Общие рекомендации
 Скачать 4.72 Mb. Скачать 4.72 Mb.
|

КЛЕТКА — ЭЛЕМЕНТАРНАЯ ЕДИНИЦА ЖИВОГОКлетка представляет собой обособленную, наименьшую по размерам структуру, которой присуща вся совокупность свойств жизни и которая может в подходящих условиях окружающей среды поддерживать эти свойства в самой себе, а также передавать их в ряду поколений. Клетка, таким образом, несет полную характеристику жизни. Вне клетки не существует настоящей жизнедеятельности. Поэтому в природе планеты ей принадлежит роль элементарной структурной, функциональной и генетической единицы. Это означает, что клетка составляет основу строения, жизнедеятельности и развития всех живых форм — одноклеточных, многоклеточных и даже неклеточных. Благодаря заложенным в ней механизмам клетка обеспечивает обмен веществ, использование биологической информации, размножение, свойства наследственности и изменчивости, обусловливая тем самым присущие органическому миру качества единства и разнообразия. Занимая в мире живых существ положение элементарной единицы, клетка отличается сложным строением. При этом определенные черты обнаруживаются во всех без исключения клетках, характеризуя наиболее важные стороны клеточной организации как таковой. 2.1. КЛЕТОЧНАЯ ТЕОРИЯКлеточная теория сформулирована немецким исследователем, зоологом Т. Шванном (1839). Поскольку при создании этой теории Шванн широко пользовался работами ботаника М. Шлейдена, последнего по праву считают соавтором клеточной теории. Исходя из предположения о схожести (гомологичности) растительных и животных клеток, доказываемой одинаковым механизмом их возникновения, Шванн обобщил многочисленные данные в виде теории, согласно которой клетки являются структурной и функциональной основой живых существ. В конце XIX столетия немецкий патолог Р. Вирхов на основе новых фактов пересмотрел клеточную теорию. Ему принадлежит вывод о том, что клетка может возникнуть лишь из предсуществующей клетки. Им также создана вызвавшая критику концепция «клеточного государства», согласно которой многоклеточный организм состоит из относительно самостоятельных единиц (клеток), поставленных в своей жизнедеятельности в тесную зависимость друг от друга. Клеточная теория в современном виде включает три главных положения. Первое положение соотносит клетку с живой природой планеты в целом. Оно утверждает, что жизнь, какие бы сложные или простые (например, вирусы) формы она ни принимала, в ее структурном, функциональном и генетическом отношении обеспечивается в конечном итоге только клеткой. Выдающаяся роль клетки как первоисточника жизни обусловливается тем, что именно она является биологической единицей, с помощью которой происходит извлечение из внешней среды, превращение и использование организмами энергии и веществ. Непосредственно в клетке сохраняется и используется биологическая информация. Второе положение указывает, что в настоящих условиях единственным способом возникновения новых клеток является деление предсуществующих клеток. В обосновании клеточной природы жизни на Земле тезису о единообразии путей возникновения клеток принадлежит особая роль. Именно этот тезис был использован М. Шлейденом и Т. Шванном для обоснования представления о гомологии разных типов клеток. Современная биология расширила круг доказательств этому. Независимо от индивидуальных структурно-функциональных особенностей все клетки одинаковым образом: а) хранят биологическую информацию, б) редуплицируют генетический материал с целью его передачи в ряду поколений, в) используют информацию для осуществления своих функций на основе синтеза белка, г) хранят и переносят энергию, д) превращают энергию в работу, е) регулируют обмен веществ. Третье положение клеточной теории соотносит клетку с многоклеточными организмами, для которых характерен принцип целостности и системной организации. Для системы свойственно наличие новых качеств благодаря взаимному влиянию и взаимодействию единиц, составляющих эту систему. Структурно-функциональными единицами многоклеточных существ являются клетки. Вместе с тем многоклеточный организм характеризуется рядом особых свойств, которые нельзя свести к свойствам и качествам отдельных клеток. В третьем положении клеточной теории мы встречаемся с проблемой соотношения части и целого. Системный подход как научное направление используется в биологических исследованиях с начала прошлого столетия. Системный характер организации и функционирования свойствен не только организму, но и другим главным биологическим образованиям — геному, клетке, популяции, биогеоценозу, биосфере. 9.Биологическая мембрана, молекулярная организация и функции. Транспорт веществ через мембрану (модели транспорта). Клетка – система мембран, отграничивающих участки внутриклеточного пространства. Мембраны участвуют в разных процессах. Мембраны нервных клеток – генерация нервного импульса, мембраны ЖКТ – всасывание и переваривание пищи, клеточные мембраны скелетных мышц и клеток миокарда – расслабление и сокращение, мембраны клеток органов чувств – преобразование одного вида раздражения в другой. Белки природных мембран плохо растворимы в воде, образуют комплексы с липидами. Функции: рецепторная, структурная, транспортная, каталитическая (большинство белков – ферменты – иммуноглобулины – белки с наибольшей активностью). Жидкостно-мозаическап модель строения мембраны (бислой липидов, белки – периферические, погруженные, интегральные). Транспорт веществ не всегда происходит путем диффузии или градиента. Существуют транспортные белки. АТФ---АДФ+Ф Активный транспорт – перенос веществ через мембрану с затратой АТФ и при участии транспортных белков. Активный сопряженный транспорт (одни и те же белки – несколько веществ). Могут быть 2 периферических белка, могут идти вещества по каналу, 2-3 переносчика, транспорт может быть несопряженным. Бывает экзцитоз (пиноцитоз и фагоцитоз). Существование обменной диффузии (при помощи градиента концентрации), В эукариотических клетках эндоплазматическая мембрана составляет единое целое с внутриклеточными мембранами (цитоплазматические мембраны). Цитоплазматические мембраны образуют канал со сложной сетью переплетающихся каналов и пузырьков (1959 Картер) каналы ЭПР – временные коммуникационные системы, участвующие в перемещении пузырьков из клетки наружу. Мембраны гЭПР имеют трубчатые структуры (более молодой эволюционный тип). Биологическая роль: гидролиз фосфолипидов, синтез стероидных гормонов, синтез липидов и т.д. как и плазматическая, цитоплазматическая мембрана переходит в ядерную и комплекс Гольджи(1898 ит. Гольджи) главная особенность – отсутствие рибосом. Это плоскопараллельные цистерны, пузырьки. Комплекс специализирован для синтеза белков. Собственные клеточные белки, секреторные белки синтезируются в большинстве клеток. Большинство белков сложные, имеют углеводную и белковую часть – гликопротеины и протеогликаны. Комплекс Гольджи участвует в синтезе сложных сахаров из простых моносахаридов, которые затем связываются с белками, пришедшими в комплекс, возникают гликопротеины, образовавшись, они отшнуровываются, происходит экзоцитоз. В комплексе Гольджи образуются лизосомы. Комплекс Гольджи поставляет необходимый материал для образования борозд дробления в животной клетке. Высокая упорядоченность внутреннего содержимого эукариотиче-ской клетки достигается путем компартментации ее объема — подразделения на «ячейки», отличающиеся деталями химического (ферментного) состава. Компартментация (рис. 2.3) способствует пространственному разделению веществ и процессов в клетке. Отдельный компартмент представлен органеллой (лизосома) или ее частью (пространство, отграниченное внутренней мембраной митохондрии). 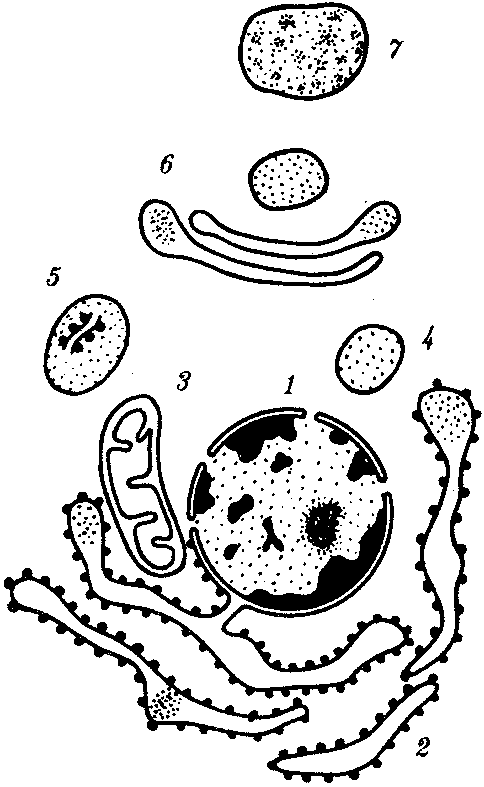 Рис. 2.3. Компартментация объема клетки с помощью мембран: 1—ядро, 2—шероховатая цитоплазматическая есть, 3—митохондрия, 4—транспортный цитоплазматический пузырек, 5—лизосома, 6—пластинчатый комплекс, 7 — гранула секрета 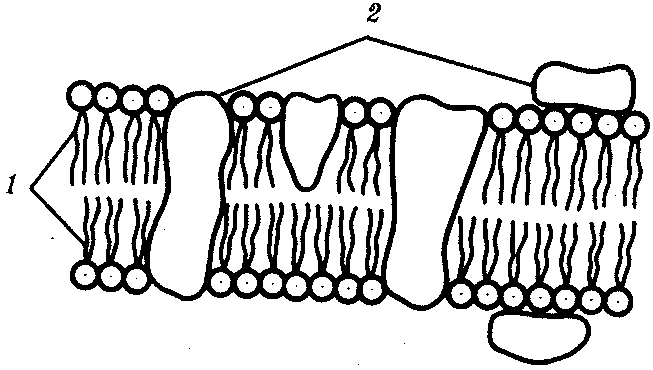 Рис. 2.4. Молекулярная организация биологической мембраны: 1 — бимолекулярный слой липидов, 2 — белки Предложено несколько схем взаимоотношения в мембране основных химических компонентов — белков и липидов, а также веществ, размещаемых на мембранной поверхности. В настоящее время принята точка зрения, согласно которой мембрана составлена из бимолекулярного слоя липидов. Гидрофобные участки их молекул повернуты друг к другу, а гидрофильные — находятся на поверхности слоя. Разнообразные белковые молекулы встроены в этот слой или размещены на его поверхностях (рис. 2.4). Благодаря компартментации клеточного объема в эукариотической клетке наблюдается разделение функций между разными структурами. Одновременно различные структуры закономерно взаимодействуют друг с другом. 10.Ядро. Строение и функции. В клетке выделяют ядро и цитоплазму. Клеточное ядро состоит из оболочки, ядерного сока, ядрышка и хроматина. Функциональная роль ядерной оболочки заключается в обособлении генетического материала (хромосом) эукариотической клетки от цитоплазмы с присущими ей многочисленными метаболическими реакциями, а также регуляции двусторонних взаимодействий ядра и цитоплазмы. Ядерная оболочка состоит из двух мембран, разделенных околоядерным (перинуклеарным) пространством. Последнее может сообщаться с канальцами цитоплазматической сети. Ядерная оболочка пронизана порожу диаметром 80—90 нм. Область поры или поровый комплекс с диаметром около 120 нм имеет определенное строение, что указывает на сложный механизм регуляции ядерно-цитоплазматических перемещений веществ и структур. Количество пор зависит от функционального состояния клетки. Чем выше синтетическая активность в клетке, тем больше их число. Подсчитано, что у низших позвоночных животных в эритробластах, где интенсивно образуется и накапливается гемоглобин, на 1 мкм2 ядерной оболочки приходится около 30 пор. В зрелых эритроцитах названных животных, сохраняющих ядра, на 1 мк»г оболочки остается до пяти пор, т.е. в 6 раз меньше. В области перового комплекса начинается так называемая плотная пластинка — белковый слой, подстилающий на всем протяжении внутреннюю мембрану ядерной оболочки. Эта структура выполняет прежде всего опорную функцию, так как при ее наличии форма ядра сохраняется даже в случае разрушения обеих мембран ядерной оболочки. Предполагают также, что закономерная связь с веществом плотной пластинки способствует упорядоченному расположению хромосом в интерфазном ядре. Основу ядерного сока, или матрикса, составляют белки. Ядерный сок образует внутреннюю среду ядра, в связи с чем он играет важную роль в обеспечении нормального функционирования генетического материала. В составе ядерного сока присутствуют нитчатые, или фибриллярные, белки, с которыми связано выполнение опорной функции: в матриксе находятся также первичные продукты транскрипции генетической информации — гетероядерные РНК (гя-РНК), которые здесь же подвергаются процессингу, превращаясь в м-РНК (см. 3.4.3.2). Ядрышко представляет собой структуру, в которой происходит образование и созревание рибосомальных РНК (рРНК). Гены рРНК занимают определенные участки (в зависимости от вида животного) одной или нескольких хромосом (у человека 13—15 и 21—22 пары) — ядрышковые организаторы, в области которых и образуются ядрышки. Такие участки в метафазных хромосомах выглядят как сужения и называются вторичными перетяжками. С помощью электронного микроскопа в ядрышке выявляют нитчатый и зернистый компоненты. Нитчатый (фибриллярный) компонент представлен комплексами белка и гигантских молекул РНК-предшественниц, из которых затем образуются более мелкие молекулы зрелых рРНК. В процессе созревания фибриллы преобразуются в рибонуклеопротеиновые зерна (гранулы), которыми представлен зернистый компонент. Хроматиновые структуры в виде глыбок, рассеянных в нуклеоплазме, являются интерфазной формой существования хромосом клетки (см. 3.5.2.2). 11.Цитоплазма. Органеллы общего значения и специальные, их строение и функции. В цитоплазме различают основное вещество (матрикс, гиалоплазма), включения и органеллы. Основное вещество цитоплазмы заполняет пространство между плазмалеммой, ядерной оболочкой и другими внутриклеточными структурами. Обычный электронный микроскоп не выявляет в нем какой-либо внутренней организации. Белковый состав гиалоплазмы разнообразен. Важнейшие из белков представлены ферментами гаиколиза, обмена Сахаров, азотистых оснований, аминокислот и липидов. Ряд белков гиалоплазмы служит субъединицами, из которых происходит сборка таких структур, как микротрубочки. Основное вещество цитоплазмы образует истинную внутреннюю среду клетки, которая объединяет все внутриклеточные структуры и обеспечивает взаимодействие их друг с другом. Выполнение матриксом объединяющей, а также каркасной функции может быть связано с выявляемой с помощью сверхмощного электронного микроскопа микротрабекулярной сети, образованной тонкими фибриллами толщиной 2—3 нм и пронизывающей всю цитоплазму. Через гиалоплазму осуществляется значительный объем внутриклеточных перемещений веществ и структур. Основное вещество цитоплазмы следует рассматривать так же, как сложную коллоидную систему, способную переходить из золеобразного (жидкого) состояния в гелеобразное. В процессе таких переходов совершается работа. О функциональном значении таких переходов см. разд. 2.3.8. Включениями (рис. 2.5) называют относительно непостоянные компоненты цитоплазмы, которые служат запасными питательными веществами (жир, гликоген), продуктами, подлежащими выведению из клетки (гранулы секрета), балластными веществами (некоторые пигменты). Органеллы — это постоянные структуры цитоплазмы, выполняющие в клетке жизненно важные функции. Выделяют органеллы общего значения и специальные. Последние в значительном количестве присутствуют в клетках, специализированных к выполнению определенной функции, но в незначительном количестве могут встречаться и в других типах клеток. К ним относят, например, микроворсинки всасывающей поверхности эпителиальной клетки кишечника, реснички эпителия трахеи и бронхов, синаптические пузырьки, транспортирующие вещества — переносчики нервного возбуждения с одной нервной клетки на другую или клетку рабочего органа, миофибриллы, от которых зависит сокращение мышцы. Детальное рассмотрение специальных органелл входит в задачу курса гистологии. К органеллам общего значения относят элементы канальцевой и вакуолярной системы в виде шероховатой и гладкой цитоплазматической сети, пластинчатый комплекс, митохондрии, рибосомы и полисомы, лизосомы, пероксисомы, микрофибриллы и микротрубочки, центриоли клеточного центра. В растительных клетках выделяют также хлоропласта, в которых происходит фотосинтез. Канальцевая и вакуолярная системы образованы сообщающимися или отдельными трубчатыми или уплощенными (цистерна) полостями, ограниченными мембранами и распространяющимися по всей цитоплазме клетки. Нередко цистерны имеют пузыревидные расширения. В названной системе выделяют шероховатую и гладкую цитоплазматическую сети (см. рис. 2.3). Особенность строения шероховатой сети состоит в прикреплении к ее мембранам полисом. В силу этого она выполняет функцию синтеза определенной категории белков, преимущественно удаляемых из клетки, например секретируемых клетками желез. В области шероховатой сети происходит образование белков и липидов цитоплазматических мембран, а также их сборка. Плотно упакованные в слоистую структуру цистерны шероховатой сети являются участками наиболее активного белкового синтеза и называются эргастоплазмой. Мембраны гладкой цитоплазматической сети лишены полисом. Функционально эта сеть связана с обменом углеводов, жиров и других веществ небелковой природы, например стероидных гормонов (в половых железах, корковом слое надпочечников). По канальцам и цистернам происходит перемещение веществ, в частности секретируемого железистой клеткой материала, от места синтеза в зону упаковки в гранулы. В участках печеночных клеток, богатых структурами гладкой сети, разрушаются и обезвреживаются вредные токсические вещества, некоторые лекарства (барбитураты). В пузырьках и канальцах гладкой сети поперечно-полосатой мускулатуры сохраняются (депонируются) ионы кальция, играющие важную роль в процессе сокращения. Рибосома — это округлая рибонуклеопротеиновая частица диаметром 20—30 нм. Она состоит из малой и большой субъединиц, объединение которых происходит в присутствии матричной (информационной) РНК (мРНК). Одна молекула мРНК обычно объединяет несколько рибосом наподобие нитки бус. Такую структуру называют полисомой. Полисомы свободно располагаются в основном веществе цитоплазмы или прикреплены к мембранам шероховатой цитоплазматической сети. В обоих случаях они служат местом активного синтеза белка. Сравнение соотношения количества свободных и прикрепленных к мембранам полисом в эмбриональных недифференцированных и опухолевых клетках, с одной стороны, и в специализированных клетках взрослого организма —с другой, привело к заключению, что на полисомах гиалоплазмы образуются белки для собственных нужд (для «домашнего» пользования) данной клетки, тогда как на полисомах гранулярной сети синтезируются белки, выводимые из клетки и используемые на нужды организма (например, пищеварительные ферменты, белки грудного молока). Пластинчатый комплекс Голъджи образован совокупностью диктиосом числом от нескольких десятков (обычно около 20) до нескольких сотен и даже тысяч на клетку. Диктиосома (рис. 2.6, А) представлена стопкой из 3—12 уплощенных дискообразных цистерн, от краев которых отшнуровываются пузырьки (везикулы). Ограниченные определенным участком (локальные) расширения цистерн дают более крупные пузырьки (вакуоли). В дифференцированных клетках позвоночных животных и человека диктиосомы обычно собраны в околоядерной зоне цитоплазмы. В пластинчатом комплексе образуются секреторные пузырьки или вакуоли, содержимое которых составляют белки и другие соединения, подлежащие выводу из клетки. При этом предшественник секрета (просекрет), поступающий.в диктиосому из зоны синтеза, подвергается в ней некоторым химическим преобразованиям. Он также обособляется (сегрегируется) в виде «порций», которые здесь же одеваются мембранной оболочкой. В пластинчатом комплексе образуются лизосомы. В диктиосомах синтезируются полисахариды, а также их комплексы с белками (гликопротеины) и жирами (гликолипиды), которые затем можно обнаружить в гликокаликсе клеточной оболочки. Митохондрии (рис. 2.6, Б) — это структуры округлой или палочко-видной, нередко ветвящейся формы толщиной 0,5 мкм и длиной обычно до 5—10 мкм. В большинстве животных клеток количество митохондрий колеблется от 150 до 1500, однако в женских половых клетках их число достигает нескольких сотен тысяч. В сперматозоидах нередко присутствует одна гигантская митохондрия, спирально закрученная вокруг осевой части жгутика. Одна разветвленная митохондрия обнаружена в клетке такого паразита человека, как трипаносома. Оболочка митохондрий состоит из двух мембран, различающихся по химическому составу, набору ферментов и функциям. Внутренняя мембрана образует впячивания листовидной (кристы) или трубчатой (тубулы) формы. Пространство, ограниченное внутренней мембраной, составляет матрикс органеллы. В нем с помощью электронного микроскопа обнаруживаются зерна диаметром 20—40 нм. Они накапливают ионы кальция и магния, а также полисахариды, например гликоген. В матриксе размещен собственный аппарат биосинтеза белка органеллы. Он представлен 2—б копиями кольцевой и лишенной гистонов (как у прокариот) молекулы ДНК, рибосомами, набором транспортных РНК (тРНК), ферментами редупликации ДНК, транскрипции и трансляции наследственной информации. По основным свойствам: размерам и структуре рибосом, организации собственного наследственного материала — этот аппарат сходен с таковым у прокариот и отличается от аппарата биосинтеза белка цитоплазмы эукариотической клетки (чем подтверждается симбиотическая гипотеза происхождения митохондрий; см. § 1.5). Гены собственной ДНК кодируют нуклеотидные последовательности митохондриальных рРНК и тРНК, а также последовательности аминонокислот некоторых белков органеллы, главным образом ее внутренней мембраны. Аминокислотные последовательности (первичная структура) большинства белков митохондрий закодированы в ДНК клеточного ядра и образуются вне органеллы в цитоплазме. Главная функция митохондрий состоит в ферментативном извлечении из определенных химических веществ энергии (путем их окисления) и накоплении энергии в биологически используемой форме (путем синтеза молекул аденозинтрифосфата —АТФ). В целом этот процесс называется окислительным (расформированием. В энергетической функции митохондрий активно участвуют компоненты матрикса и внутренняя мембрана. Именно с этой мембраной связаны цепь переноса электронов (окисление) и АТФ-синтетаза, катализирующая сопряженное с окислением фосфорилирование АДФ в АТФ. Среди побочных функций митохондрий можно назвать участие в синтезе стероидных гормонов и некоторых аминокислот (глутаминовая). Лизосомы (рис. 2.6, В) представляют собой пузырьки диаметром обычно 0,2—0,4 мкм, которые содержат набор ферментов кислых гидролаз, катализирующих при низких значениях рН гидролитическое (в водной среде) расщепление нуклеиновых кислот, белков, жиров, полисахаридов. Их оболочка образована одинарной мембраной, покрытой иногда снаружи волокнистым белковым слоем (на электронограммах «окаймленные» пузырьки). Функция лизосом — внутриклеточное переваривание оазличных химических соединений и структур. Первичными лизосомами (диаметр 100 нм) называют неактивные органеллы, вторичными — органеллы, в которых происходит процесс переваривания. Вторичные лизосомы образуются из первичных. Они подразделяются на гетеролизосомы (фаголизосомы) и аутолизосомы (цитолизосомы). В первых (рис. 2.6, Г) переваривается материал, поступающий в клетку извне путем пиноцитоза и фагоцитоза, во вторых разрушаются собственные структуры клетки, завершившие свою функцию. Вторичные лизосомы, в которых процесс переваривания завершен, называют остаточными тельцами (телолизосомы). В них отсутствуют гидролазы и содержится непереваренный материал. Микротельца составляют сборную группу органелл. Это ограниченные одной мембраной пузырьки диаметром 0,1—1,5 мкм с мелкозернистым матриксом и нередко кристаллоидными или аморфными белковыми включениями. К этой группе относят, в частности, пероксисомы. Они содержат ферменты оксидазы, катализирующие образование пероксида водорода, который, будучи токсичным, разрушается затем под действием фермента пероксидазы. Эти реакции включены в различные метаболические циклы, например в обмен мочевой кислоты в клетках печени и почек. В печеночной клетке число пероксисом достигает 70—100. К органеллам общего значения относят также некоторые постоянные структуры цитоплазмы, лишенные мембран. Микротрубочки (рис. 2.6, Д) — трубчатые образования различной длины с внешним диаметром 24 нм, шириной просвета 15 нм и толщиной стенки около 5 нм. Встречаются в свободном состоянии в цитоплазме клеток или как структурные элементы жгутиков, ресничек, митотического веретена, центриолей. Свободные микротрубочки и микротрубочки ресничек, жгутиков и центриолей имеют разную устойчивость к разрушающим воздействиям, например химическим (колхицин). Микротрубочки строятся из стереотипных субьединиц белковой природы путем их полимеризации. В живой клетке процессы полимеризации протекают одновременно с процессами деполимеризации. Соотношением этих процессов определяется количество микротрубочек. В свободном состоянии микротрубочки выполняют опорную функцию, определяя форму клеток, а также являются факторами направленного перемещения внутриклеточных компонентов. Микрофиламентами (рис. 2.6, Е) называют длинные, тонкие образования, иногда образующие пучки и обнаруживаемые по всей цитоплазме. Существует несколько разных типов микрофиламентов. Актиновые микрофиламенты благодаря присутствию в них сократимых белков (актин) рассматривают в качестве структур, обеспечивающих клеточные формы движения, например амебоидные. Им приписывают также каркасную роль и участие в организации внутриклеточных перемещений органелл и участков гиалоплазмы. По периферии клеток под плазмалеммой, а также в околоядерной зоне обнаруживаются пучки микрофиламентов толщиной 10 нм — промежуточные филстенты. В эпителиальных, нервных, глиальных, мышечных клетках, фибробластах они построены из разных белков. Промежуточные филаменты выполняют, по-видимому, механическую, каркасную функцию. Актиновые микрофибриллы и промежуточные филаменты, как и микротрубочки, построены из субъединиц. В силу этого их количество зависит от соотношения процессов полимеризации и деполимеризации. Для животных клеток, части клеток растений, грибов и водорослей характерен клеточный центр, в состав которого входят центриоли. Центриолъ (под электронным микроскопом) имеет вид «полого» цилиндра диаметром около 150 нм и длиной 300—500 нм. Ее стенка образована 27 микротрубочками, сгруппированными в 9 триплетов. В функцию центриолей входит образование нитей митотического веретена, которые также образованы микротрубочками. Центриоли поляризуют процесс деления клетки, обеспечивая расхождение сестринских хроматид (хромосом) в анафазе митоза. Эукариотическая клетка имеет клеточный скелет (цитоскелет) из внутриклеточных волокон (Кольцов) – начало ХХ века, в конце 1970 вновь открыт. Эта структура позволяет клетке иметь свою форму, иногда изменяя ее. Цитоплазма находится в движении. Цитоскелет участвует с процессе переноса органоидов, участвует в регенерации клеток. Митохондрии – сложные образования с двойной мембраной(0,2-0,7мкм) и разной формой. Внутренняя мембрана имеет кристы. Наружная мембрана проницаема практически для всех химических веществ, внутренняя – только активный транспорт. Между мембранами – матрикс. Митохондрии располагаются там, где необходима энергия. Митохондрии имеют систему рибосом, молекулу ДНК. Возможно возникновение мутаций (более66 заболеваний). Как правило, они связаны с недостаточной энергией АТФ, часто связаны с сердечно-сосудистой недостаточностью, патологиями. Количество митохондрий разное (в клетке трипаносомы- 1 митохондрия). Количество зависит от возраста, функции, активности ткани (печень – более1000). Лизосомы – тельца, окруженные элементарной мембраной. Содержат 60 ферментов( 40 лизосомальных, гидролитических). Внутри лизосомы – нейтральная среда. Активизируются низкими значениями рН, выходя в цитоплазму (самопереваривание). Мембраны лизосом защищают цитоплазму и клетку от разрушения. Образуются в комплексе Гольджи (внутриклеточный желудок, могут перерабатывать отработавшие свое структуры клетки). Есть 4 вида. 1-первичные, 2-4 – вторичные. С помощью эндоцитоза в клетку попадает вещество. Первичная лизосома (запасающая гранула) с набором ферментов, поглощает вещество и образуется пищеварительная вакуоль (при полном переваривании расщепление идет до низкомолекулярных соединений). Непереваренные остатки остаются в остаточных тельцах, которые могут накапливаться (лизосомные болезни накопления). Остаточные тельца, накапливающиеся в эмбриональном периоде, приводят к гаргалеизму, уродствам, мукополисахаридозам. Аутофагирующие лизосомы уничтожают собственные структуры клетки( ненужные структуры). Могут содержать митохондрии, части комплекса Гольджи. Часто образуются при голодании. Могут возникать при воздействии других клеток (эритроциты). 12.Поток информации, энергии и вещества в клетке. Жизнедеятельность клетки как единицы биологической активности обеспечивается совокупностью взаимосвязанных, приуроченных к определенным внутриклеточным структурам, упорядоченных во времени и пространстве обменных (метаболических) процессов. Эти процессы образуют три потока: информации, энергии и веществ. Благодаря наличию потока информации клетка на основе многовекового эволюционного опыта предков приобретает структуру, отвечающую критериям живого, поддерживает ее во времени, а также передает в ряду поколений. В потоке информации участвуют ядро (конкретно ДНК хромосом), макромолекулы, переносящие информацию в цитоплазму (мРНК), цитоплазматический аппарат трансляции (рибосомы и полисомы, тРНК, ферменты активации аминокислот). На завершающем этапе этого потока полипептиды, синтезированные на полисомах, приобретают третичную и четвертичную структуры и используются в качестве катализаторов или структурных белков (рис. 2.7). Кроме основного по объему заключенной информации ядерного генома в эукариотических клетках функционируют также геномы митохондрий, а в зеленых растениях — и хлоропластов. 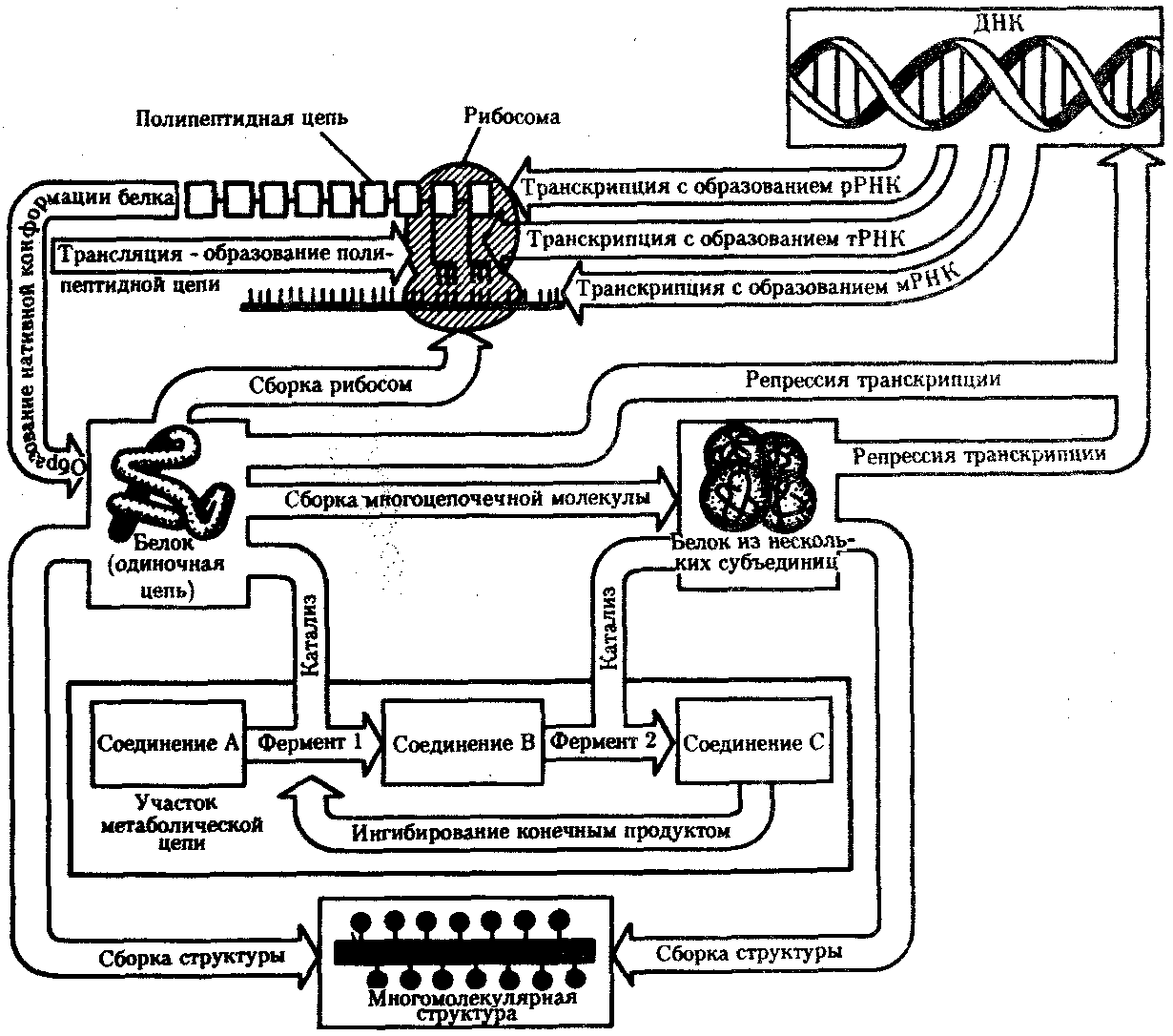 Рис. 2.7. Поток биологической информации в клетке 2.3.4. Внутриклеточный поток энергииПоток энергии у представителей разных групп организмов обеспечивается механизмами энергоснабжения —брожением, фото- или хемосинтезом, дыханием. Центральная роль в биоэнергетике клеток животных принадлежит дыхательному обмену. Он включает реакции расщепления низкокалорийного органического «топлива» в виде глюкозы, жирных кислот, аминокислот, а также использование выделяемой энергии для образования высококалорийного клеточного «топлива» в виде аденозинтрифосфата (АТФ). Энергия АТФ, непосредственно или будучи перенесена на другие макроэргические соединения (например, креатинфосфат), в разнообразных процессах преобразуется в тот или иной вид работы — химическую (синтезы), осмотическую (поддержание перепадов концентрации веществ), электрическую, механическую, ре-гуляторную. Макроэргическим называют соединение, в химических связях которого запасена энергия в форме, доступной для использования в биологических процессах. Универсальным соединением такого рода служит АТФ. Основное количество энергии заключено в связи, присоединяющей третий остаток фосфорной кислоты. 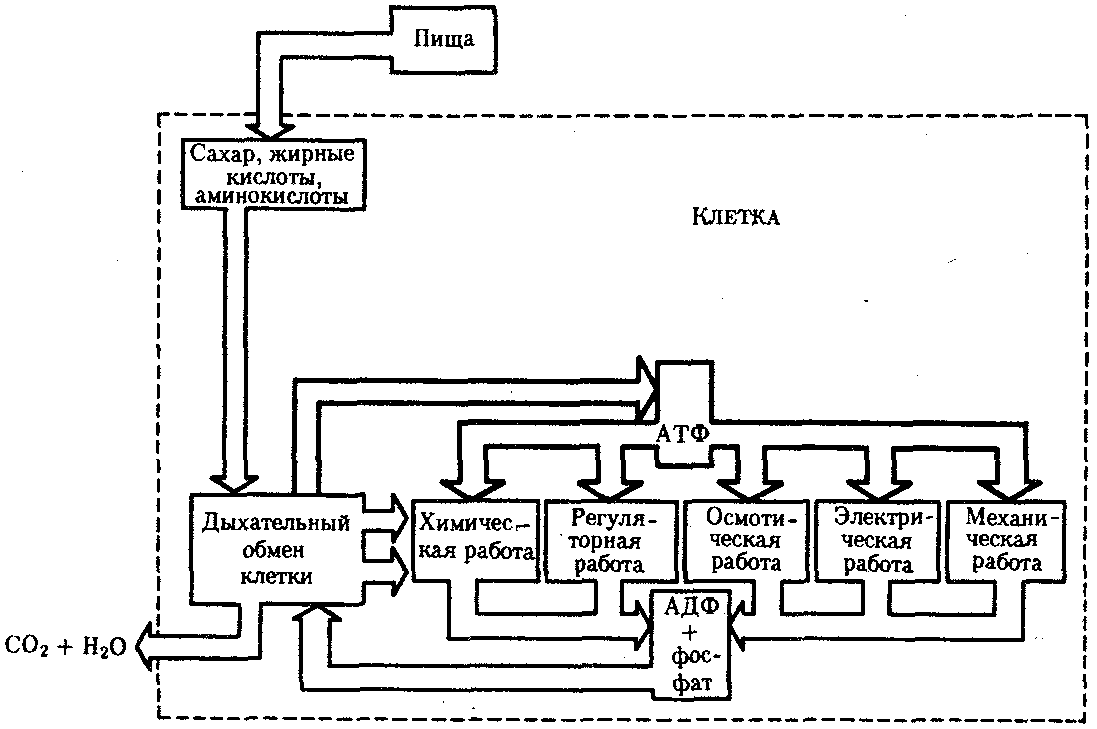 Рис. 2.8. Поток энергии в клетке Среди органелл животной клетки особое место в дыхательном обмене принадлежит митохондриям, выполняющим функцию окислительного фосфорилирования, а также матриксу цитоплазмы, в котором протекает процесс бескислородного расщепления глюкозы — анаэробный гликолиз (рис. 2.8). Из двух механизмов, обеспечивающих жизнедеятельность клетки энергией, анаэробный гликолиз менее эффективен. В связи с неполным (в отсутствие кислорода) окислением, прежде всего глюкозы, в процессе гликолиза для нужд клетки извлекается не более 10% энергии. Недоокисленные продукты гликолиза (пируват) поступают в митохондрий, где в условиях полного окисления, сопряженного с фосфорилированием АДФ до АТФ, отдают для нужд клетки оставшуюся в их химических связях энергию. Из преобразователей энергии химических связей АТФ в работу наиболее изучена механохимическая система поперечно-полосатой мышцы. Она состоит из сократительных белков (актомиозиновый комплекс) и фермента аденозинтрифосфатазы, расщепляющего АТФ с высвобождением энергии. Особенность потока энергии растительной клетки состоит в наличии фотосинтеза — механизма преобразования энергии солнечного света в энергию химических связей органических веществ. Механизмы энергообеспечения клетки отличаются эффективностью. Коэффициенты полезного действия хлоропласта и митохондрий, достигая соответственно 25 и 45—60%, существенно превосходят аналогичный показатель паровой машины (8%) или двигателя внутреннего сгорания (17%). 2.3.5. Внутриклеточный поток веществРеакции дыхательного обмена не только поставляют энергию, но и снабжают клетку строительными блоками для синтеза разнообразных молекул. Ими являются многие продукты расщепления пищевых веществ. Особая роль в этом принадлежит одному из этапов дыхательного обмена — циклу Кребса,осуществля-емому в митохондриях. Через этот цикл проходит путь углеродных атомов (углеродных скелетов) большинства соединений, служащих промежуточными продуктами синтеза химических компонентов клетки. В цикле Кребса происходит выбор пути превращения того или иного соединения, а также переключение обмена клетки с одного пути на другой, например с углеводного на жировой. Таким образом, дыхательный обмен одновременно составляет ведущее звено потока веществ, объединяющего метаболические пути расщепления и образования углеводов, белков, жиров, нуклеиновых кислот (рис. 2.9). 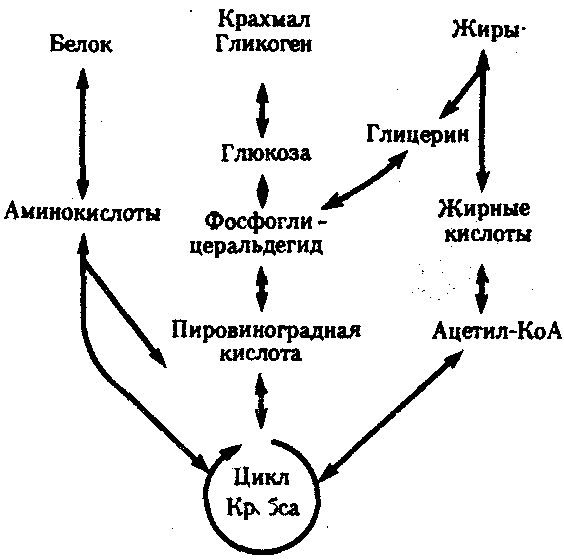 Рис. 2.9. Взаимосвязь внутриклеточного обмена белков, жиров и углеводов через цикл Кребса 13.Жизненный и митотический (пролиферативный) цикл клетки. Фазы митотического цикла, их характеристика и значение. 14.Наследственность и изменчивость – свойства, определяющие непрерывность существования и развития жизни. Уровни структурно-функциональной организации наследственного материала: генный, хромосомный и геномный. 15.Структура ДНК, её свойства и функции. Репликация ДНК. 16.Классификация нуклеотидных последовательностей в геноме эукариот (уникальные и повторяющиеся последовательности). 17.Мутации, их классификации и механизмы возникновения. Медицинское и эволюционное значение. 18.Репарация как механизм поддержания генетического гомеостаза. Виды репарации. Мутации, связанные с нарушением репарации и их роль в патологии. 19.Ген, его свойства. Генетический код, его свойства. Структура и виды РНК. Процессинг, сплайсинг. Роль РНК в процессе реализации наследственной информации. 20.Рибосомный цикл синтеза белка (инициация, элонгация, терминация). Посттрансляционные преобразования белков. 21.Взаимосвязь между геном и признаком. Гипотеза «один ген - один фермент», ее современная трактовка: «один ген – одна полипептидная цепь» 22.Ген как единица изменчивости. Генные мутации и их классификация. Причины и механизмы возникновения генных мутаций. Последствия генных мутаций. 23.Хромосома, её химический состав. Хроматин: структурная организация (уровни компактизации), классификация (гетерохроматин, эухроматин). Метафазная хромосома. Морфология хромосом. 24.Хромосомные мутации, их классификация. Причины и механизмы возникновения хромосомных мутаций. Роль хромосомных мутаций в развитии патологических состояний и эволюционном процессе. 25.Геном, кариотип как видовые характеристики. Характеристика кариотипа человека в норме. 26.Геном как эволюционно сложившаяся система генов. Функциональная классификация генов (структурные, регуляторные). Регуляция экспрессии генов у прокариот и эукариот. 27.Геномные мутации, причины и механизмы их возникновения. Классификация и значение геномных мутаций. 28.Эволюция генома. Роль ампфликации генов, хромосомных перестроек, полиплоидизации, подвижных генетических элементов, горизонтального переноса информации в эволюции генома. Секвенирование генома. 29.Размножение. Способы и формы размножения организмов. Половое размножение, его эволюционное значение. 30.Гаметогенез. Мейоз. Цитологическая и цитогенетическая характеристика. Особенности ово- и сперматогенеза у человека. Мейоз состоит из 2 последовательных делений с короткой интерфазой между ними.
Второе деление мейоза следует непосредственно за первым, без выраженной интерфазы: S-период отсутствует, поскольку перед вторым делением не происходит репликации ДНК.
В результате из одной диплоидной клетки образуется четыре гаплоидных клетки. В тех случаях, когда мейоз сопряжён с гаметогенезом (например, у многоклеточных животных), при развитии яйцеклеток первое и второе деления мейоза резко неравномерны. В результате формируется одна гаплоидная яйцеклетка и два так называемых редукционных тельца (абортивные дериваты первого и второго делений). 31.Морфология половых клеток. 32.Оплодотворение, его фазы, биологическая сущность. Партеногенез. Типы определения пола. 33.Предмет, задачи, методы генетики. История развития генетики. Роль отечественных ученых (Н. И. Вавилов, Н. К. Кольцов, А. С. Серебровский, С. С. Четвериков) в развитии генетики. 34.Понятия: генотип, фенотип, признак. аллельные и неаллельные гены, гомозиготные и гетерозиготные организмы, понятие гемизиготности. Признак - единица морфологической, физиологической, биохимической, иммунологической, клинической и любой другой дискретности организмов (клеток), т.е. отдельное качество или свойство, по которому они отличаются друг от друга. Генотип — это генетическая конституция организма, представляющая собой совокупность всех наследственных задатков его клеток, заключенных в их хромосомном наборе — кариотипе. Генотип (от ген и тип), совокупность всех генов, локализованных в хромосомах данного организма. Фенотип (Phenotype) — присущая индивидууму совокупность всех признаков и свойств, которые сформировались в процессе его индивидуального развития. Фенотип - совокупность всех признаков организма, сформировавшаяся во взаимодействии генотипа с окружающей средой. Гомозиготность, состояние наследственного аппарата организма, при котором гомологичные хромосомы имеют одну и ту же форму данного гена. Гетерозиготность, присущее всякому гибридному организму состояние, при котором его гомологичные хромосомы несут разные формы (аллели) того или иного гена. Гемизиготность (от греч hemi- — полу- и zygotós — соединённый вместе), состояние, связанное с тем, что у организма один или несколько генов не парные, т. е. не имеют аллельных партнёров. (В сцепленном с полом наследовании, Хr или ХR — r – дальтонзим) 35.Закономерности наследования при моногибридном скрещивании. Моногибридное скрещивание — скрещивание форм, отличающихся друг от друга по одной паре альтернативных признаков. 1 закон Менделя: при скрещивании двух гомозиготных организмов, различающихся друг от друга по одной паре альтернативных признаков в первом поколении наблюдается единообразие по генотипу и по фенотипу. (фиброматоз десен — А, здоровые десны — а, ребенок в любом случае болен) 2 закон менделя: при скрещивании 2х гетерозиготных организмов, различающихся по одной паре альтернативных признаков (гибриды F1) в их потомстве (гибриды F2) наблюдает расщепление по фенотипу 3:1, по генотипу 1:2:1 Полное доминирование — явление, при котором один из аллельных генов имеет преобладающее значение и проявляется как в гетерозиготном, так и в гомозиготном состоянии. 36.Дигибридное и полигибридное скрещивание. Закон независимого комбинирования генов и его цитологические основы. Общая формула расщепления при независимом наследовании. Дигибридное скрещивание - скрещивание форм, различающихся по двум парам изучаемых признаков При скрещивании гомозиготных особей, которые отличаются двумя и большим количеством пар альтернативных признаков, во втором гибридном поколении (при инбридинге гибридов 1 поколения) фиксируют независимое наследование по каждой паре признаков и появляются особи, с новыми сочетаниями признаков, не свойственных родительским и прародительским формам (закон независимого распределения, или III закон Менделя) (Карие глаза — B, голубые — b, правша — A, левша — a) Очевидно, этому закону должны подчиняться в первую очередь неаллельные гены, располагающиеся в разных (негомологичных) хромосомах. В таком случае независимый характер наследования признаков объясняется закономерностями поведения негомологичных хромосом в мейозе. Названные хромосомы образуют со своими гомологами разные пары, или биваленты, которые в метафазе I мейоза случайно выстраиваются в плоскости экватора веретена деления. Затем в анафазе I мейоза гомологи каждой пары расходятся к разным полюсам веретена независимо от других пар. В результате у каждого из полюсов возникают случайные сочетания отцовских и материнских хромосом в гаплоидном наборе (см. рис. 3.75). Следовательно, различные гаметы содержат разные комбинации отцовских и материнских аллелей неал-лельных генов. Разнообразие типов гамет, образуемых организмом, определяется степенью его гетерозиготности и выражается формулой 2n, где n — число локусов в гетерозиготном состоянии. В связи с этим дигетерозиготные гибриды F1 образуют четыре типа гамет с одинаковой вероятностью. Реализация всех возможных встреч этих гамет при оплодотворении приводит к появлению в F2 четырех фенотипических групп потомков в соотношении 9:3:3:1. Анализ потомков F2 по каждой паре альтернативных признаков в отдельности выявляет расщепление в соотношении 3:1. 37.Множественные аллели. Наследование групп крови человека системы АВО. Множественный аллелизм — различные состояния (три и более) одного и того же локуса хромосом, возникшие в результате мутаций. Присутствие в генофонде вида одновременно различных аллелей гена называют множественным аллелизмом. Примером этому служат разные варианты окраски глаз у плодовой мухи: белая, вишневая, красная, абрикосовая, эозиновая,— обусловленные различными аллелями соответствующего гена. У человека, как и у других представителей органического мира, множественный аллелизм свойствен многим генам. Так, три аллеля гена I определяют групповую принадлежность крови по системе АВ0 (IA, IB, I0). Два аллеля имеет ген, обусловливающий резус-принадлежность. Более ста аллелей насчитывают гены α- и β-полипептидов гемоглобина. Причиной множественного аллелизма являются случайные изменения структуры гена (мутации), сохраняемые в процессе естественного отбора в генофонде популяции. Многообразие аллелей, рекомбинирующихся при половом размножении, определяет степень генотипического разнообразия среди представителей данного вида, что имеет большое эволюционное значение, повышая жизнеспособность популяций в меняющихся условиях их существования. Кроме эволюционного и экологического значения аллельное состояние генов оказывает большое влияние на функционирование генетического материала. В диплоидных соматических клетках эукариотических организмов большинство генов представлено двумя аллелями, которые совместно влияют на формирование признаков. 38.Взаимодействие неаллельных генов: комплементарность, эпистаз, полимерия, модифицирующее действие. Комплементарность — такой тип взаимодействия, когда 2 неаллельных гена, попадая в генотип в доминирующем состоянии, совместно определяют появление нового признака, который каждый из них по отдельности не детерминирует.(R- розовидный гребень, P – гороховидный, rp – листовидный, RP – ореховидный) Если присутствует один из пары – проявляется он. Примером служат группы крови у человека. Комплементарность может быть доминантная и рецессивная. Для того чтобы человек имел нормальный слух, необходимо чтобы работали, согласовано многие гены, и доминантные и рецессивные. Если, хотя бы по одному гену он будет гомозиготен по рецессиву – слух будет ослаблен. Эпистаз — маскирование генов одной аллельной пары генами другой. Эпистаз (от греч. epi - над + stasis -- препятствие) -- взаимодействие неаллельных генов, при котором наблюдается подавление проявления одного гена действием другого, неаллелъного гена. Ген, подавляющий фенотипические проявления другого, называется эпистатичным; ген, чья активность изменена или подавлена, называется гипостатичным. Это обусловлено тем, что ферменты катализируют разные процессы клетки, Когда на одном метаболическом пути действуют несколько генов. Действие их должно быть согласовано во времени. Механизм: если В выключится, он замаскирует действие С В  одних случаях развитие признака при наличии двух неаллельных генов в доминантном состоянии рассматривают как комплементарное взаимодействие, в других — неразвитие признака, определяемого одним из генов при отсутствии другого гена в доминантном состоянии, расценивают как рецессивный эпистаз; если же признак развивается при отсутствии доминантного аллеля неаллельного гена, а в его присутствии не развивается, говорят о доминантном эпистазе. Полимерия — явление, когда различные неаллельные гены могут оказывать однозначное действие на один и тот же признак, усиливая его проявление. Наследование признаков при полимерном взаимодействии генов. В том случае, когда сложный признак определяется несколькими парами генов в генотипе и их взаимодействие сводится к накоплению эффекта действия определенных аллелей этих генов, в потомстве гетерозигот наблюдается разная степень выраженности признака, зависящая от суммарной дозы соответствующих аллелей. Например, степень пигментации кожи у человека, определяемая четырьмя парами генов, колеблется от максимально выраженной у гомозигот по доминантным аллелям во всех четырех парах (Р1Р1Р2Р2Р3Р3Р4Р4) до минимальной у гомозигот по рецессивным аллелям (р1р1р2р2р3р3р4р4) (см. рис. 3.80). При браке двух мулатов, гетерозиготных по всем четырем парам, которые образуют по 24 = 16 типов гамет, получается потомство, 1/256 которого имеет максимальную пигментацию кожи, 1/256 — минимальную, а остальные характеризуются промежуточными показателями экспрессивности этого признака. В разобранном примере доминантные аллели полигенов определяют синтез пигмента, а рецессивные — практически не обеспечивают этого признака. В клетках кожи организмов, гомозиготных по рецессивным аллелям всех генов, содержится минимальное количество гранул пигмента. В некоторых случаях доминантные и рецессивные аллели полигенов могут обеспечивать развитие разных вариантов признаков. Например, у растения пастушьей сумки два гена одинаково влияют на определение формы стручочка. Их доминантные аллели образуют одну, а рецессивные — другую форму стручочков. При скрещивании двух дигетерозигот по этим генам (рис. 6.16) в потомстве наблюдается расщепление 15:1, где 15/16 потомков имеют от 1 до 4 доминантных аллелей, а 1/16, не имеет доминантных аллелей в генотипе. Если гены располагаются, каждый в своем отдельном локусе, но их взаимодействие проявляется в одном и том же направлении – это полигены. Один ген проявляет признак незначительно. Полигены дополняют друг друга и оказывают мощное действие – возникает полигенная система – т.е. система является результатом действия одинаково направленных генов. Гены подвергаются значительному влиянию главных генов, которых более 50. полигенных систем известно множество. При сахарном диабете наблюдается умственная отсталость. Рост, уровень интеллекта - определяются полигенными системами Модифицирующее действие. Гены модификаторы сами по себе не определяют какой- то признак, но могут усиливать или ослаблять действие основных генов, вызывая таким образом изменение фенотипа. В качестве примера обычно приводится наследование пегости у собак и лошадей. Числового расщепления никогда не даётся, так как характер наследования больше напоминает полигенное наследование количественных признаков. 1919 год Бриджес ввел термин ген-модификатор. Теоретически любой ген может взаимодействовать с другими генами, а значит, и проявлять модифицирующее действие, но некоторые гены – модификаторы в большей степени. Они часто не имеют собственного признака, но способны усиливать или ослаблять проявление признака, контролируемого другим геном. В формировании признака кроме основных генов проявляют свое действие и модифицирующие гены. Брахидактилия – может быть резкая или незначительная. Помимо основного гена, есть еще модификатор, который усиливает эффект. Окраска млекопитающих – белая, черная + модификаторы. 39.Хромосомная теория наследственности. Сцепление генов. Группы сцепления. Кроссинговер как механизм, определяющий нарушения сцепления генов. |