Общая физиология возбудимых тканей
 Скачать 472.21 Kb. Скачать 472.21 Kb.
|

|
4 вопрос Мембранный потенциал (или потенциал покоя) – это разность потенциалов между наружной и внутренней поверхностью мембраны в состоянии относительного физиологического покоя. Потенциал покоя возникает в результате двух причин: 1) неодинакового распределения ионов по обе стороны мембраны; 2) избирательной проницаемости мембраны для ионов. В состоянии покоя мембрана неодинаково проницаема для различных ионов. Клеточная мембрана проницаема для ионов K, малопроницаема для ионов Na и непроницаема для органических веществ. За счет этих двух факторов создаются условия для движения ионов. Это движение осуществляется без затрат энергии путем пассивного транспорта – диффузией в результате разности концент-рации ионов. Ионы K выходят из клетки и увеличивают положительный заряд на наружной поверхности мембраны, ионы Cl пассивно переходят внутрь клетки, что приводит к увеличению положительного заряда на наружной поверхности клетки. Ионы Na накапливаются на наружной поверхности мембраны и увеличивают ее положительный заряд. Органические соединения остаются внутри клетки. В результате такого движения наружная поверхность мембраны заряжается положительно, а внутренняя – отрицательно. Внутренняя поверхность мембраны может не быть абсолютно отрицательно заряженной, но она всегда заряжена отрицательно по отношению к внешней. Такое состояние клеточной мембраны называется состоянием поляризации. Движение ионов продолжается до тех пор, пока не уравновесится разность потенциалов на мембране, т. е. не наступит электрохимическое равновесие. Момент равновесия зависит от двух сил: 1) силы диффузии; 2) силы электростатического взаимодействия. Значение электрохимического равновесия: 1) поддержание ионной асимметрии; 2) поддержание величины мембранного потенциала на постоянном уровне. В возникновении мембранного потенциала участвуют сила диффузии (разность концентрации ионов) и сила электростатического взаимодействия, поэтому мембранный потенциал называется концентра-ционно-электрохимическим. Для поддержания ионной асимметрии электрохимического равновесия недостаточно. В клетке имеется другой механизм – натрий-калиевый насос. Натрий-калиевый насос – механизм обеспечения активного транспорта ионов. В клеточной мембране имеется система переносчиков, каждый из которых связывает три иона Na, которые находятся внутри клетки, и выводит их наружу. С наружной стороны переносчик связывается с двумя ионами K, находящимися вне клетки, и переносит их в цитоплазму. Энергия берется при расщеплении АТФ. Любая живая клетка поддерживает на мембране определенной величины потенциал. Его величина колеблется в значительных пределах, у нервных и мышечных клеток его величина составляет обычно 60 - 90 мВ, у других тканей не превышает 10 мВ. Различные внешние воздействия, способные изменяющие ионную проницаемость мембраны (то есть повлиять на воротный механизм канала), вызывают изменения величины ПП. Его уменьшение называется деполяризацией, а увеличение - гиперполяризацией мембраны 5 вопрос Возбужде́ние в физиологии — ответ ткани на раздражение, проявляющийся помимо неспецифических реакций (генерация потенциала действия, метаболические изменения) в выполнении специфической для этой ткани функции; возбудимыми являются нервная (проведение возбуждения), мышечная (сокращение) и железистая (секреция) ткани. Возбудимость — свойство клеток отвечать на раздражение возбуждением. Реакция клеток, тканей на раздражитель определяется законами раздражения 1. Закон "все или ничего": При допороговых раздражениях клетки в ткани ответной реакции не возникает. При пороговой силе раздражителя развивается максимальная ответная реакция, поэтому увеличение силы раздражения выше пороговой не сопровождается ее усилением. В соответствии с этим законом реагирует на раздражения одиночное нервное и мышечное волокно, сердечная мышца. 2. Закон силы: Чем больше сила раздражителя, тем сильнее ответная реакция. Однако выраженность ответной реакции растет лишь до определенного максимума. Закону силы подчиняется целостная скелетная, гладкая мышца, так как они состоят из многочисленных мышечных клеток, имеющих различную возбудимость. 3. Закон силы-длительности. Между силой и длительностью действия раздражителя имеется определенная взаимосвязь. Чем сильнее раздражитель, тем меньшее время требуется для возникновения ответной реакции. Зависимость между пороговой силой и необходимой длительностью раздражения отражается кривой силы длительности. По этой кривой можно определить ряд параметров возбудимости. а) Порог раздражения - это минимальная сила раздражителя, при которой возникает возбуждение. б) Реобаза - это минимальная сила раздражителя, вызывающая возбуждение при его действии в течение неограниченно долгого времени. На практике порог и реобаза имеют одинаковый смысл. Чем ниже порог раздражения или меньше реобаза, тем выше возбудимость ткани. в) Полезное время - минимальное время действия раздражителя силой в одну реобазу за которое возникает возбуждение. г) Хронаксия - это минимальное время действия раздражителя силой в две реобазы, необходимое для возникнов4. Закон градиента или аккомодации. Реакция ткани на раздражение зависит от его градиента, т.е. чем быстрее нарастает сила раздражителя во времени тем быстрее возникает ответная реакция. При низкой скорости нарастания силы раздражителя растет порог раздражения. Поэтому если сила раздражителя , возрастает очень медленно возбуждения не будет. Это явление называется аккомодацией. Физиологическая лабильность (подвижность) - это большая или меньшая частота реакций, которыми может отвечать ткань на ритмическое раздражение. Чем быстрее восстанавливается ее возбудимость после очередного раздражения, тем Выше ее лабильность. Определение лабильности предложено Н.Е.Введенским. Наибольшая, лабильность у нервов, наименьшая у сердечной мышцы. ения возбуждения. Действие постоянного тока на возбудимые ткани. •возбудимость повышается, а под положительным - анодом снижается. Это называется законом действия постоянного Тока. Изменение возбудимости ткани (например: нерва) под действием постоянного тока в области анода или катода называется физиологическим электротоном. В настоящее время установлено, что под действием отрицательного электрода - катода потенциал мембраны клеток снижается. Это явление называется физическим катэлектротоном, Под положительным - анодом, он возрастает. Возникает физический катэлектртон. Так как, под катодом мембранный потенциал приближается к критическому уровню деполяризации, возбудимость клеток и тканей повышается. Под анодом мембранный потенциал возрастает и удаляется от критического уровня деполяризации, поэтому возбудимость клетки, ткани падает. Следует отметить, что при очень кратковременном действии постоянного тока (1 мсек и менее)МП не успевает измениться, поэтому не изменяется и возбудимость ткани под электродами. Критический уровень деполяризации (Ек)- это та разность потенциалов, которая должна быть достигнута, чтобы местные изменения перешли в пик потенциала действия. Ек - пороговая величин 6 вопрос Фаза деполяризации возникает в результате лавинообразного движения Nа+ внутрь клетки. Этому способствуют две причины: открываются потенциалзависимые Nа+-каналы. В этом случае происходит деполяризация по типу процесса с положительной обратной связью (самоподкрепляющийся процесс). Освобождение натриевых каналов от Са2+. Заряд клеточной мембраны сначала снижается до 0 (это собственно деполяризация), а затем меняется на противоположный (инверсия или овершут). Для характеристики фазы деполяризации вводится понятие реверсии - это та разность потенциалов, на которую потенциал действия превышает потенциал покоя. Р (реверсия)- это то количество мВ на которое произошла перезарядка мембраны. Фаза деполяризации продолжается до достижения электрохимического равновесия по Nа+. Затем наступает следующая фаза. Амплитуда потенциала действия не зависит от силы раздражителя. Она зависит от концентрации Nа+ (как снаружи, так и внутри клетки), от количества натриевых каналов, особенностей натриевой проницаемости. Фаза реполяризации характеризуется: снижением проницаемости клеточной мембраны для Nа+ (Nа-инактивация). Натрий накапливается на наружной поверхности клеточной мембраны; возрастает проницаемость мембраны для К+, в результате повышается выход К+ из клетки с увеличением положительного заряда на мембране; изменение активности Nа+-К+ насоса. Реполяризация - это процесс восстановления заряда мембраны. Но полного восстановления нет, т. к. возникают следовые потенциалы. Гиперполяризация – при частом возбуждении канал не успевает захлопнуться, калий выходит из клетки, мембрана становится более электроотрицателной. Так как в формировании потенциала покоя мембраны участвуют многие ионы, равновесие клетки может нарушаться посредством изменений проводимости различных ионов. Так, например, при дополнительном выходящем токе ионов К+ или при входящем токе ионов Сl может увеличиваться потенциал покоя мембраны, это означает что она гиперполяризуется. Гиперполяризация мембраны - противоположность возбуждения, т.е. определенные химические процессы на постсинаптической мембране могут вызывать торможение нейрона. 7. Потенциал действия – это сдвиг мембранного потенциала, возникающий в ткани при действии порогового и сверхпорогового раздражителя, что сопровождается перезарядкой клеточной мембраны. При действии порогового или сверхпорогового раздражителя изменяется проницаемость клеточной мембраны для ионов в различной степени. Для ионов Na она повышается в 400–500 раз, и градиент нарастает быстро, для ионов К – в 10–15 раз, и градиент развивается медленно. В результате движение ионов Na происходит внутрь клетки, ионы К двигаются из клетки, что приводит к перезарядке клеточной мембраны. Наружная поверхность мембраны несет отрицательный заряд, внутренняя – положительный. Стадия покоя. Эта стадия представлена мембранным потенциалом покоя, который предшествует потенциалу действия. Мембрана во время этой стадии поляризована в связи с наличием отрицательного мембранного потенциала Фаза деполяризации. В это время мембрана внезапно становится высокопроницаемой для ионов натрия, позволяя огромному числу положительно заряженных ионов натрия диффундировать внутрь клетки. Нормальное поляризованное состояние немедленно нейтрализуется поступающими внутрь положительно заряженными ионами натрия, в результате потенциал стремительно нарастает в положительном направлении. Этот процесс называют деполяризацией, Фаза реполяризации. В течение нескольких долей миллисекунды после резкого повышения проницаемости мембраны для ионов натрия, натриевые каналы начинают закрываться, а калиевые — открываться. В результате быстрая диффузия ионов калия наружу восстанавливает нормальный отрицательный мембранный потенциал покоя. Этот процесс называют реполя-ризацией мембраны. Изменения мембранного потенциала,следующие за пиком потэнциала действия,называют СЛЕДОВЫМИ ПОТЕНЦИАЛАМИ. Различают 2 вида следовых потенциалов-следовую деполяризацию и следовую гиперполяризацию.Амплитуда следовых потенциалов обычно не превышает нескольких милливольт,а длительность их у различных волокон составляет от нескольких миллисекунд до десятков и сотен секунд. Механизм проведения возбуждения по нервным волокнам зависит от их типа. Существуют два типа нервных волокон: миелиновые и безмиелиновые. Процессы метаболизма в безмиелиновых волокнах не обеспечивают быструю компенсацию расхода энергии. Распространение возбуждения будет идти с постепенным затуханием – с декрементом. Декрементное поведение возбуждения характерно для низкоорганизованной нервной системы. Возбуждение распространяется за счет малых круговых токов, которые возникают внутрь волокна или в окружающую его жидкость. Между возбужденными и невозбужденными участками возникает разность потенциалов, которая способствует возникновению круговых токов. Ток будет распространяться от «+» заряда к «—». В месте выхода кругового тока повышается проницаемость плазматической мембраны для ионов Na, в результате чего происходит деполяризация мембраны. Между вновь возбужденным участком и соседним невозбужденным вновь возникает разность потенциалов, что приводит к возникновению круговых токов. Возбуждение постепенно охватывает соседние участки осевого цилиндра и так распространяется до конца аксона. В миелиновых волокнах благодаря совершенству метаболизма возбуждение проходит, не затухая, без декремента. За счет большого радиуса нервного волокна, обусловленного миелиновой оболочкой, электрический ток может входить и выходить из волокна только в области перехвата. При нанесения раздражения возникает деполяризация в области перехвата А, соседний перехват В в это время поляризован. Между перехватами возникает разность потенциалов, и появляются круговые токи. За счет круговых токов возбуждаются другие перехваты, при этом возбуждение распространяется сальтаторно, скачкообразно от одного перехвата к другому. Сальтаторный способ распространения возбуждения экономичен, и скорость распространения возбуждения гораздо выше (70—120 м/с), чем по безмиелиновым нервным волокнам (0,5–2 м/с). Возникновению потенциала действия предшествует в точке раздражения мышцы или нерва активные под пороговые изменения мембранного потенциала. Они проявляются в форме локального (местного) ответа. 8. Для локального ответа характерны: 1) зависимость от силы раздражения 2) нарастание постепенно величины ответа. 3) нераспространение по нервному волокну. Первые признаки локального ответа обнаруживаются при действии стимулов составляющих 50-70% пороговой величины. Локальный ответ как и потенциал действия обусловлен повышением натриевой проницаемости. Однако это повышение было недостаточно, чтобы вызвать потенциал действия. Потенциал действия возникает когда деполяризация мембраны достигнет критического уровня. Но локальный ответ важен. Он подготавливает ткани к последующим воздействиям. Сравнительная характеристика локального потенциала и ПД
Повышение возбудимости клетки во время локального потенциала объясняется тем, что мембрана оказывается частично деполяризованной. Если КУД остается на постоянном уровне, то для его достижения требуется гораздо меньший раздражитель. Амплитуда ПД не зависит от силы раздражителя, потому что он возникает вследствие регенеративных процессов. При стимуляции рецептора внешним воздействием, к которому чувствителен рецептор , происходит деполяризация сомы , которая исчезает с прекращением стимуляции. Деполяризация называется рецепторным потенциалом (или генераторным потенциалом ). Длительность его соответствует длительности стимула, а его амплитуда возрастает с увеличением интенсивности стимуляции, таким образом, он является отражением стимула, а не ответом по типу "все или ничего", как потенциал действия . Рецепторный потенциал обусловлен повышением Na+ - проводимости мембраны дендритов , в результате чего вход ионов натрия создает деполяризующий рецепторный потенциал, который электротонически распространяется к соме . Эта первичная трансформация стимула в рецепторный потенциал называется преобразованием, а рецептор, таким образом, является преобразователем, датчиком. Пороговая Сила тока увеличивается при уменьшении крутизны его нарастания,а при некоторой минимальной крутизне ответы на раздражение исчезают.Это я вление принято обозначать термином « аккомодация » В основе аккомодации лежат инактивация натриевой и повышение калиевой проводимостей,развивающиеся во время медленно нарастающей деполяризации мембраны.Аккомодация различных нервных волокон варьирует в широких пределах, но у двигательных неравных волокон скорость аккомодации,как правило,значительно выше,чем у чувствительных волокон. Кратковременное снижение возбудимости (См. Возбудимость) нервной и мышечной тканей непосредственно вслед за потенциалом действия (См. Потенциал действия). Р. обнаруживается при стимуляции нервов и мышц парными электрическими импульсами. Если сила 1-го импульса достаточна для возникновения потенциала действия, ответ на 2-й будет зависеть от длительности паузы между импульсами. При очень коротком интервале ответ на 2-й импульс отсутствует, как бы ни увеличивалась интенсивность стимуляции (абсолютный Рефрактерный период). Удлинение интервала приводит к тому, что 2-й импульс начинает вызывать ответ, но меньший по амплитуде, чем 1-й импульс (в опытах на нервных стволах, состоящих из большого числа параллельных нервных проводников), либо для возникновения ответа на 2-й импульс необходимо увеличить силу раздражающего тока (в опытах на одиночных нервных волокнах). Период сниженной возбудимости нервной или мышечной клетки называется относительным рефракторным периодом. За ним следует супернормальный период, или фаза экзальтации (См. Экзальтация), т. е. фаза повышенной возбудимости, сменяющаяся периодом несколько сниженной возбудимости — субнормальным периодом. В основе наблюдаемых колебаний возбудимости лежит изменение проницаемости биологических мембран (См. Проницаемость биологических мембран), сопровождающее возникновение потенциала действия (см. Биоэлектрические потенциалы). Длительность каждого периода определяется кинетикой этих процессов в данной ткани. В быстропроводящих нервных волокнах Р. длится не более 3—5 мсек, в мышце сердца период изменений возбудимости занимает до 500 мсек. Р. — один из факторов, ограничивающих частоту воспроизведения биологических сигналов, их суммацию и скорость проведения. При изменении температуры или действии некоторых лекарственных веществ длительность рефракторных периодов может меняться, чем пользуются для управления возбудимостью ткани, например сердечной мышцы: удлинение относительного рефрактерного периода приводит к снижению частоты сердечных сокращений и устранению нарушений ритма работы сердца. 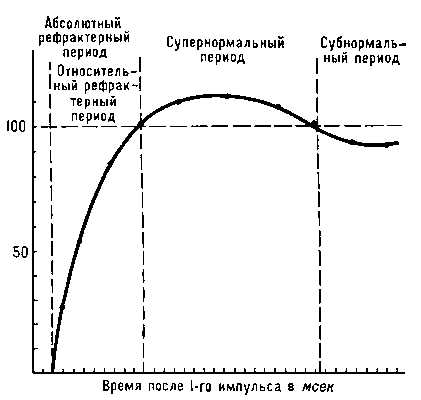 Нервно мышечная физиология 1)Особенности проведения возбуждения по миелинизированным и немиелинезированным нервным волокнам. Скорость проведения по нервным и мышечным волокнам. Классификация нервных волоком по скорости проведения возбуждения. Законы проведения возбуждения по НВ. Нервные волокна: МИЕЛИНИЗИРОВАННЫЕ – Большая часть аксона покрыта жировой оболочкой как изолятором. Миелиновые волокна состоят из осевого цилиндра, покрытого миелиновой оболочкой, образованной шванновскими клетками. Осевой цилиндр имеет мембрану и оксоплазму. Миелиновая оболочка состоит на 80 % из липидов, обладающих высоким омическим сопротивлением, и на 20 % из белка. Миелиновая оболочка прерывается и оставляет открытыми участки осевого цилиндра, которые называются узловыми перехватами (перехваты Ранвье).. Контакт с вн. средой только в перехватах Ранвье. Длина участков между перехватами различна и зависит от толщины нервного волокна: чем оно толще, тем длиннее расстояние между перехватами. НЕМИЕЛИНИЗИРОВАННЫЕ- мембрана напрямую контактирует с вн. Средой. Обмен ионов между внутри и внеклеточной средой проходит в любой точке. Немиелинизированными являются волокна вегетативных нервов. Скорость проведения возбуждения и потенциалов действия, сопровождающих возбуждение в миелинизированных нервных волокнах выше, чем в немиелинизированных волокнах того же диаметра. Это обусловлено сальтаторным проведением возбуждения - скачкообразным распространением возбуждения по возбудимым мембранам миелинизированных нервных волокон. Различия проведения возбуждения: - только перехваты Ранвье за счёт большой плотности Na+ каналов генерируют максимальный по амплитуде ПД - в мякотном волокне возбуждение передаётся сальтаторно (скачкообразно) и с большей скоростью - передача возбуждения в мякотных волокнах экономична, не требует энергии для работы Na\K насоса. - проведение возбуждения по мякотным волокнам более надежно. Про повреждении нескольких перехватов Ранвье возбуждение может проводиться на следующие перехваты (перескакивая) Скорость проведения по нервным и мышечным волокнам. Механизм проведения возбуждения по нервным волокнам зависит от их типа. Существуют два типа нервных волокон: миелиновые и безмиелиновые. Процессы метаболизма в безмиелиновых волокнах не обеспечивают быструю компенсацию расхода энергии. Распространение возбуждения будет идти с постепенным затуханием – с декрементом. Декрементное поведение возбуждения характерно для низкоорганизованной нервной системы. Возбуждение распространяется за счет малых круговых токов, которые возникают внутрь волокна или в окружающую его жидкость. Между возбужденными и невозбужденными участками возникает разность потенциалов, которая способствует возникновению круговых токов. Ток будет распространяться от «+» заряда к «—». В месте выхода кругового тока повышается проницаемость плазматической мембраны для ионов Na, в результате чего происходит деполяризация мембраны. Между вновь возбужденным участком и соседним невозбужденным вновь возникает разность потенциалов, что приводит к возникновению круговых токов. Возбуждение постепенно охватывает соседние участки осевого цилиндра и так распространяется до конца аксона. В миелиновых волокнах благодаря совершенству метаболизма возбуждение проходит, не затухая, без декремента. За счет большого радиуса нервного волокна, обусловленного миелиновой оболочкой, электрический ток может входить и выходить из волокна только в области перехвата. При нанесения раздражения возникает деполяризация в области перехвата А, соседний перехват В в это время поляризован. Между перехватами возникает разность потенциалов, и появляются круговые токи. За счет круговых токов возбуждаются другие перехваты, при этом возбуждение распространяется сальтаторно, скачкообразно от одного перехвата к другому. Сальтаторный способ распространения возбуждения экономичен, и скорость распространения возбуждения гораздо выше (70—120 м/с), чем по безмиелиновым нервным волокнам (0,5–2 м/с). - миелиновые волокна- имеют оболочку обладающую высоким сопротивлением, электрогенные свойства только в перехватах Ранвье. Под действием раздражителя возбуждение возникает в ближайшем перехвате Ранвье. Соседний перехват в состоянии поляризации. Возникающий ток вызывает деполяризацию соседнего перехвата. В перехватах Ранвье высокая плотность Nа-каналов, поэтому в каждом следующем перехвате возникает чуть больший (по амплитуде) потенциал действия, за счет этого возбуждение распространяется без декремента и может перескакивать через несколько перехватов. Это сальтаторная теория Тасаки. Доказательство теории - в нервное волокно вводили препараты, блокирующие несколько перехватов, но проведение возбуждения регистрировалось и после этого. Это высоко надежный и выгодный способ, т. к. устраняются небольшие повреждения, увеличивается скорость проведения возбуждения, уменьшаются энергетические затраты - безмиелиновые волокна- поверхность обладает электрогенными свойствами на всем протяжении. Поэтому малые круговые токи возникают на расстоянии в несколько микрометров. Возбуждение имеет вид постоянно бегущей волны. Классификация нервных волоком по скорости проведения возбуждения.
Законы проведения возбуждения по НВ.
Проведение импульсов по нервному волокну возможно лишь в том случае, если не нарушена его целостность. При нарушении физиологических свойств нервного волокна путем охлаждения, применения различных наркотических средств, сдавливания, а также порезами и повреждениями анатомической целостности проведение нервного импульса по нему будет невозможно.
Существует ряд особенностей распространения возбуждения в периферических, мякотных и безмякотных нервных волокнах. В периферических нервных волокнах возбуждение передается только вдоль нервного волокна, но не передается на соседние, которые находятся в одном и том же нервном стволе. В мякотных нервных волокнах роль изолятора выполняет миелиновая оболочка. За счет миелина увеличивается удельное сопротивление и происходит уменьшение электрической емкости оболочки. В безмякотных нервных волокнах возбуждение передается изолированно. Это объясняется тем, что сопротивление жидкости, которая заполняет межклеточные щели, значительно ниже сопротивления мембраны нервных волокон. Поэтому ток, возникающий между деполяризованным участком и неполяризованным, проходит по межклеточным щелям и не заходит при этом в соседние нервные волокна.
2)Виды мышечных волокон. Иннервация скелетной мышцы. Нейромоторная ( двигательная) единица. Нейротрофический контроль свойств скелетной мышцы. Виды мышц: -скелетные: произвольные, поперечно-полосатые -сердечные: непроизвольные, поперечно-полосатые -гладкие: непроизвольные, нет поперечной исчерченности Типы мышечных волокон: 1)Фазные волокна: - медленные (красные) – много миоглобина ( связывает О2) и митохондрий; поддержание позы; утомление наступает очень медленно, быстро проходит. - быстрые (окислительного типа) – много митохондрий; образует АТФ путём окислительного фосфорилирования; быстрые энергичные движения без заметного утомления. - быстрые (гликолитического типа, «белые») – мало митохондрий, миоглобина нет; АТФ образуется за счёт гликолиза; быстрое и сильное сокращение, но сравнительно быстро утомляется. 2) Тонические - сокращение и расслабление происходят медленно; входят в состав наружных мышц глаза. Иннервация скелетной мышцы: Соматическая нервная система (произвольная); один двигательный нейрон (тело в ЦНС); осуществляет возбуждение. Двигательную (эфферентную) иннервацию скелетные мышцы туловища и конечностей получают от мотонейронов передних рогов спинного мозга, а мышцы лица и головы - от двигательных нейронов определенных черепных нервов. При этом к каждому мышечному волокну подходит или ответвление от аксона мотонейрона, или же весь аксон. В мышцах, обеспечивающих тонкие координированные движения (мышцы кистей, предплечий, шеи), каждое мышечное волокно иннервируется одним мотонейроном. В мышцах, обеспечивающих преимущественно поддержание позы, десятки и даже сотни мышечных волокон получают двигательную иннервацию от одного мотонейрона, посредством разветвления его аксона. Двигательное нервное волокно, подойдя к мышечному волокну, проникает под эндомизий и базальную пластинку и распадается на терминали, которые вместе с прилежащим специфическим участком миосимпласта образуют аксо-мышечный синапс или моторную бляшку. Под влиянием нервного импульса волна деполяризации с нервного окончания передается на плазмолемму миосимпласта, распространяется далее по Т-канальцам и в области триад передается на терминальные цистерны саркоплазматической сети, обуславливая выход ионов кальция и начало процесса сокращения мышечного волокна. Нейромоторная единица - совокупность нейрона и группы мышечных волокон, иннервируемых аксоном этого нейрона. В состав нейромоторной единицы входят: нервная клетка - в основном мотонейроны, тела которых лежат в передних рогах спинного мозга; аксон мотонейрона - миелиновые волокна; группа мышечных волокон - в зависимости от вида деятельности количества волокон различно. Если тонкая работа 2-4, если грубая - до нескольких тысяч. Виды нейромоторных единиц По характеру возбуждения, возникающего в мышечных волокнах все нейромоторные единицы делятся на 2 группы. Фазные нейромоторные единицы- образуются альфа-мотонейронами. Это самые крупные мотонейроны (диаметр 10-20 мкм). Скорость проведения возбуждения по отростку этого аксона - 120 м/с. Аксон обеспечивает одиночную иннервацию, образуя на каждом волокне 1-2 синапса. В каждом синапсе при возбуждении возникает достаточный потенциал концевой пластинки, который обеспечивает возникновение потенциала действия в мышце (импульсное возбуждение). Лучше работают под действием одиночного возбуждения (импульса), при котором возникает распространяющееся возбуждение. Среди фазных нейромоторных единиц выделяют быстрые и медленные. Быстрые - продолжительность потенциала действия в 2 раза меньше, чем в медленных. Волна сокращения в 5 раз меньше, чем в медленных. Скорость распространения возбуждения в быстрых нейромоторных единицах в 2 раза больше, чем в медленных. Таким образом, быстрые фазные нейромоторные единицы обеспечивают динамическую работу, когда быстрое сокращение сменяется быстрым расслаблением. Для этого вида нейромоторных единиц характерно анаэробное образование энергии. Эти мышечные волокна практически не содержат миоглобина - светлые, белые мышцы; Медленные- обеспечивают в основном статическую работу, медленное, длительное сокращение мышц. Основной поставщик энергии окислительно-восстановительные процессы. Содержит миоглобин, который депонирует кислород. По цвету темные, красные мышцы. Тонические нейромоторные единицы- образуются гамма-мотонейронами передних рогов спинного мозга (самые мелкие, диаметр 4-6 мкм). Скорость проведения импульса по отросткам этих нейронов - 30 м/с, в синапсах возникает местный потенциал концевой пластинки (местное возбуждение). Аксоны тонических нейронов обеспечивают множественную иннервацию, т. е. на каждом мышечном волокне образуются несколько десятков синапсов и за счет суммации местных потенциалов и возникает потенциал действия. Возбуждение тонической нейромоторной единицы происходит под влиянием серии импульсов (частота около 10 Гц). Тонические нейромоторные единицы обеспечивают медленное сокращение мышц, участвуют в возникновении тонуса. Скелетные мышцы образуются различными мышечными волокнами, которые входят в состав фазных и тонических нейромоторных единиц. Попеременное включение нейромоторных единиц обеспечивает изменение функционального состояния мышцы. В состоянии покоя работают тонические нейромоторные единицы, в состоянии активности - фазные нейромоторные единицы. Кроме двигательной иннервации присутствует вегетативная. Все скелетные мышцы получают импульсы из симпатической нервной системы, которая регулирует обменные процессы. Нейротрофический контроль свойств скелетной мышцы. 3)Особенности проведения возбуждения в нервно-мышечном синапсе. Везикулярная гипотеза. Квантовая секреция медиатора. Механизм слияния пресинаптических везикул с постсинаптической мембраной (роль пептидов нервного окончания и мембраны везикул). Спонтанная квантовая секреция. Синапс – специализированный межклеточный контакт ,обеспечивающий передачу информации между двумя возбудимыми клетками. Cтруктура синапса: 1) пресинаптическая мембрана (электрогенная мембрана в терминале аксона, образует синапс на мышечной клетке); 2) постсинаптическая мембрана (электрогенная мембрана иннервируемой клетки, на которой образован синапс); 3) синаптическая щель (пространство между пресинаптической и постсинаптической мембраной, заполнена жидкостью, которая по составу напоминает плазму крови). 2. Механизмы передачи возбуждения в синапсах на примере мионеврального синапса (Полный ответ) Мионевральный (нервно-мышечный) синапс – образован аксоном мотонейрона и мышечной клеткой. Нервный импульс возникает в тригерной зоне нейрона, по аксону направляется к иннервируемой мышце, достигает терминали аксона и при этом деполяризует пресинаптическую мембрану. После этого открываются натриевые и кальциевые каналы, и ионы Ca из среды, окружающей синапс, входят внутрь терминали аксона. При этом процессе броуновское движение везикул упорядочивается по направления к пресинаптической мембране. Ионы Ca стимулируют движение везикул. Достигая пресинаптическую мембрану, везикулы разрываются, и освобождается ацетилхолин (4 иона Ca высвобождают 1 квант ацетилхолина). Синаптическая щель заполнена жидкостью, которая по составу напоминает плазму крови, через нее происходит диффузия АХ с пресинаптической мембраны на постсинаптическую, но ее скорость очень мала. Кроме того, диффузия возможна еще и по фиброзным нитям, которые находятся в синаптической щели. После диффузии АХ начинает взаимодействовать с хеморецепторами (ХР) и холинэстеразой (ХЭ), которые находятся на постсинаптической мембране. Холинорецептор выполняет рецепторную функцию, а холинэстераза выполняет ферментативную функцию. На постсинаптической мембране они расположены следующим образом: ХР + АХ = МПКП – миниатюрные потенциалы концевой пластины. Затем происходит суммация МПКП. В результате суммации образуется ВПСП – возбуждающий постсинаптический потенциал. Постсинаптическая мембрана за счет ВПСП заряжается отрицательно, а на участке, где нет синапса (мышечного волокна), заряд положительный. Возникает разность потенциалов, образуется потенциал действия, который перемещается по проводящей системе мышечного волокна. ХЭ + АХ = разрушение АХ до холина и уксусной кислоты. В состоянии относительного физиологического покоя синапс находятся в фоновой биоэлектрической активности. Ее значение заключается в том, что она повышает готовность синапса к проведению нервного импульса. В состоянии покоя 1–2 пузырька в терминале аксона могут случайно подойти к пресинаптической мембране, в результате чего вступят с ней в контакт. Везикула при контакте с пресинаптической мембраной лопается, и ее содержимое в виде 1 кванта АХ поступает в синаптическую щель, попадая при этом на постсинаптическую мембрану, где будет образовываться МПКН. Проведение возбуждения в НМС ( по лекции) 1.ПД нервного окончания 2.активания потенциал зависимых Са2+ каналов, вход ионов Са2+ в нервное окончание 3.освобождение ацетилхолина путем экзоцитоза везикул в синаптическую щель 4.активания постсинаптических н-холинорецепторов 5.Вход Na+ возникновение потенциала концевой пластинки, затем потенциала действия, вход Са2+ 6.сокращение мышцы Везикулярная гипотеза- 1) нейромедиатор выделяется из нервного окончания мультимолекулярными порчиями – квантами. 2)Один квант находится в одной синаптической везикуле. 3) везикулярный медиатор осовбождается путем слияния мембраны везикулы с пресинаптической мембраной (экзоцитоз) Кватовая секреция медиатора – Са2+ зависимый процесс. Повышение Са2+ в аксоплазе от 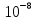 (покой) до (покой) до 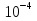 (активация) вызывает массивное освобождение квантов медиатора (экзоцитоз). Основной пусть выхода Са2+ в терминаль – потенциал зависимые Са2+ каналы с пресинаптической мембране, открываемые при деполяризации. Блокада этих каналов ( например кадмием) устраняет эффект стимуляции. (активация) вызывает массивное освобождение квантов медиатора (экзоцитоз). Основной пусть выхода Са2+ в терминаль – потенциал зависимые Са2+ каналы с пресинаптической мембране, открываемые при деполяризации. Блокада этих каналов ( например кадмием) устраняет эффект стимуляции. Последовательность явлений: 1.Приход ПД в терминаль 2.Активания Са2+ каналов 3.Быстрый локальный рост Са2+ 4.Вызванное Са2+ повышение вероятности экзоцитоза медиатора 5.этот процесс занимает 0,2-0,5 мс что соответствует синаптической задержке между стимулом и постсинаптическим ответом Механизм слияния пресинаптических везикул с постсинаптической мембраной (роль пептидов нервного окончания и мембраны везикул). В пресинаптическом нервном окончании в синаптических везикулах аккумулируются медиаторы (трансмиттеры). Потенциал действия, достигая нервного окончания, деполяризует его мембрану. Деполяризация вызывает открытие кальциевых каналов (насосов), по которым в окончание входит ток ионов Са2+. Ионы Са2+ вызывают процесс слияния везикул с пресинаптической мембраной с последующим открытием их и выбрасыванием своего содержимого (медиаторов) в синаптическую щель. Синаптические везикулы фиксируются большей частью на цитоскелете посредством протеина синапсина, локализованного на цитоплазматической поверхности каждой везикулы, к протеину спектрину, расположенному на волокнах F-актина цитоскелета, и образуют тем самым трансмиттерный резервуар. Меньшая часть везикул связана специфическими протеинами с внутренней стороной пресинаптической мембраны. Это взаимодействие осуществляется посредством белка мембраны везикулы синаптобревина и белка пресинаптической мембраны синтаксина . Именно эти везикулы непосредственно поставляют трансмиттер для очередного выброса. Если потенциал действия достиг пресинаптической области и в пресинаптическом окончании концентрация Са2+ поднялась до необходимого уровня, то происходят два процесса. Во-первых, на уже связанных с пресинаптической мембраной везикулах, которые, по существу, лежат на ней, Са2+ связывается с протеином, входящим в состав их мембраны, - синаптотагмином. Это приводит к тому, что мембрана везикулы раскрывается. Одновременно комплекс полипептида синаптофизина сливается с неидентифицированными протеинами пресинаптической мембраны. При этом возникает пора, через которую осуществляется регулируемый экзоцитоз, т.е. секреция трансмиттера в синаптическую щель, причем еще один протеин везикулы, rab3A , регулирует этот процесс. В одной везикуле сосредоточено примерно 6000-8000 молекул трансмиттера; это наименьшее количество трансмиттера, освобожденного в синаптическую щель, которое называется "один квант трансмиттера ". В совокупности локальная концентрация трансмиттера в синаптической щели после его освобождения относительно высока и находится в миллимолярном диапазоне. Таким образом, главную роль для процесса выброса трансмиттера по типу экзоцитоза играет не деполяризация окончания, а именно входящий ток ионов Са2+. Ионы Са2+ служат при этом не для дополнительной деполяризации, а в качестве вещества-посредника (вторичного мессенджера), которое запускает механизм слияния везикул. Повышение концентрации экстрацеллюлярного Са2+ повышает входящий ток ионов Са2+ и, тем самым, увеличивает освобождение трансмиттера. Наоборот, искусственное повышение концентрации экстрацеллюлярного Mg2+ посредством замещения им ионов Са2+ ведет к снижению входящего тока Са2+ и, тем самым, уменьшению освобождения трансмиттера. Небольшие G-белки , вероятно, также участвуют в управлении везикулярным экзоцитозом. После прекращения пресинаптического потенциала действия ионы Са2+ удаляются из пресинаптической области посредством активного ионного транспорта с участием Са2+-АТФазы и 3Na+/Ca2+-обменника. Спонтанная квантовая секреция. Медиатор секретируется мультимолекулярными пакетами-квантами. Квант соответствует содержимому одного синаптического пузырька. МПКП – миниатюрный потенциал концевой пластины – результат постсинаптического действия одного кванта медиатора. МПКП: - возникают спонтанно вне зависимости от стимуляции нерва - имеют тот же временный ход что и вызванные ответы - амплитула МПКП зависит от состояния АХ-рецепторов и конц. АХ в везикуле - частота МПКП зависит от состояния терминали |