ответы по гистологии(Галка Яна). Ответ 14! Немембранные органеллы. Рибосомы. Строение, химический состав, функции. Понятие о полисомах. Роль свободных и связанных с мембранами эндоплазматической сети рибосом в биосинтезе клеточных белков. Рибосомы
 Скачать 275.25 Kb. Скачать 275.25 Kb.
|

Гуморальная регуляцияПервая древнейшая форма взаимодействия между клетками многоклеточных организмов — это химическое взаимодействие посредством продуктов обмена веществ, поступающих в жидкости организма. Такими продуктами обмена веществ, или метаболитами, служат продукты распада белков, углекислота и др. Это гуморальная передача влияний, гуморальный механизм корреляции или связи между органами. Гуморальная связь характеризуется следующими особенностями. Во-первых, отсутствием точного адреса, по которому направляется химическое вещество, поступающее в кровь или другие жидкости тела. Химическое вещество может, следовательно, действовать на все органы и ткани. Его действие не локализовано, не ограничено определенным местом. Во-вторых, химическое вещество распространяется относительно медленно. И, наконец, в-третьих, оно действует в ничтожных количествах и обычно быстро разрушается или выводится из организма. Гуморальные связи являются общими и для мира животных и мира растений. ПлазмодесмыМикроскопические цитоплазматические мостики, соединяющие соседние клетки растений. Простое межклеточное соединениеПри простом межклеточном соединении оболочки клеток сближены на расстояние 15 – 20 нм. Это соединение занимает наиболее обширные участки соприкасающихся клеток. Посредством простых соединений осуществляется слабая механическая связь, не препятствующая транспорту веществ в межклеточных пространствах. Разновидностью простого соединения является контакт типа «замок», когда билипидные мембраны соседних клеток вместе с участком цитоплазмы вдавливаются друг в друга, чем достигается большая поверхность соприкосновения и более прочная механическая связь. Плотное соединение (запирающая зона)В плотном соединении клеточные мембраны максимально сближены, здесь фактически происходит их слияние. Роль плотного соединения заключается в механическом сцеплении клеток и препятствии транспорту веществ по межклеточным пространствам. Эта область непроницаема для макромолекул и ионов, она ограждает межклеточные щели от внешней среды. Плотные соединения обычно образуются между эпителиальными клетками в тех органах (желудке, кишечнике и пр.), где эпителий ограничивает содержимое этих органов (желудочный сок, кишечный сок). В этих участках плотные контакты охватывают по периметру каждую клетку, межмембранные пространства отсутствуют, а соседние клеточные оболочки слиты в одну. Если же плотное сцепление происходит на ограниченном участке, то образуется пятно слипания (десмосома).Частными случаями плотного соединения являются зоны замыкания и слипания. Зона замыканияВ зоне замыкания две соседние мембраны сливаются своими наружными слоями, эта зона непроницаема для макромолекул и ионов. Зона слипания (промежуточный контакт)В зоне слипания мембраны разделены щелью в 10-20 нм, заполненной плотным веществом (белковой природы). Десмосома (пятно сцепления, липкое соединение)Десмосома представляет собой небольшую площадку, иногда слоистого вида, диаметром до 0,5 мкм. Их функциональная роль заключается главным образом в механической связи между клетками. Существуют 3 типа десмосом – точечные, опоясывающие и полудесмосомы. Десмосомой называется образованное клетками соединение, прочно склеивающее клетки. Если они образуются между клетками и внеклеточным матриксом, то они называются полудесмосомами. Количество десмосом на одной клетке может достигать 2000. Такие контакты встречаются между клетками, которые могут подвергаться трению и другим механическим воздействиям (эпителиальные клетки, клетки сердечной мышцы). Со стороны цитоплазмы к десмосомам прикрепляются промежуточные филаменты, которые формируют остов цитоплазмы, обладающий большой прочностью на разрыв. Таким образом, через десмосомы промежуточные филаменты соседних клеток объединяются в непрерывную сеть по всей ткани. Тип промежуточных филаментов зависит от типа клеток: в большинстве эпителиальных клеток они кератиновые, а в клетках сердечной мышцы – десминовые. Нексус (щелевой контакт)Нексус представляет собой ограниченный участок контакта двух клеточных мембран диаметром 0,5 – 3 мкм с расстоянием между мембранами 2-3 нм. Обе эти мембраны пронизаны белковыми молекулами коннексонами, содержащими гидрофильные каналы. Через эти каналы осуществляется обмен ионами и микромолекулами соседних клеток. Поэтому нексусы называют также проводящими соединениями. Их функциональная роль заключается в переносе ионов и мелких молекул от клетки к клетке, минуя межклеточное пространство. Этот тип соединения встречается во всех группах тканей. Синапс (синаптическое соединение)Синапсы являются особыми формами межклеточных соединений. Они характерны для нервной ткани и встречаются между нейронами (межнейронные синапсы) или между нейроном и клеткой-мишенью (нервно-мышечные синапсы и пр.). Синапсы – участки контакта двух клеток, специализированных для односторонней передачи возбуждения или торможения от одной клетки к другой. Их функция – именно передача нервного импульса с нейрона на другую нервную клетку или клетку-мишень.4. Реакция клеток на внешние воздействия Описанная морфология клеток не является стабильной (постоянной). При воздействии на организм различных неблагоприятных факторов в строении различных структур проявляются различные изменения. В зависимости от факторов воздействия изменения клеточных структур проявляются неодинаково в клетках разных органов и тканей. При этом изменения клеточных структур могут быть адаптивными (приспособительными) и обратимыми, или же дезадаптивными, необратимыми (патологическими). Однако определить четкую грань между адаптивными и дезадаптивными изменениями не всегда возможно, так как приспособительные изменения могут перейти в патологические. Поскольку объектом изучения гистологии являются клетки, ткани и органы здорового организма человека, то здесь будут рассмотрены прежде всего адаптивные изменения клеточных структур. Изменения отмечаются как в строении ядра, так и цитоплазмы. Изменения в ядре – набухание ядра и сдвиг его на периферию клетки, расширение перинуклеарного пространства, образование инвагинаций кариолеммы (впячивание внутрь ядра его оболочки), конденсация хроматина. К патологическим изменениям ядра относят: · пикноз – сморщивание ядра и коагуляция (уплотнение) хроматина; · кариорексис – распад ядра на фрагменты; · кариолизис – растворение ядра. Изменения в цитоплазме – уплотнение, а затем набухание митохондрий, дегрануляция зернистой эндоплазматической сети (слущивание рибосом), а затем и фрагментация канальцев на отдельные вакуоли, расширение цистерн, а затем распад на вакуоли пластинчатого комплекса Гольджи, набухание лизосом и активация их гидролаз, увеличение числа аутофагосом, в процессе митоза – распад веретена деления и развитие патологических митозов. Изменения цитоплазмы могут быть обусловлены структурными изменениями плазмолеммы, что приводит к усилению ее проницаемости и гидратации гиалоплазмы, нарушением обмена веществ, что сопровождается снижением содержания АТФ, снижением расщепления или увеличением синтеза включений (гликогена, липидов) и их избыточном накоплении. После устранения неблагоприятных воздействий на организм реактивные (адаптивные) изменения структур исчезают и морфология клетки восстанавливается. При развитии патологических (дезадаптивных) изменений даже после устранения неблагоприятных воздействий структурные изменения нарастают и клетка погибает. Ответ 23! Воспроизведение клеток. Клеточный цикл. Определение, этапы клеточного цикла для клеток, сохранивших и утративших способность к делению. Морфофункциональная характеристика процессов роста и дифференцировки, периода активного функционирования, старения и гибели клеток. Апоптоз (программированная гибель клеток). Определение понятия и его биологическое значение. ВОСПРОИЗВЕДЕНИЕ КЛЕТОК Все живые организмы состоят из клеток. Развитие, рост, становление типичной структуры организма осуществляется благодаря одной или группы исходных клеток. В процессе жизнедеятельности часть клеток изнашивается, стареет и погибает. Для поддержания структуры и нормального функционирования организм должен производить новые клетки на смену старым. Единственным способом образования клеток является деление предыдущих. Деление клеток – жизненно важный процесс для всех организмов. 2. Клеточный, или жизненный, цикл клетки – это время существования клетки от деления до следующего деления, или от деления до смерти. Для разных типов клеток клеточный цикл различен. В организме млекопитающих и человека различают следующие три группы клеток, локализующиеся в разных тканях и органах: · часто делящиеся клетки (малодифференцированные клетки эпителия кишечника, базальные клетки эпидермиса и другие); · редко делящиеся клетки (клетки печени – гепатоциты); · неделящиеся клетки (нервные клетки центральной нервной системы, меланоциты и другие). Жизненный цикл у этих клеточных типов различен. Жизненный цикл у часто делящихся клеток – это время их существования от начала деления до следующего деления. Жизненный цикл таких клеток нередко называют митотическим циклом. Такой клеточный цикл подразделяется на два основных периода: · митоз или период деления; · интерфаза – промежуток жизни клетки между двумя делениями. Различают два основных способа размножения клеток: · митоз (кариокенез) – непрямое деление клеток, которое присуще в основном соматическим клеткам; · мейоз или редукционное деление – характерно только для половых клеток. В литературе нередко описывают третий способ деления клеток – амитоз или прямое деление клеток, которое осуществляется посредством перетяжки ядра и цитоплазмы, с образованием двух дочерних клеток или одной двуядерной. Однако в настоящее время принято считать, что прямой способ деления характерен только для старых и дегенерирующих клеток и является отражением патологии клетки. Возможен четвертый тип репродукции клетки – эндорепродукция, характеризуется увеличением объема клетки, увеличением количеством ДНК в хромосомах, увеличивается количество функциональных органелл. Клетка является гипертрофированной, но к увеличению числа клеток эндорепродукция не приводит, а лишь повышается функциональная активность клеток. Она наблюдается в клетках печенигепатоцитах, в эпителии мочевого пузыря. Отмеченные выше два основных периода в жизненном цикле часто делящихся клеток (митоз и интерфаза) в свою очередь подразделяются на фазы или периоды. Митоз подразделяется на 4 фазы: · профаза; · метофаза; · анафаза; · телофаза. В каждой фазе происходят определенные структурные преобразования. Профаза характеризуется морфологическими изменениями ядра и цитоплазмы. В ядре происходит: конденсация хроматина и образование хромосом, состоящих из двух хроматид, исчезновение ядрышка, распад кариолеммы на отдельные пузырьки. В цитоплазме отмечается редупликация (удвоение) центриолей и расхождение их к противоположным полюсам клетки, формирование из микротрубочек веретена деления, репродукция зернистой эндоплазматической сети, а также уменьшение числа свободных и прикрепленных рибосом. В метафазе происходит образование метафазной пластинки, или материнской звезды, неполное обособление сестринских хроматид друг от друга. Анафаза характеризуется полным обособлением (расхождением) хроматид и образованием двух равноценных диплоидных наборов хромосом, расхождением хромосомных наборов к полюсам митотического веретена и расхождением самих полюсов. Телофаза характеризуется деконденсацией хромосом каждого хромосомного набора, формированием из пузырьков ядерной оболочки, цитотомиейперетяжкой двуядерной клетки на две дочерние самостоятельные клетки, появлением ядрышка в ядрах дочерних клеток. Интерфаза подразделяется на 3 периода: · J1, или пресинтетический; · S, или синтетический; · J2, или постсинтетический. Каждый период характеризуется прежде всего некоторыми функциональными особенностями. В J1 (пресинтетическом) периоде происходит: · усиленное формирование синтетического аппарата клетки – увеличение числа рибосом, а также количества различных видов РНК (информационной, рибосомальной, транспортных); · усиление синтеза белков, необходимых для роста клетки; · подготовка клетки к синтетическому периоду – синтез ферментов, необходимых для образования новых молекул ДНК. Для S-периода характерно удвоение (редупликация) ДНК, что приводит к удвоению плоидности диплоидных ядер и является обязательным условием для последующего митотического деления клетки. J2-период (постсинтетический, или премитотический) характеризуется усиленным синтезом информационной РНК, а также усиленным синтезом всех клеточных белков, но особенно белков-тубулинов, необходимых для последующего (в профазе митоза) формирования митотического веретена деления. Описанные закономерности жизненного цикла характерны прежде всего для часто делящихся клеток. Однако клетки некоторых тканей (например, клетки печеночной ткани – гепатоциты), по выходе из митоза, вступают в так называемый J0-период, во время которого они выполняют свои многочисленные функции в течении многих лет, не вступая в S-период. Однако при определенных обстоятельствах (при поражении или удалении части печени) они вступают в нормальный клеточный цикл, то есть в S-период, синтезируют ДНК, а затем митотически делятся. Такие клетки относятся к редко делящимся клеткам, и их жизненный цикл подразделяется на митоз, J0-период, S-период, J2-период. Большинство клеток нервной ткани, особенно нейроциты центральной нервной системы, по выходе из митоза еще в эмбриональном периоде, в дальнейшем не делятся. Жизненный цикл таких неделящихся клеток состоит из следующих периодов: митоза, роста, длительного функционирования, старения, смерти. Однако на протяжении длительного жизненного цикла такие клетки постоянно регенерируют по внутриклеточному типу: белковые и липидные молекулы, входящие в разнообразные структурные компоненты клеток, постепенно заменяются новыми, а следовательно такие клетки постепенно обновляются. Вместе с тем на протяжении жизненного цикла в цитоплазме неделящихся клеток постепенно накапливаются различные, прежде всего липидные включения, в частности липофусцин, который рассматривается как пигмент старения. Обычно клеточный рост сопровождается преимущественным увеличением объема цитоплазмы, тогда как величина ядра изменяется в меньшей степени. На протяжении онтогенеза строение клеток непрерывно изменяется. Прогрессирующие изменения клеток, связанные с появлением морфологических и функциональных различий между ранее индифферентными (однородными) клетками и обусловленные их специализацией в процессе развития, называют диф-ференцировкой клеток. Биохимической основой этого процесса является синтез специфических белков и других веществ. Молекулярные основы синтеза белков складываются из транскрипции первичной структуры матричной РНК на основе информации ДНК-гена, кодирующей области генов — экзонные области; процессинга мРНК, в результате которого из новообразованной цепи удаляются несмысловые последовательности нуклеотидов (интроны), перехода новообразованной мРНК в цитоплазму и трансляции — синтеза белка на аппарате синтеза белков клетки.  Дифференцировка сопровождается качественными, количественными и временными параметрами, т. е. характеризуется изменениями клеточной структуры, темпом развития (ускоренная или замедленная) и степенью (малодифференцированные — высокодифференцированные клетки). Усложнение структуры клетки сопровождается следующими изменениями: приобретением определенной формы и размеров ядра и клетки; сдвигом ядерно-цитоплазменного отношения в связи с более значительным ростом цитоплазмы по сравнению с ядром; развитием орга-нелл; образованием специализированных клеточных структур; синтезом специфических включений; образованием межклеточного вещества; появлением межклеточных взаимодействий и установлением межклеточных и специализированных контактов. Так, эпителиальные клетки приобретают кубическую, призматическую или плоскую форму. Клетки тканей внутренней среды более разнообразны по форме. Некоторые соединительнотканные клетки вырабатывают межклеточное вещество. Мышечные клетки содержат миофибриллы. Между нейронами формируются синаптические контакты (подробно — см. соответствующие главы). Для разных тканевых клеток характерны определенные взаимоотношения между процессами дифференцировки и деления. Однако в целом по мере повышения степени дифференцировки способность клеток к делению закономерно уменьшается. В гистогенезе клетки определенного цитотипа интегрируются, частично теряя автономность, присущую ранней стадии (пролиферативной), вследствие формирования регуляторных механизмов, оказывающих влияние на цитодифференцировку. Клетки в составе тканей многоклеточных организмов имеют различную продолжительность жизни. В некоторых тканях, например в эпителиях, смена клеточного состава происходит довольно быстро. По данным ряда исследователей, жизнь эпителиальных клеток (в частности, в кишечном эпителии) измеряется несколькими сутками. В других тканях (например, в мышечной и нервной) продолжительность жизни клеток сравнима с продолжительностью жизни организма. Так или иначе, клеточный состав организма не остается постоянным, он непрерывно изменяется в результате гибели части клеток и замены их новыми клеточными поколениями. В нормальных физиологических условиях гибели клеток предшествуют процессы старения. Они сопровождаются снижением функциональной активности, а также появлением ряда дистрофических изменений. В общих чертах гибель клеток характеризуется сморщиванием ядра и клетки в целом, диффузной окрашиваемостью и исчезновением специализированных структур, распадом клетки на фрагменты, которые фагоцитируются соседними клетками или макрофагами. Апоптоз – это тип гибели клеток, при котором сама клетка активно участвует в процессе своей гибели, т.е. происходит самоуничтожение клетки. Апоптоз, в отличие от некроза, является процессом активным, после воздействия этиологических факторов запускается генетически запрограммированный каскад реакций, сопровождающийся активацией определенных генов, синтезом белков, ферментов, приводящих к эффективному и быстрому удалению клетки из ткани. Биологическое значение апоптозаРезультатом апоптоза является постепенное и медленное избавление от "ненужных" в функциональном отношении на данный момент клеток. При этом не развивается воспаление и не нарушается нормальное функционирование соседних клеток, а также не происходит соединительнотканного замещения, что позволяет сохранить структуру органа. Функциональные элементы клетки, находящейся в состоянии апоптоза, не разрушаются, а поглощаются другими клетками и могут использоваться дальше. Особенно большую роль апоптоз играет в эмбриогенезе, когда важно постепенно избавляться от выполнивших свою функцию клеток, а активное фагоцитирование с развитием реакции воспаления может нарушить созревание плода. Апоптоз активно включается в развитие той или иной морфофункциональной системы организма. Наиболее ярко это можно продемонстрировать на примере созревания иммунной системы. На начальном этапе все иммунокомпетентные клетки проходят "обучение" в тимусе и лимфатических узлах, при этом каждый клон клеток приобретает способность распознавать определенный антиген. В ходе этого процесса возможно "патологическое научение" с последующим распознаванием как чужеродных антигенов своего организма и формированием иммунного ответа на них. В данном случае апоптоз является защитным механизмом, уничтожающим ставшие опасными клетки. В то же время лимфоцитарные клоны, распознающие антигены, не встречающиеся в течение жизни человека, не имеют функционального значения и апоптозируют. Апоптоз необходим также для элиминации клеток, выполнивших на определенном этапе развития свое функциональное значение и ставших ненужными. Помимо этого, апоптоз активно включается в процессы уничтожения клеток, подвергшихся мутации; в большей степени это относится к активно делящимся тканям (кроветворная, лимфатическая система и др.). Ответ 24! Митотический цикл. Фазы цикла (интерфаза, митоз). Биологическое значение митоза и его механизм. Преобразование структурных компонентов клетки на различных этапах митоза. Роль клеточного центра в митотическом делении клеток. Морфология митотических хромосом. Мейоз. Его механизм и биологическое значение. Важнейшим компонентом клеточного цикла является митотический (пролиферативный) цикл. Он представляет собой комплекс взаимосвязанных и согласованных явлений во время деления клетки, а также до и после него. Митотический цикл — это совокупность процессов, происходящих в клетке от одного деления до следующего и заканчивающихся образованием двух клеток следующей генерации. Кроме этого, в понятие жизненного цикла входят также период выполнения клеткой своих функций и периоды покоя. В это время дальнейшая клеточная судьба неопределенна: клетка может начать делиться (вступает в митоз) либо начать готовиться к выполнению специфических функций. Основные стадии митоза. 1.Редупликация (самоудвоение) генетической информации материнской клетки и равномерное распределение ее между дочерними клетками. Это сопровождается изменениями структуры и морфологии хромосом, в которых сосредоточено более 90% информации эукариотической клетки. 2.Митотический цикл состоит из четырех последовательных периодов: пресинтетического (или постмитотического) G1, синтетического S, постсинтетического (или премитотического) G2 и собственно митоза. Они составляют автокаталитическую интерфазу (подготовительный период). Фазы клеточного цикла: 1) пресинтетическая (G1). Идет сразу после деления клетки. Синтеза ДНК еще не происходит. Клетка активно растет в размерах, запасает вещества, необходимые для деления: белки (гистоны, структурные белки, ферменты), РНК, молекулы АТФ. Происходит деление митохондрий и хлоропластов (т. е. структур, способных к ауторепродукции). Восстанавливаются черты организации интерфазной клетки после предшествующего деления; 2) синтетическая (S). Происходит удвоение генетического материала путем репликации ДНК. Она происходит полуконсервативным способом, когда двойная спираль молекулы ДНК расходится на две цепи и на каждой из них синтезируется комплементарная цепочка. В итоге образуются две идентичные двойные спирали ДНК, каждая из которых состоит из одной новой и старой цепи ДНК. Количество наследственного материала удваивается. Кроме этого, продолжается синтез РНК и белков. Также репликации подвергается небольшая часть митохонд-риальной ДНК (основная же ее часть реплицируется в G2 период); 3) постсинтетическая (G2). ДНК уже не синтезируется, но происходит исправление недочетов, допущенных при синтезе ее в S период (репарация). Также накапливаются энергия и питательные вещества, продолжается синтез РНК и белков (преимущественно ядерных). S и G2 непосредственно связаны с митозом, поэтому их иногда выделяют в отдельный период — препрофазу. После этого наступает собственно митоз, который состоит из четырех фаз. Процесс деления включает в себя несколько последовательных фаз и представляет собой цикл. Его продолжительность различна и составляет у большинства клеток от 10 до 50 ч. При этом у клеток тела человека продолжительность самого митоза составляет 1—1,5 ч, G2-периода интерфазы — 2—3 ч, S-периода интерфазы — 6—10 ч. Стадии митоза. Процесс митоза принято подразделять на четыре основные фазы: профазу, метафазу, анафазу и телофазу (рис. 1–3). Так как он непрерывен, смена фаз осуществляется плавно — одна незаметно переходит в другую. В профазе увеличивается объем ядра, и вследствие спирализации хроматина формируются хромосомы. К концу профазы видно, что каждая хромосома состоит из двух хроматид. Постепенно растворяются ядрышки и ядерная оболочка, и хромосомы оказываются беспорядочно расположенными в цитоплазме клетки. Центриоли расходятся к полюсам клетки. Формируется ахроматиновое веретено деления, часть нитей которого идет от полюса к полюсу, а часть — прикрепляется к центромерам хромосом. Содержание генетического материала в клетке остается неизменным (2n2хр). В метафазе хромосомы достигают максимальной спирализации и располагаются упорядоченно на экваторе клетки, поэтому их подсчет и изучение проводят в этот период. Содержание генетического материала не изменяется (2n2хр). В анафазе каждая хромосома «расщепляется» на две хроматиды, которые с этого момента называются дочерними хромосомами. Нити веретена, прикрепленные к центромерам, сокращаются и тянут хроматиды (дочерние хромосомы) к противоположным полюсам клетки. Содержание генетического материала в клетке у каждого полюса представлено диплоидным набором хромосом, но каждая хромосома содержит одну хроматиду (2nlxp). В телофазе расположившиеся у полюсов хромосомы деспирализуются и становятся плохо видимыми. Вокруг хромосом у каждого полюса из мембранных структур цитоплазмы формируется ядерная оболочка, в ядрах образуются ядрышки. Разрушается веретено деления. Одновременно идет деление цитоплазмы. Дочерние клетки имеют диплоидный набор хромосом, каждая из которых состоит из одной хроматиды (2n1хр). Биологическое значение митоза. Оно состоит в том, что митоз обеспечивает наследственную передачу признаков и свойств в ряду поколений клеток при развитии многоклеточного организма. Благодаря точному и равномерному распределению хромосом при митозе все клетки единого организма генетически одинаковы. Митотическое деление клеток лежит в основе всех форм бесполого размножения как у одноклеточных, так и у многоклеточных организмов. Митоз обусловливает важнейшие явления жизнедеятельности: рост, развитие и восстановление тканей и органов и бесполое размножение организмов. В каждой фазе происходят определенные структурные преобразования. Профаза характеризуется морфологическими изменениями ядра и цитоплазмы. В ядре происходит: конденсация хроматина и образование хромосом, состоящих из двух хроматид, исчезновение ядрышка, распад кариолеммы на отдельные пузырьки. В цитоплазме отмечается редупликация (удвоение) центриолей и расхождение их к противоположным полюсам клетки, формирование из микротрубочек веретена деления, репродукция зернистой эндоплазматической сети, а также уменьшение числа свободных и прикрепленных рибосом. В метафазе происходит образование метафазной пластинки, или материнской звезды, неполное обособление сестринских хроматид друг от друга. Анафаза характеризуется полным обособлением (расхождением) хроматид и образованием двух равноценных диплоидных наборов хромосом, расхождением хромосомных наборов к полюсам митотического веретена и расхождением самих полюсов. Телофаза характеризуется деконденсацией хромосом каждого хромосомного набора, формированием из пузырьков ядерной оболочки, цитотомиейперетяжкой двуядерной клетки на две дочерние самостоятельные клетки, появлением ядрышка в ядрах дочерних клеток. В профазе хорошо видны центриоли - образования, находящиеся в клеточном центре и играющие роль в делении дочерних хромосом животных. (Напомним, что у высших растений нет центриолей в клеточном центре, который организует деление хромосом.) Мы же рассмотрим митоз на примере животной клетки, поскольку присутствие центриолей делает процесс деления клетки более наглядным. Центриоли делятся и расходятся к разным полюсам клетки. От центриолей протягиваются микротрубочки, образующие нити веретена деления, которое регулирует расхождение хромосом к полюсам делящийся клетки. Морфологию митотических хромосом лучше всего изучать в момент их наибольшей конденсации, в метафазе и в начале анафазы. Хромосомы в этом состоянии представляют собой палочковидные структуры разной длины с довольно постоянной толщиной. У большинства хромосом удается легко найти зону первичной перетяжки (центромеры), которая делит хромосому на два плеча (рис. 22). Хромосомы с равными или почти равными плечами называют метацентрическими, с плечами неодинаковой длины — субметацентрическими. Палочковидные хромосомы с очень коротким, почти незаметным вторым плечом называют акроцентрическими. В области первичной перетяжки расположен кинетохор. От этой зоны во время митоза отходят микротрубочки клеточного веретена, связанные с перемещением хромосом при делении клетки. Некоторые хромосомы имеют, кроме того, вторичные перетяжки, располагающиеся вблизи одного из концов хромосомы и отделяющие маленький участок — спутник хромосомы. Вторичные перетяжки называют, кроме того, ядрышковыми организаторами (см. предыдущую лекцию), так как именно на этих участках хромосом в интерфазе происходит образование ядрышка. В этих местах локализована ДНК, ответственная за синтез рибосомных РНК. Плечи хромосом оканчиваются теломерами — конечными участками. Теломерные участки хромосом характеризуются отсутствием способности к соединению с другими хромосомами или их фрагментами и выполняют защитную функцию. В каждом цикле деления теломеры клетки укорачивается, из-за неспособности ДНК-полимеразы синтезировать копию ДНК с самого конца. Данный феномен носит название концевой недорепликации и является одним из важнейших факторов биологического старения. Специальный фермент теломераза при помощи собственной РНК-матрицы достраивает теломерные повторы и удлиняет теломеры. В большинстве дифференцированных клеток теломераза заблокирована, однако активна в стволовых и половых клетках. Мейоз Центральным событием гаметогенеза является особая форма клеточного деления — мейоз. В отличие от широко распространенного митоза, сохраняющего в клетках постоянное диплоидное число хромосом, мейоз приводит к образованию из диплоидных клеток гаплоидных гамет. При последующем оплодотворении гаметы формируют организм нового поколения с диплоидным кариотипом (пс + пс == 2n2c). В этом заключается важнейшее биологическое значение мейоза, который возник и закрепился в процессе эволюции у всех видов, размножающихся половьм путем (см. разд. 3.6.2.2). Мейоз состоит из двух быстро следующих одно за другим делений, происходящих в периоде созревания. Удвоение ДНК для этих делений осуществляется однократно в периоде роста. Второе деление мейоза следует за первым практически сразу так, что наследственный материал не синтезируется в промежутке между ними (рис. 5.5). Первое мейотическое деление называют редукционным, так как оно приводит к образованию из диплоидных клеток (2п2с) гаплоидных клеток п2с. Такой результат обеспечивается благодаря особенностям профазы первого деления мейоза. В профазе I мейоза, так же как в обычном митозе, наблюдается компактная упаковка генетического материала (спирализация хромосом). Одновременно происходит событие, отсутствующее в митозе: гомологичные хромосомы конъюгируют друг с другом, т.е. тесно сближаются соответствующими участками. В результате конъюгации образуются хромосомные пары, или биваленты, числом п. Так как каждая хромосома, вступающая в мейоз, состоит из двух хроматид, то бивалент содержит четыре хроматиды. Формула генетического материала в профазе I остается 2n4c. К концу профазы хромосомы в бивалентах, сильно спирализуясь, укорачиваются. Так же как в митозе, в профазе I мейоза начинается формирование веретена деления, с помощью которого хромосомный материал будет распределяться между дочерними клетками (рис. 5.5). 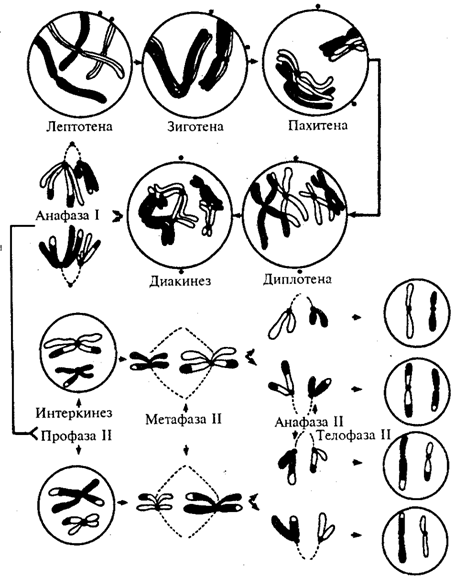 Рис. 5.5. Стадии мейоза Отцовские хромосомы обозначены черным цветом, материнские —неокрашснные. На рисунке не изображена метафаза I, в которой биваленты располагаются в плоскости экватора веретена деления, и телофаза I, быстро переходящая в профазу II Процессы, происходящие в профазе I мейоза и определяющие его результаты, обусловливают более продолжительное течение этой фазы деления по сравнению с митозом и дают возможность выделить несколько стадий в ее пределах (рис. 5.5). Лептотена —наиболее ранняя стадия профазы I мейоза, в которой начинается спирализация хромосом, и они становятся видимыми в микроскоп как длинные и тонкие нити. Зиготена характеризуется началом конъюгации гомологичных хромосом, которые объединяются синаптонемальным комплексом в бивалент (рис. 5.6). Пахитена — стадия, в которой на фоне продолжающейся спирализации хромосом и их укорочения, между гомологичными хромосомами осуществляется кроссинговер — перекрест с обменом соответствующими участками. Диплотена характеризуется возникновением сил отталкивания между гомологичными хромосомами, которые начинают отдаляться друг от друга в первую очередь в области центромер, но остаются связанными в областях прошедшего кроссинговера — хиазмах (рис. 5.7). Диакинез — завершающая стадия профазы I мейоза, в которой гомологичные хромосомы удерживаются вместе лишь в отдельных точках хиазм. Биваленты приобретают причудливую форму колец, крестов, восьмерок и т.д. (рис. 5.8). Таким образом, несмотря на возникающие между гомологичными хромосомами силы отталкивания, в профазе I не происходит окончательного разрушения бивалентов. Особенностью мейоза в овогенезе является наличие специальной стадии — диктиотены, отсутствующей в сперматогенезе. На этой стадии, достигаемой у человека еще в эмбриогенезе, хромосомы, приняв особую морфологическую форму «ламповых щеток», прекращают какие-либо дальнейшие структурные изменения на многие годы. По достижении женским организмом репродуктивного возраста под влиянием лютеинизирующего гормона гипофиза, как правило, один овоцит ежемесячно возобновляет мейоз. В метафазе I мейоза завершается формирование веретена деления. Его нити прикрепляются к центромерам хромосом, объединенных в биваленты, таким образом, что от каждой центромеры идет лишь одна нить к одному из полюсов веретена. В результате нити, связанные с центромерами гомологичных хромосом, направляясь к разным полюсам, устанавливают бивалентны в плоскости экватора веретена деления. В анафазе I мейоза ослабляются связи между гомологичными хромосомами в бивалентах и они отходят друг от друга, направляясь к разным полюсам веретена деления. При этом к каждому полюсу отходит гаплоидный набор хромосом, состоящих из двух хроматид (см. рис. 5.5). В телофазе I мейоза у полюсов веретена собирается одинарный, гаплоидный набор хромосом, каждая из них содержит удвоенное количество ДНК. Формула генетического материала образующихся дочерних клеток соответствует п2с. Второе мейотическое (эквационное) деление приводит к образованию клеток, в которых содержание генетического материала в хромосомах будет соответствовать их однонитчатой структуре пс (см. рис. 5.5). Это деление протекает, как митоз, только клетки, вступающие в него, несут гаплоидный набор хромосом. В процессе такого деления материнские двунитчатые хромосомы, расщепляясь, образуют дочерние однонитчатые. Одна из главных задач мейоза — создание клеток с гаплоидным набором однонитчатых хромосом —достигается благодаря однократной редупликации ДНК для двух последовательных делений мейоза, а также благодаря образованию в начале первого мейотического деления пар гомологичных хромосом и дальнейшего их расхождения в дочерние клетки. Процессы, протекающие в редукционном делении, обеспечивают также не менее важное следствие — генетическое разнообразие гамет, образуемых организмом. К таким процессам относят кроссинговер, расхождение гомологичных хромосом в разные гаметы и независимое поведение бивалентов в первом мейотическом делении (см. разд. 3.6.2.3). Кроссинговер обеспечивает перекомбинацию отцовских и материнских аллелей в группах сцепления (см. рис. 3.72). Ввиду того что перекрест хромосом может происходить в разных участках, кроссинговер в каждом отдельном случае приводит к обмену разным по количеству генетическим материалом. Необходимо отметить также возможность возникновения нескольких перекрестов между двумя хроматидами (рис. 5.9) и участия в обмене более чем двух хроматид бивалента (рис. 5.10). Отмеченные особенности кроссинговера делают этот процесс эффективным механизмом перекомбинации аллелей. Расхождение гомологичных хромосом в разные гаметы в случае гетерозиготности приводит к образованию гамет, различающихся по аллелям отдельных генов (см. рис. 3.74). Случайное расположение бивалентов в плоскости экватора веретена деления и последующее их расхождение в анафазе I мейоза обеспечивают перекомбинацию родительских групп сцепления в гаплоидном наборе гамет (см. рис. 3.75). Ответ 25! Ткани как системы клеток и их производных - один из иерархических уровней организации живого. Клетки как ведущие элементы ткани. Неклеточные структуры - симпласты и межклеточное вещество как производные клеток. Синцитии. Понятие о клеточных популяциях. Диффероны. Закономерности возникновения и эволюции тканей, теории параллелизма А.А.Заварзина и дивергентной эволюции Н.Г.Хлопина, их синтез на современном уровне развития науки. Ткань – исторически (филогенетически) сложившаяся система клеток и неклеточных структур, обладающая общностью строения, а иногда и происхождения и специализированная на выполнении определенных функций. Ткань – это новый (после клеток) уровень организации живой материи. Структурные компоненты ткани: клетки, производные клеток, межклеточное вещество. Характеристика структурных компонентов ткани Клетки – основные, функционально ведущие компоненты тканей. Практически все ткани состоят из нескольких типов клеток. Кроме того, клетки каждого типа в тканях могут находиться на разных этапах зрелости (дифференцировки). Поэтому в ткани различают такие понятия, как клеточная популяция и клеточный дифферон. Клеточная популяция – это совокупность клеток данного типа. Например, в рыхлой соединительной ткани (самой распространенной в организме) содержится: 1) популяция фибробластов; 2) популяция макрофагов; 3) популяция тканевых базофилов и др. Клеточный дифферон (или гистогенетический ряд) – это совокупность клеток данного типа (данной популяция), находящихся на различных этапах дифференцировки. Исходными клетками дифферона являются стволовые клетки, далее идут молодые (бластные) клетки, созревающие клетки и зрелые клетки. Различают полный дифферон или неполный в зависимости от того, находятся ли в тканях клетки всех типов развития. Однако ткани – это не просто скопление различных клеток. Клетки в тканях находятся в определенной взаимосвязи, и функция каждой из них направлена на выполнение функции ткани. Клетки в тканях оказывают влияние друг на друга или непосредственно через щелевидные контакты (нексусы) и синапсы, или на расстоянии (дистантно) посредством выделения различных биологически активных веществ. Производные клеток: 1) симпласты (слияние отдельных клеток, например мышечное волокно); 2) синцитий (несколько клеток, соединенных между собой отростками, например сперматогенный эпителий извитых канальцев семенника); 3) постклеточные образования (эритроциты, тромбоциты). Межклеточное вещество – также продукт деятельности определенных клеток. Межклеточное вещество состоит из: 1) аморфного вещества; 2) волокон (коллагеновых, ретикулярных, эластических). Межклеточное вещество неодинаково выражено в разных тканях. Развитие тканей в онтогенезе (эмбриогенезе) и филогенезе В онтогенезе различают следующие этапы развития тканей: 1) этап ортотопической дифференцировки. На этом этапе зачатки будущих определенных тканей локализуются сначала в определенных участках яйцеклетки и затем – зиготы; 2) этап бластомерной дифференцировки. В результате дробления зиготы презумптивные (предположительные) зачатки тканей оказываются локализованными в разных бластомерах зародыша; 3) этап зачатковой дифференцировки. В результате гаструляции предположительные зачатки тканей локализуются в определенных участках зародышевых листков; 4) гистогенез. Это процесс преобразования зачатков тканей и ткани в результате пролиферации, роста, индукции, детерминации, миграции и дифференцировки клеток. Имеется несколько теорий развития тканей в филогенезе: 1) закон параллельных рядов (А. А. Заварзин). Ткани животных и растений разных видов и классов, выполняющие одинаковые функции, имеют сходное строение, т. е. развиваются они параллельно у животных различных филогенетических классов; 2) закон дивергентной эволюции (Н. Г. Хлопин). В филогенезе происходит расхождение признаков тканей и появление новых разновидностей ткани в пределе тканевой группы, что приводит к усложнению животных организмов и появлению разнообразия тканей. Онтогенез. Понятия детерминации и коммитирования. Развитие организма начинается с одноклеточной стадии — зиготы. В ходе дробления возникают бластомеры, но совокупность бластомеров – это еще не ткань. Бластомеры на начальных этапах дробления еще не детерминированы (они тотипотентны). Если отделить их один от другого, - каждый может дать начало полноценному самостоятельному организму – механизм возникновения монозиготных близнецов. Постепенно на следующих стадиях происходит ограничение потенций. В основе его лежат процессы, связанные с блокированием отдельных компонентов генома клеток и детерминацией. Детерминация – это процесс определения дальнейшего пути развития клеток на основе блокирования отдельных генов. Понятие «коммитирование» тесно связано с клеточным делением (т.н. коммитирующий митоз). Коммитирование – это ограничение возможных путей развития вследствие детерминации. Коммитирование совершается ступенчато. Сначала соответствующие преобразования генома касаются крупных его участков. Затем все более детализируются, поэтому вначале детерминируются наиболее общие свойства клеток, а затем и более частные. Как известно, на этапе гаструляции возникают эмбриональные зачатки. Клетки, которые входят в их состав, еще не окончательно детерминированы, так что из одного зачатка возникают клеточные совокупности, обладающие разными свойствами. Следовательно, один эмбриональный зачаток может служить источником развития нескольких тканей. ТЕОРИЯ ЭВОЛЮЦИИ ТКАНЕЙПоследовательная ступенчатая детерминация и коммитирование потенций однородных клеточных группировок — дивергентный процесс. В общем виде эволюционная концепция дивергентного развития тканей в филогенезе и в онтогенезе была сформулирована Н.Г.Хлопиным. Современные генетические концепции подтверждают правоту его представлений. Именно Н.Г.Хлопин ввел понятие о генетических тканевых типах. Концепция Хлопина хорошо отвечает на вопрос, как и какими путями происходило развитие и становление тканей, но не останавливается на причинах, определяющих пути развития. Причинные аспекты развития тканей раскрывает теория параллелизмов А.А.Заварзина. Он обратил внимание на сходство строения тканей, которые выполняют одинаковые функции у животных, принадлежащих даже к весьма удаленным друг от друга эволюционным группировкам. Вместе с тем известно, что, когда эволюционные ветви только расходились, у общих предков таких специализированных тканей еще не было. Следовательно, в ходе эволюции в разных ветвях филогенетического древа самостоятельно, как бы параллельно, возникали одинаково организованные ткани, выполняющие сходную функцию. Причиной этого является естественный отбор: если возникали какие-то организмы, у которых соответствие строения и функции клеток, тканей, органов нарушалось, они были и менее жизнеспособны. Теория Заварзина отвечает на вопрос, почему развитие тканей шло тем, а не иным путем, раскрывает казуальные аспекты эволюции тканей. Концепции А.А.Заварзина и Н.Г.Хлопина, разработанные независимо одна от другой, дополняют друг друга и были объединены А.А.Брауном и В.П.Михайловым: сходные тканевые структуры возникали параллельно в ходе дивергентного развития. Ответ 26! Принципы классификации тканей. Классификация тканей. Компенсаторно-приспособительные и адаптационные изменения тканей, их пределы. Принятое деление тканей организма на четыре основных типа в ряде отношений неудовлетворительно. Здесь следует сказать, что критерием для выделения этих четырех типов служило не происхождение тканей из трех зародышевых листков в процессе эмбриогенеза, а главным образом их микроскопическое строение и выполняемые ими функции. Оказалось, однако, что большинство покровных тканей развивается из эктодермы и энтодермы, хотя некоторые из них мезодермального происхождения. Соединительная ткань образуется из мезодермы. Мышечная ткань почти целиком происходит из мезодермы, за исключением небольшой ее части, развивающейся из эктодермы. Нервная ткань имеет эктодермальное происхождение. Но, как мы увидим в дальнейшем, то, что принято называть мышечной тканью, на самом деле состоит из смеси мышечных волокон и соединительной ткани. Подобным же образом в некоторых участках нервной ткани к структурам эктодермального происхождения примешана соединительная ткань, происходящая из мезодермы. Тем не менее принятая классификация тканей, несмотря на свое несовершенство, чрезвычайно полезна. |