заднем гипоталамусе различают заднее гипоталамическоеперифорникальное, премамиллярное, медиальное мамиллярное, суп- рамамиллярное и латеральное мамиллярное ядра. Схема главных- ядер гипоталамуса представлена на рис. 93. Организация гипоталамуса характеризуется обширными и очень сложными афферентными и эфферентными связями. - Афферентные сигналы в. гипоталамус поступают из коры больших полушарий, из таламических структур, ядер базальных ганглиев. Одним из основных эфферентных путей является медильный мозговой пучок, или паравентрикулярная система* и мамилло- тегментальный тракт. Волокна этих путей идут в каудальном направлении по стенкам водопровода мозга или сильвиева, водопровода, дают Многочисленные ответвления ж структурам среднего мозга. Аксоны клеток гипоталамических ядер образуют также большое количество коротких эфферентных путей, идущих в таламическую и субталамическую области и в другие подкорковые образования. Ядра переднего гипоталамуса — супраоптическое и паравентрикулярное, кроме того, связаны с гипофизом особой системой волокон, которые служат не только^для проведения электрических сигналов, но и для транспорта продуктов нейросекреции, которые вырабатываются нейронами, этих ядер. Функции гипоталамуса Существуют экспериментальные данные о наличии в гипоталамусе центра сна и центра пробуждения. Гипоталамус играет важную роль в терморегуляции: Раздражение задних ядер при- юдит к гипертермии в результате повышения теплопродукции при интенсификации обменных процессов, а также вследствие дрожи скелетной мускулатуры. В области средних и боковых ядер имеются группы нейронов, рассматриваемых как центры насыщения и голода. Стимулом для изменения их деятельности являются отклонения в химическом составе притекающей крови. При голодании в крови, происходит снижение содержания аминокислот, мирных сислот, глюкозы и других веществ. Это приводит к активации определенных гипотала- лических нейронов и развитию сложных поведенческих реакций Организма, направленна на утоление чувства голода.' Приспособительные поведенческие реакции развиваются при недостатке в организме воды, что приводит к появлению чувства- жажды вследствие активации гипоталами- [еских зон, расположенных дорсолатерально. от супраоптического ядра. В результате >езко усиливается потребление воды(полидипсия). Наоборот, разрушение указанных •ипоталамических центров приводит к отказу от воды (адипсия). В гипоталамусе расположены центры,'связанные с регуляцией полового поведения. )пыты с вживлением электродов ^ эти центры (задний гипоталамус) показали, что при федоставлении животному возможности самораздражения (путем нажатия педали, жлючающей ток, проходящий через вживленные электроды) оно может проводить само- >аздражение с высокой'частотой в течение длительного времени. Поэтому эти центры бы- ш названы центрами удовольствия. Установлено, что они являются компонентом нейрон- юй системы, принимающей участие в,регуляции эмоциональной сферы полового поведения. Гипоталамус принимает участие в процессе чередования сна и бодрствования. Супраоптическое ядро и супраоптико-гипофизарный тракт связаны с задней долей 'ипофиза, выделяющей в кровь ряд гормонов. Установлено, что по крайней мере часть этих гормонов (многие из которых являются полипептидами) может выделяться окон чаниями нейронов других отделов нервной системы в качестве нейромедиаторов или ней- ромодуляторов. Задняя доля гипофиза, являющаяся по происхождению производным нервной системы (нейрогипофиз)*, специализировалась на депонировании и выделение указанных веществ в кровяное русло. Эти вещества продуцируются клетками супраопти- чес кого ядра и передаются в кровь по их аксонам в результате генерации потенциал? действия в нейрогипофиз подобно тому как потенциалы действия, приходящие в аксо нальные окончания обычных нервных клеток, вызывают процесс высвобождения ме диатора. Основными гормонами, выделяемыми задней долей гипофиза, являются антидиуретический, гормон, регулирующий водный метаболизм, а также гормоны, регулирующие деятельность матки, функцию молочных желез. . По-иному осуществляется связь гипоталамуса с передней долей гипофиза (аденоги- пофиз),. вырабатывающего такие гормоны белковой природы, как адренокортикотроп- ный, фолликулостимулирующий и лютеинизирующий, тиреотропный, гормон роста, v средней долей гипофиза (меланофорный гормон). Регуляция гипоталамусом этой част* гипофиза осуществляется через кровь --нейрогуморальным путем. Важнейшие функ ции гипоталамуса приведены в табл 12. Лимбическая система, особенности ее структурно-функциональной организации. Основные функции лимбической системы. Лимбическая система — совокупность нервных структур и их связей, расположенных в медиобазальной части больших полушарий, участвующих в управлении вегетативными функциями и эмоциональным, инстинктивным поведением, а также оказывающих влияние на смену фаз сна и бодрствования. К лимбической системе относится наиболее древняя часть коры головного мозга, расположенная на внутренней стороне больших полушарий. К ней относятся: гиппокамп, поясная извилина, миндалевидные ядра, грушевидная извилина. Лимбические образования относятся к высшим интегративным центрам регуляции вегетативных функций организма. Нейроны лимбической системы получают импульсы с коры, подкорковых ядер, таламуса, гипоталамуса, ретикулярной формации и всех внутренних органов. Характерным свойством лимбической системы является наличие хорошо выраженных кольцевых нейронных связей, объединяющих различные ее структуры. Среди структур, ответственных за память и обучение, главную роль играют гиппокамп и связанные с ним задние зоны лобной коры. Их деятельность важна для перехода кратковременной памяти в долговременную. Лимбическая система участвует в афферентном синтезе, в контроле электрической активности мозга, регулирует процессы обмена веществ и обеспечивает ряд вегетативных реакций. Раздражение различных участков этой системы у животного сопровождается проявлениями оборонительного поведения и изменениями деятельности внутренних органов. Лимбическая система участвует и в формировании поведеческих реакций у животных. В ней находится корковый отдел обонятельного анализатора. Структурно-функциональная организация лимбической системы Большой круг Пейпеса:
Малый круг Наута:
Лимбическая система состоит из филогенетически старых отделов переднего мозга. В названии (limbus — край) отражена особенность ее расположения в виде кольца между новой корой и конечной частью ствола мозга. К лимбической системе относят ряд функционально объединенных структур среднего, промежуточного и конечного мозга. Это поясная, парагиппокампальная и зубчатая извилины, гиппокамп, обонятельная луковица, обонятельный тракт и прилежащие участки коры. Кроме того, к лимбической системе относят миндалину, переднее и септальное таламические ядра, гипоталамус и мамиллярные тела. Лимбическая система имеет множественные афферентные и эфферентные связи с другими структурами мозга. Ее структуры взаимодействуют друг с другом. Функции лимбической системы реализуются на основе протекающих в ней интегративных процессов. В то же время отдельным структурам лимбической системы присущи более или менее очерченные функции. 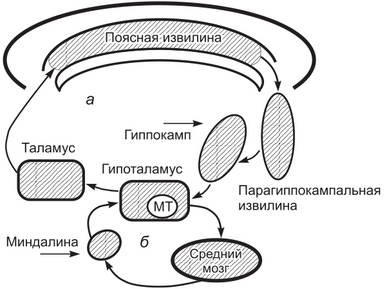 Рис. 1. Важнейшие связи между структурами лимбической системы и ствола мозга: а — круг Пайпеца, б — круг через миндалину; МТ — мамиллярные тела Основные функции лимбической системы:
13. Базальные ядра и их структурно-функциональная организация. Основные функции базальных ядер. Базальные ганглии, или подкорковые ядра, — это тесно связанные между собой структуры мозга, расположенные в глубине больших полушарий между лобными долями и промежуточным мозгом. Базальные ганглии являются парными образованиями и состоят из ядер серого вещества, разделенных прослойками белого — волокон внутренней и наружной капсул мозга. В состав базальных ганглиев входят: полосатое тело, состоящее из хвостового ядра и скорлупы, бледный шар и ограда. С функциональной точки зрения иногда к понятию базальных ганглиев относят также субталамическое ядро и черную субстанцию (рис. 1). Большой размер этих ядер и подобие в структуре у различных видов дают основание предполагать, что они вносят большой вклад в организацию работы мозга наземных позвоночных животных. Основные функции базальных ганглиев:
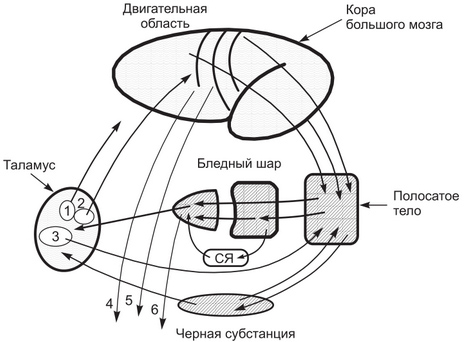 Рис. 1. Важнейшие афферентные и эфферентные связи базальных ганглиев: 1 паравентрикулярное ядро; 2 вентролатеральное ядро; 3 срединные ядра таламуса; СЯ — субталамическое ядро; 4 — кортикоспинальный тракт; 5 — кортикомостовой тракт; 6 — эфферентный путь от бледного шара к среднему мозгу Из клинических наблюдений давно известно, что одним из последствий заболеваний базальных ганглиев является нарушение тонуса мышц и движений. На этом основании можно было бы предполагать, что базальные ганглии должны быть связаны с моторными центрами ствола и спинного мозга. Современными методами исследования показано, что аксоны их нейронов не следуют в нисходящем направлении к моторным ядрам ствола и спинного мозга, а повреждение ганглиев не сопровождается парезами мышц, как это имеет место при повреждении других нисходящих моторных путей. Большая часть эфферентных волокон базальных ганглиев следует в восходящем направлении к моторным и другим областям коры больших полушарий мозга. Афферентные связи Структурой базальных ганглиев, к нейронам которой поступает большая часть афферентных сигналов, является полосатое тело. Его нейроны получают сигналы из коры больших полушарий мозга, ядер таламуса, клеточных групп черной субстанции промежуточного мозга, содержащих дофамин, и от нейронов ядра шва, содержащих серотонин. При этом нейроны скорлупы полосатого тела получают сигналы преимущественно из первичной соматосенсорной и первичной моторной коры, а нейроны хвостатого ядра (уже предварительно интегрированные полисенсорные сигналы) из нейронов ассоциативных областей коры больших полушарий мозга. Анализ афферентных связей базальных ядер с другими структурами мозга предполагает, что от них в ганглии поступает не только информация, связанная с движениями, но и информация, которая может отражать состояние общей активности мозга и быть связана с его высшими, познавательными функциями и эмоциями. Полученные сигналы подвергаются в базальных ганглиях сложной обработке, в которой участвуют его различные структуры, связанные между собой многочисленными внутренними связями и содержащие различные типы нейронов. Среди этих нейронов большинство составляют ГАМК-ергические нейроны полосатого тела, которые посылают аксоны к нейронам бледного шара и черной субстанции. Эти нейроны продуцируют также динорфин и энкефалин. Большой удельный вес в передаче и обработке сигналов внутри базальных ганглиев занимают его возбуждающие холинергические интернейроны с широко ветвящимися дендритами. К этим нейронам конвергируют аксоны нейронов черной субстанции, секретирующие дофамин. Эфферентные связи базальных ганглиев используются для посылки сигналов, обработанных в ганглиях, в другие структуры мозга. Нейроны, формирующие основные эфферентные пути базальных ганглиев, располагаются главным образом в наружном и внутреннем сегментах бледного шара и в черной субстанции, получающих афферентные сигналы в основном из полосатого тела. Часть эфферентных волокон бледного шара следует в интраламинарные ядра таламуса и оттуда — в полосатое тело, образуя подкорковую нейронную сеть. Большая часть аксонов эфферентных нейронов внутреннего сегмента бледного шара следует через внутреннюю капсулу к нейронам вентральных ядер таламуса, а от них — в префронтальную и дополнительную моторную кору больших полушарий. Через связи с моторными областями коры мозга базальные ганглии оказывают влияние на контроль движений, осуществляемый корой через кортикоспинальный и другие нисходящие двигательные пути. Хвостатое ядро получает афферентные сигналы с ассоциативных областей коры мозга и, обработав их, посылает эфферентные сигналы преимущественно в префронтальную кору. Предполагается, что эти связи являются основой для участия базальных ганглиев в решении задач, связанных с подготовкой и исполнением движений. Так, при повреждении хвостатого ядра у обезьян нарушается способность выполнять движения, требующие сведений из аппарата пространственной памяти (например, учета, где расположен предмет). Базальные ганглии связаны эфферентными связями с ретикулярной формацией промежуточного мозга, через которые участвуют в контроле ходьбы, а также с нейронами верхних холмиков, через которые они могут контролировать движения глаз и головы. С учетом афферентных и эфферентных связей базальных ганглиев с корой и другими структурами мозга выделяют несколько нейронных сетей или петель, проходящих через ганглии или заканчивающихся внутри их. Моторная петляобразована нейронами первичной моторной, первичной сенсомоторной и дополнительной моторной коры, чьи аксоны следуют к нейронам скорлупы и затем через бледный шар и таламус достигают нейронов дополнительной моторной коры. Глазодвигательная петля образована нейронами моторных полей 8, 6 и сенсорного поля 7, аксоны которых следуют в хвостатое ядро и далее к нейронам лобного глазного поля 8. Префронтальные петли образованы нейронами префронтальной коры, аксоны которых следуют к нейронам хвостатого ядра, черного тела, бледного шара и вентральных ядер таламуса и затем достигают нейронов прсфронтальной коры. Каемчатая петля образована нейронами круговой извилины, орбитофронтальной коры, некоторых областей височной коры, тесно связанных со структурами лимбической системы. Аксоны этих нейронов следуют к нейронам вентральной части полосатого тела, бледного шара, медиодорсального таламуса и далее — к нейронам тех областей коры, в которых петля начиналась. Как можно видеть, каждая петля формируется множественными корковостриарными связями, которые после их прохождения через базальные ганглии следуют через ограниченную область таламуса в определенную одиночную область коры. Области коры, посылающие сигналы в ту или иную петлю, функционально связаны друг с другом. Функции базальных ганглиев Нейронные петли базальных ганглиев являются морфологической основой выполняемых ими основных функций. Среди них — участие базальных ганглиев в подготовке и осуществлении движений. Особенности участия базальных ганглиев в выполнении этой функции вытекают из наблюдений за характером нарушения движений при заболеваниях ганглиев. Предполагается, что базальные ганглии играют важную роль в планировании, программировании и выполнении сложных движений, инициируемых корой больших полушарий. С их участием абстрактный замысел движения превращается в моторную программу сложных произвольных действий. Их примером могут быть такие действия, как одновременное осуществление нескольких движений в отдельных суставах. Действительно, при регистрации биоэлектрической активности нейронов базальных ганглиев во время выполнения произвольных движений отмечается се повышение в нейронах субталамических ядер, ограды, внутреннего сегмента бледного шара и ретикулярной части черного тела. Повышение активности нейронов базальных ганглиев инициируется притоком возбуждающих сигналов к нейронам полосатого тела из коры больших полушарий, опосредованных высвобождением глутамата. К этим же нейронам поступает поток сигналов из черной субстанции, оказывающий на нейроны полосатого тела притормаживающее действие (через высвобождение ГАМК) и способствующий фокусированию влияния нейронов коры на определенные группы нейронов полосатого тела. В это же время к его нейронам поступают афферентные сигналы из таламуса с информацией о состоянии активности других областей мозга, имеющих отношение к организации движений. Нейроны полосатого тела интегрируют все эти потоки информации и передают ее нейронам бледного шара и ретикулярной части черной субстанции и далее но эфферентным путям эти сигналы передаются через таламус в моторные области коры мозга, в которых осуществляется подготовка и инициирование предстоящего движения. Предполагается, что базальные ганглии еще на этапе подготовки движения осуществляют выбор типа движения, необходимого для достижения поставленной цели, отбор мышечных групп, необходимых для его эффективного выполнения. Вероятно, базальные ганглии участвуют в процессах моторного обучения путем повторения движений, причем их роль заключается в выборе оптимальных путей осуществления сложных движений для достижения желаемого результата. С участием базальных ганглиев достигается устранение избыточности движений. Еще одной из моторных функций базальных ганглиев является участие в осуществлении автоматических движений или моторных навыков. Когда базальные ганглии повреждены, человек выполняет их в более замедленном темпе, менее автоматизировано, с меньшей точностью. Двустороннее разрушение или повреждение ограды и бледного шара у человека сопровождается возникновением навязчиво-принудительного двигательного поведения и появлением элементарных стереотипных движений. Двустороннее повреждение или удаление бледного шара ведет к снижению двигательной активности и гипокинезии, в то время как одностороннее повреждение этого ядра или не влияет, или слабо сказывается на двигательных функциях. |