Общая физиология цнс
 Скачать 0.64 Mb. Скачать 0.64 Mb.
|

|
Общая физиология ЦНС 1. Нейрон как структурная и функциональная единица ЦНС, его физиологические свойства и взаимосвязь с глиальными клетками. Объединение нейронов в нервные цепи. Нейрон или по-другому нервная клетка, состоящая из тела и отходящих от нее отростков – относительно коротких дендритов и длинного аксона. Ф-ции сомы: обеспечивает метаболизм клетки. В ней синтезируется медиатор. Восприятие возбуждения. Ф-я дендрита: воспринимает возбуждения. Ф-я аксонного холмика: здесь образуется ПД, его мембрана наиболее возбудима. ПД не может быть ни на соме, ни на дендритах. Ф-я аксона: проведение ПД, возбуждение. Взаимосвязь: нейроны и глиальные клетки разделены межклеточными щелями. Глиальныеклетки окружают нейрон и играют вспомогательную роль. Они выполняют опорную и защитную функции и называются глиальнымиклетками или глией. Функциональная классификация: 1.Афферентные (чувствительные, сенсорные – воспринимают сигналы из внешней или внутренней среды); 2.Вставочные связывающие нейроны друг с другом (обеспечивают передачу информации внутри ЦНС: с афферентных нейронов на эфферентные). 3. Эфферентные (двигательные, мотонейроны – передают первые импульсы от нейрона к исполнительным органам). Главная структурная особенностьнейрона – наличие отростков (дендритов и аксонов). 1– дендриты; 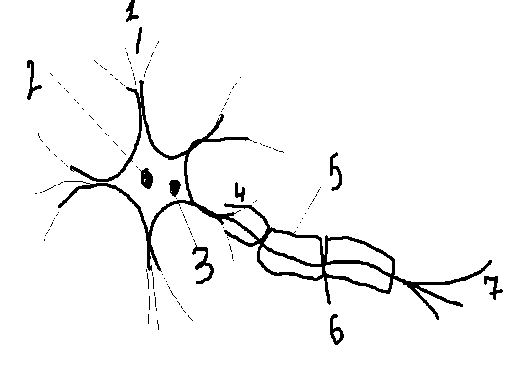 2 – тело клетки; 3 – аксонный холмик; 4 – аксон; 5 –Швановская клетка; 6 – перехват Ранвье; 7 – эфферентные нервные окончания. Последовательное синоптическое объединение всех 3х нейронов образует рефлекторную дугу. 2. Особенности передачи возбуждения в синапсах ЦНС. Возбуждающие и тормозные синапсы и их медиаторные механизмы, ВПСП и ТПСП (тормозной псп). В синапсах ЦНС возбуждение проводится медленнее, чем в нервных волокнах. Это объясняется особенностями синаптической передачи (синаптическая задержка), а именно диффузией медиатора к постсинаптической мембраны, возникновением ВПСП (возбуждающий постинаптический потенциал), нарастанием его до критической величины, генерацией ПД. Синапс возбуждающий – синапс, в котором возбуждается постсинаптическая мембрана; в ней возникает ВПСП и пришедшее к синапсу возбуждение распространяется дальше. Во-первых в ЦНС могут сущ-ть синапсы не только с химическим, но и с электрическим, а также и со смешанным механизмом передачи. Чисто электрические синапсы чаще образуются м/д дендритами однотипных, близко расположенных нейронов. Электрические синапсы способны к двухстороннему проведению возбуждения. Во-вторых возбуждение возникает только на аксоном холмике при достижении там критического уровня деполяризации. Достигается Екр в рез-те суммации ПСП. 3. – 5. Основные принципы и особенности распространения возбуждения в ЦНС (дивергенция, конвергенция, реверберация, окклюзия, облегчение и др.). Основные принципы координационной деятельности ЦНС. Роль доминанты (А.А. Ухтомский). Конвергенция - схождение к одному нейрону двух или нескольких возбуждений от сенсорных раздражителей (например звук, свет). Дивергенция- направляется в разные стороны - способность одиночного нейрона устанавливать многочисленные синаптические связи с различными нервными клетками. Иррадиация в физиологии, распространение процесса возбуждения или торможения в центральной нервной системе. Важную роль И. играет в деятельности коры больших полушарий головного мозга. Реверберация - длительная циркуляция импульсов возбуждения в сетях нейронов или между различными структурами ц.н.с. Одностороннее проведение возбуждения- возбуждение передается с афферентного на эфферентный нейрон. Причина: клапанное свойство синапса. Координационная деятельность ЦНС — это согласование деятельности различных отделов ЦНС с помощью упорядочения распространения возбуждения между ними. Основой координационной деятельности ЦНС является взаимодействие процессов возбуждения и торможения. Если два нервных центра рефлекторных реакций имеют частично перекрываемые рецептивные поля, то при совместном раздражении обоих рецептивных полей реакция будет меньше, чем арифметическая сумма реакций при изолированном раздражении каждого из рецептивных полей — феномен окклюзии. Феномен облегчения участвует в процессах обеспечения координационной деятельности ЦНС при выработке навыков. Недостаточно координированные движения в начале выработки навыка постепенно становятся более точными — координированными. Дополнительные, ненужные движения постепенно устраняются. Возбуждение распространяется в ЦНС быстрее по проторенным путям, возбудимость которых повышена Доминанта играет важную роль в координационной деятельности ЦНС. Доминанта — это стойкий, господствующий очаг возбуждения, подчиняющий себе активность других нервных центров. Доминантное состояние двигательных центров обеспечивает автоматизированное выполнение двигательных актов, например, в процессе трудовой деятельности человека, при выполнении гимнастических элементов и т.п. Реципрокная (сочетанная) связь — вид функциональной связи, обеспечивающий торможение центра-антагониста при возбуждении центра-агониста. Например, при вызове сгибательного рефлекса конечности импульсы из рефлексогенной зоны (кожа) поступают через вставочные нейроны к мотонейронам центра мышц-сгибателей, а также одновременно — к центру- антагонисту (мышц-разгибателей), но с включением на пути тормозного нейрона, который образует тормозный синапс на нейронах центра-разгибателя. Мышцы-разгибатели поэтому не сокращаются и не препятствуют сгибанию конечности. Реципрокные взаимоотношения между центрами встречаются довольно широко. Так, при возбуждении центра глотания тормозится центр жевания, рефлекс глотания тормозит вдох, возбуждение центра вдоха тормозит центр выдоха. Прямая связь — управление другим центром (ядром) или рабочим органом с помощью посылки к ним эфферентных импульсов (команд). Например, нейроны дыхательного центра продолговатого мозга посылают импульсы к сс-мотонейронам спинного мозга, от которых нервные импульсы поступают к дыхательным мышцам. Мозжечок посылает импульсы к ядрам ствола мозга и т.д. Обратная связь (обратная афферентация) — управление нервным центром или рабочим органом с помощью афферентных импульсов, поступающих от них. В данном случае центр имеет, естественно, и прямую связь с образованиями, функцию которых контролирует, но обратная афферентация делает прямую связь более совершенной в функциональном отношении (принцип обратной связи в регуляции функций организма). Если нарушить прямую связь центра с регулируемым центром или органом, то управление становится вообще невозможным. Если же нарушить только обратную связь, управление сильно страдает. Фактор силы. Известно, что к одному и тому же центру могут подходить пути от различных рефлексогенных зон{принцип общего конечного пути). В случае их одномоментной активации центр будет реагировать на более сильное возбуждение. Например, слабое раздражение кожи туловищау собаки вызывает чесательный рефлекс нижней конечности — собака почесывает кожу туловища. Одностороннее проведение возбуждения в химических синапсах ЦНС способствует упорядочению распространения возбуждения, ограничивая иррадиацию возбуждения в ЦНС. Фактор субординации — подчинение нижележащих отделов ЦНС вышележащим. Например, пирамидные клетки коры большого мозга, нейроны красного ядра управляют активностью а- и у-мотонейронов спинного мозга. 4. Торможение в ЦНС (И.М. Сеченов), его виды и роль. Современное представление о механизмах центрального торможения. Интегративная и координационная деятельность центральных нервных образований осуществляется при обязательном участии тормозных процессов. Торможение в ЦНС открыл И.М.Сеченов (1863). В опыте на таламической лягушке он определял латентное время сгибательного рефлекса при погружении задней конечности в слабый раствор серной кислоты. Было показано, что латентное время рефлекса значительно увеличивается, если на зрительный бугор предварительно положить кристаллик поваренной соли. Открытие И.М.Сеченова послужило толчком для дальнейших исследований торможения в ЦНС. В частности, обнаружил проявления торможения у спинальной лягушки Ф.Гольц (1870). Он также исследовал латентное время рефлекса. При этом оказалось, что механическое раздражение кончиков пальцев одной конечности лягушки существенно удлиняет латентный период сгибательного рефлекса другой конечности при погружении ее в раствор кислоты.Наличие специальных тормозных структур в продолговатом мозге доказал Х.Мегун (1944). В опытах на кошках при изучении разгиба- тельного рефлекса Х.Мегун установил, что раздражение медиальной части ретикулярной формации продолговатого мозга тормозит рефлекторную активность спинного мозга. Тонкий анализ тормозных явлений в ЦНС позволил выделить две разновидности торможения: 1) постсинаптическое торможение и 2) пресинаптическое торможение. ПОСТСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ А. Этот вид торможения открыл Д.Экклс (1952) при регистрации потенциалов мотонейронов спинного мозга у кошки во время раздражения мышечных афферентов группы 1а. При этом оказалось, что в мотонейронах мышцы-актагониста регистрируются не деполяризация и возбуждение, агиперполяризационный постсинаптический потенциал, уменьшающий возбудимость мотонейрона, угнетающий его способность реагировать на возбуждающие влияния. По этой причине вызванный гиперполяризационный потенциал был названтормозным постсинаптическимпотенциалом, ТПСП. У кошки ТПСП регистрируется на 0,5 мс позже, чем ВПСП, что объясняется наличием на пути проведения возбуждения, запускающего ТПСП, одного дополнительного синапса. Б. Механизм постсинаптического торможения. Возбудимость клетки от ТПСП (гиперполяризационного постсинаптического потенциала) уменьшается, потому что увеличивается пороговый потенциал (AV), так как Екр. (критический уровень деполяризации, КУД) остается на прежнем уровне, а мембранный потенциал (Ео) возрастает. ТПСП возникает под влиянием и аминокислоты глицина, и ГАМК — гамма-аминомасляной кислоты. В спинном мозге глицин выделяется особыми тормозными клетками (клетки Реншоу) в синапсах, образуемых этими клетками на мембране нейрона-мишени. Действуя на ионотропный рецептор постсинаптической мембраны, глицин увеличивает ее проницаемость для СГ, при этом СП поступает в клетку согласно концентрационному градиенту вопреки электрическому градиенту, в результате чего развивается гиперполяризация. В бесхлорной среде тормозная роль глицина не реализуется. Разновидности постсинаптического торможения. Возвратное постсинаптическое торможение — торможение, при котором тормозные вставочные нейроны действуют на те же нервные клетки, которые их активируют. Параллельное торможение может выполнять подобную же роль, когда возбуждение блокирует само себя, за счет дивергенции по коллатерали с включением тормозной клетки на своем пути и возвратом импульсов к нейрону, который активировался этим же возбуждением. Латеральное постсинаптическое торможение. Тормозные вставочные нейроны соединены таким образом, что они активируются импульсами от возбужденного центра и влияют на соседние клетки с такими же функциями. ПРЕСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ А. Открытие. Пресинаптическое торможение первоначально выявлено также в спинном мозге в опыте с регистрацией активности мотонейронов моносинаптической рефлекторной дуги при раздражении антагонистических мышечных нервов. Так, известно, что раздражение первичных афферентов мышечных веретен сопровождается возбуждением гомонимных а-мотонейронов (а-мотонейронов этой же мышцы). Однако опережающее раздражение афферентов сухожильных рецепторов мышц-антагонистов предотвращает возбуждение активируемых а-мотонейронов. Мембранный потенциал и возбудимость исследуемых а-мотонейронов не изменялись либо регистрировался низкоамплитудный ВПСП, недостаточный для возникновения ПД. Мотонейроны не возбуждаются вследствие процессов, происходящих в пресинаптическом окончании, что определяет название этого вида торможения. Б. Механизм пресинаптического торможения. Электрофизиологическое изучение процессов на уровне пресинаптических окончаний регистрируется выраженная и продолжительная деполяризация, что и ведет к развитию торможения. В очаге деполяризации нарушается процесс распространения возбуждения — следовательно, поступающие импульсы, не имея возможности пройти зону деполяризации в обычном количестве и обычной амплитуде, не обеспечивают выделения медиатора в синаптическую щель в достаточном количестве, поскольку мало ионов Са2+ входит в нервное окончание — нейрон не возбуждается, его функциональное состояние, естественно, остается неизменным. Деполяризацию пресинаптической терминали вызывают специальные тормозные вставочные клетки, аксоны которых образуют синапсы на пресинаптических окончаниях аксона- мишени. Торможение (деполяризация) после одного афферентного залпа продолжается 300—400 мс, медиатором является гамма-аминомасляная кислота (ГАМК), которая действует на ГАМК,-рецепторы. Деполяризация является следствием повышения проницаемости для СГ, в результате чего он выходит из клетки. 6. Функциональные особенности соматической и вегетативной (автономной) нервной систем. Первое и основное отличие ВНС от соматической состоит в расположении эфферентного (моторного) нейрона. В соматической НС вставочный и моторный нейроны располагаются в сером веществе спинного мозга, в ВНС эффекторный нейрон вынесен на периферию, за пределы спинного мозга, и лежит в одном из ганглиев – пара-, превертебральном или интраорганном. Более того, в метасимпатической части ВНС весь рефлекторный аппарат полностью находится в интрамуральных ганглиях и нервных сплетениях внутренних органов. Второе отличие касается выхода нервных волокон из ЦНС. Соматические нервные волокна покидают спинной мозг сегментарно и перекрывают иннервацией не менее трех смежных сегментов. Волокна же ВНС выходят из трех участков ЦНС – головного мозга, грудопоясничного и крестцового отделов спинного мозга. Они иннервируют все органы и ткани без исключения. Большинство висцеральных систем имеет тройную – симпатическую, парасимпатическую и метасимпатическую – иннервацию. Третье отличие касается иннервации органов соматической и ВНС. Перерезка у животных вентральных корешков спинного мозга сопровождается полным перерождением всех соматических эфферентных волокон. Она не затрагивает дуги автономного рефлекса ввиду того, что ее эффекторный нейрон вынесен в пара- или превертебральный ганглий. В этих условиях эффекторный орган управляется импульсами данного нейрона. Именно это обстоятельство подчеркивает относительную автономию указанного отдела нервной системы. Четвертое отличие относится к свойствам нервных волокон. В ВНС, они в большинстве своем безмякотные или тонкие мякотные, как, например, преганглионарные волокна, диаметр которых не превышает 5 мкм. Такие волокна принадлежат к типу В. Постганглионарные волокна еще тоньше, большая часть их лишена миелиновой оболочки, они относятся к типу С. В отличие от них соматические эфферентные волокна толстые, мякотные, диаметр их составляет 12 – 14 мкм. Кроме того, пре- и постганглионарные волокна отличаются низкой возбудимостью. Для вызова в них ответной реакции необходима значительно большая, чем для моторных соматических волокон, сила раздражения. Волокна ВНС характеризуются большим рефрактерным периодом. Скорость распространения по ним нервных импульсов невелика и составляет в преганглионарных волокнах до 18 м/с, в постганглионарных – до 3 м/с. 7. Сравнительная характеристика симпатического и парасимпатического отделов вегетативной нервной системы, синергизм и относительный антагонизм их влияний.
Существуют антагонитистические отношения между симпатической и парасимпатической частями (однонаправленное синергическое влияние). Но, например слюноотделение стимулируется раздражением волокон симпатической и парасимпатической природы, так что здесь проявляется согласованная реакция, необходимая для пищеварения. Проявляется синергизм противоположных симпатических и парасимпатических влияний=это разнонаправленные влияния симпатических и парасипатических нервов. Например, сочетание усиления сердечной деятельности с одновременным снижением активности ЖКТ. ПСНС – сужение зрачка, усиление секреции слезной железы, урежение сердечного ритма, сужение просвета бронхов, уменьшение потоотделения, усиление моторики желудка. |