Пособие по биологии для поступающих в вузы Авторы Пименов А. В., Гончаров О. В
 Скачать 34.81 Mb. Скачать 34.81 Mb.
|

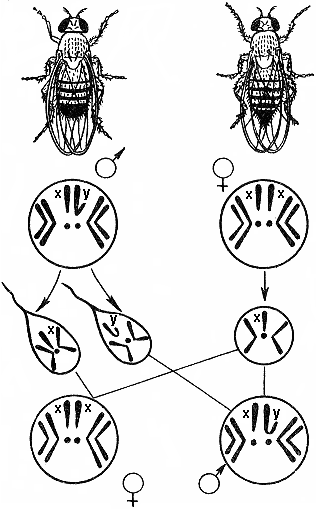
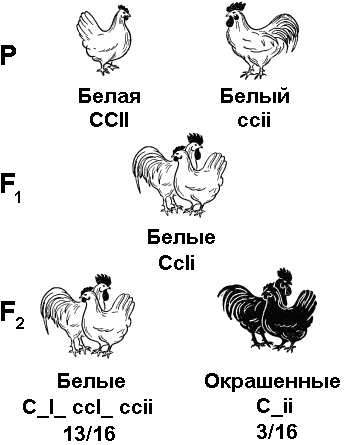
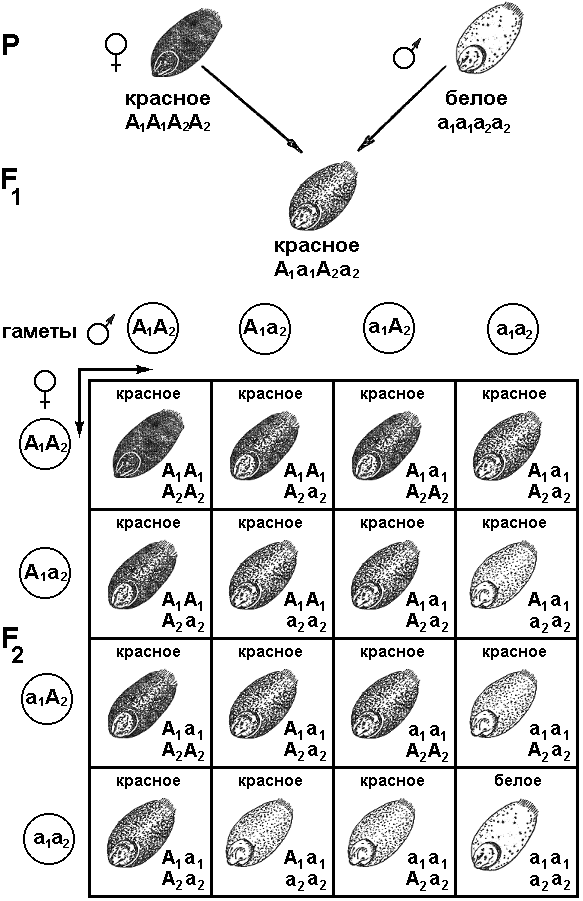
4 Как известно, большинство животных и двудомных растений являются раздельнополыми организмами, причем внутри вида количество особей мужского пола приблизительно равно количеству особей женского пола. Пол можно рассматривать как один из признаков организма. Наследование признаков организма, как правило, определяется генами. Механизм же определения пола имеет иной характер — хромосомный (рис. 328). Пол чаще всего определяется в момент оплодотворения. У человека женский пол является гомогаметным, то есть все яйцеклетки несут Х-хромосому. Мужской организм — гетерогаметен, то есть образует два типа гамет — 50% гамет несет Х-хромосому и 50% — Y-хромосому. Если образуется зигота, несущая две Х-хромосомы, то из нее будет формироваться женский организм, если Х-хромосому и Y-хромосому — мужской. С 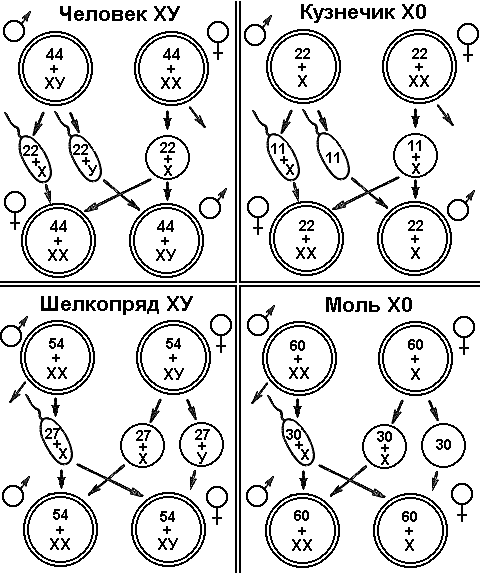 Рис. 329. Основные типы хромосомного определения пола. оотношение полов, близкое к расщеплению 1:1, соответствует расщеплению при анализирующем скрещивании. Поскольку женский организм имеет две одинаковые половые хромосомы, его можно рассматривать как гомозиготный, мужской, образующий два типа гамет — как гетерозиготный. Из приведенной схемы видно, как происходит формирование в равных количествах двух групп особей, отличающихся набором половых хромосом. Существует четыре основных типа хромосомного определения пола (рис. 329):
40.5. Наследование признаков, | ||||||||||||||||||||||||
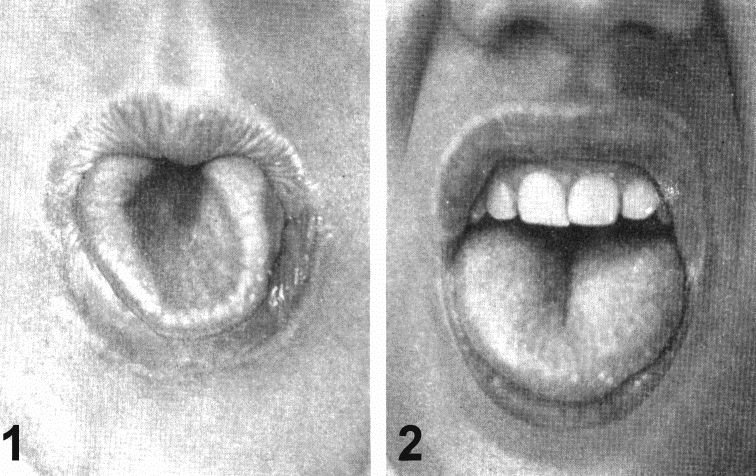
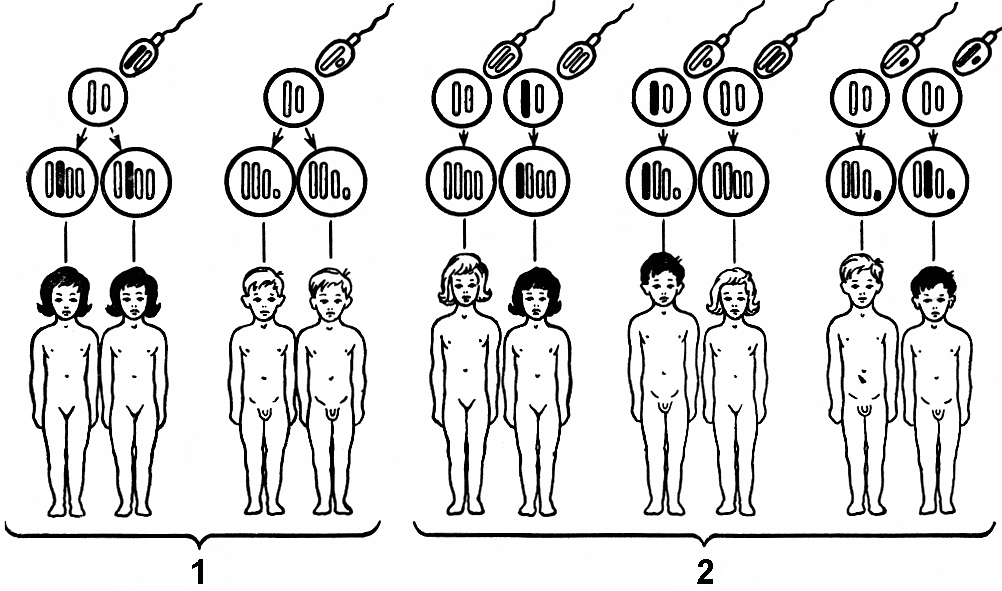
| Признаки | Конкордантность, % | |
| | Монозиготные близнецы | Дизиготные близнецы |
| Нормальные | ||
| Группа крови (АВ0) Цвет глаз Цвет волос | 100 99,5 97 | 46 28 23 |
| Признаки | Конкордантность, % | |
| | Монозиготные близнецы | Дизиготные близнецы |
| Патологические | ||
| Косолапость "Заячья губа" Бронхиальная астма Корь Туберкулез Эпилепсия Шизофрения | 32 33 19 98 37 67 70 | 3 5 4,8 94 15 3 13 |
Как видно из таблицы, степень коркондантности монозиготных близнецов по всем приведенным признакам значительно выше, чем у дизиготных, однако она не является абсолютной. Как правило, дискордантность однояйцевых близнецов возникает в результате нарушений внутриутробного развития одного из них или под влиянием внешней среды, если она была разной.
Благодаря близнецовому методу, была выяснена наследственная предрасположенность человека к ряду заболеваний: шизофрении, умственной отсталости, эпилепсии, сахарного диабета и других.
Наблюдения за однояйцевыми близнецами дают материал для выяснения роли наследственности и среды в развитии признаков. Причем под внешней средой понимают не только физические факторы среды, но и
социальные условия.
Цитогенетический метод основан на изучении хромосом человека в норме и при патологии. В норме кариотип человека включает 46 хромосом — 22 пары аутосом и две половые хромосомы. Использование данного метода позволило выявить группу болезней, связанных либо с изменением числа хромосом, либо с изменениями их структуры. Такие болезни получили название хромосомных. К их числу относятся: синдром Клайнфельтера, синдром Шерешевского-Тернера, трисомия Х, синдром Дауна, синдром Патау, синдром Эдвардса и другие.
Больные с синдромом Клайнфельтера (47,ХХУ) всегда мужчины. Они характеризуются недоразвитием половых желез, дегенерацией семенных канальцев, часто умственной отсталостью, высоким ростом (за счет непропорционально длинных ног).
Синдром Шерешевского-Тернера (45,Х0) наблюдается у женщин. Он проявляется в замедлении полового созревания, недоразвитии половых желез, аменорее (отсутствии менструаций), бесплодии. Женщины с синдромом Шерешевского-Тернера имеют малый рост, тело диспропорционально — более развита верхняя часть тела, плечи широкие, таз узкий — нижние конечности укорочены, шея короткая со складками, "монголоидный" разрез глаз и ряд других признаков.
Синдром Дауна — одна из самых часто встречающихся хромосомных болезней. Она развивается в результате трисомии по 21 хромосоме (47, 21,21,21). Болезнь легко диагностируется, так как имеет ряд характерных признаков: укороченные конечности, маленький череп, плоское, широкое переносье, узкие глазные щели с косым разрезом, наличие складки верхнего века, психическая отсталость. Часто наблюдаются и нарушения строения внутренних органов.
Хромосомные болезни возникают и в результате изменения самих хромосом. Так, делеция 5-й хромосомы приводит к развитию синдрома "крик кошки". У детей с этим синдромом нарушается строение гортани, и они в раннем детстве имеют своеобразный "мяукающий" тембр голоса. Кроме того, наблюдается отсталость психомоторного развития и слабоумие. Делеция 21 хромосомы приводит к возникновению одной из форм белокровия.
Чаще всего хромосомные болезни являются результатом мутаций, произошедших в половых клетках одного из родителей.
Биохимический метод позволяет обнаружить нарушения в обмене веществ, вызванные изменением генов и, как следствие, изменением активности различных ферментов. Наследственные болезни обмена веществ подразделяются на болезни углеводного обмена (сахарный диабет), обмена аминокислот, липидов, минералов и др.
Фенилкетонурия относится к болезням аминокислотного обмена. Блокируется превращение незаменимой аминокислоты фенилаланин в тирозин, при этом фенилаланин превращается в фенилпировиноградную кислоту, которая выводится с мочой. Заболевание приводит к быстрому развитию слабоумия у детей. Ранняя диагностика и диета позволяют приостановить развитие заболевания.
Генетика человека — одна из наиболее интенсивно развивающихся отраслей науки. Она является теоретической основой медицины, раскрывает биологические основы наследственных заболеваний. Знание генетической природы заболеваний позволяет вовремя поставить точный диагноз и осуществить нужное лечение.
40.8. Генетика популяций
Популяция — это совокупность особей одного вида, длительное время обитающих на определенной территории, свободно скрещивающихся друг с другом, имеющих общее происхождение, определенную генетическую структуру и в той или иной степени изолированных от других таких совокупностей особей данного вида. Популяция не только единица вида, форма его существования, но и единица эволюции. В основе микроэволюционных процессов, завершающихся видообразованием, лежат генетические преобразования в популяциях.
Изучением генетической структуры и динамики популяций занимается особый раздел генетики — популяционная генетика.
С генетической точки зрения, популяция является открытой системой, а вид — закрытой. В общей форме процесс видообразования сводится к преобразованию генетически открытой системы в генетически закрытую.
Каждая популяция имеет определенный генофонд и генетическую структуру. Генофондом популяции называют совокупность генотипов всех особей популяции. Под генетической структурой популяции понимают соотношение в ней различных генотипов и аллелей.
Одними из основных понятий популяционной генетики являются частота генотипа и частота аллеля. Под частотой генотипа (или аллеля) понимают его долю, отнесенную к общему количеству генотипов (или аллелей) в популяции. Частота генотипа, или аллеля, выражается либо в процентах, либо в долях единицы (если общее количество генотипов или аллелей популяции принимается за 100% или 1). Так, если ген имеет две аллельные формы и доля рецессивного аллеля а составляет ¾ (или 75%), то доля доминантного аллеля А будет равна ¼ (или 25%) общего числа аллелей данного гена в популяции.
Большое влияние на генетическую структуру популяций оказывает способ размножения. Например, популяции самоопыляющихся и перекрестноопыляющихся растений существенно отличаются друг от друга.
Впервые исследование генетической структуры популяции было предпринято В.Иоганнсеном в 1903 г. В качестве объектов исследования были выбраны популяции самоопыляющихся растений. Исследуя в течение нескольких поколений массу семян у фасоли, он обнаружил, что у самоопылителей популяция состоит из генотипически разнородных групп, так называемых чистых линий, представленных гомозиготными особями. Причем из поколения в поколение в такой популяции сохраняется равное соотношение гомозиготных доминантных и гомозиготных рецессивных генотипов. Их частота в каждом поколении увеличивается, в то время как частота гетерозиготных генотипов будет уменьшаться. Таким образом, в популяциях самоопыляющихся растений наблюдается процесс гомозиготизации, или разложения на линии с различными генотипами.
Большинство растений и животных в популяциях размножаются половым путем при свободном скрещивании, обеспечивающем равновероятную встречаемость гамет. Равновероятную встречаемость гамет при свободном скрещивании называют панмиксией, а такую популяцию — панмиктической.
Закон Харди-Вайнберга
В 1908 г. английский математик Г.Харди и немецкий врач Н.Вайнберг независимо друг от друга сформулировали закон, которому подчиняется распределение гомозигот и гетерозигот в панмиктической популяции, и выразили его в виде алгебраической формулы.Частоту встречаемости гамет с доминантным аллелем А обозначают p, а частоту встречаемости гамет с рецессивным аллелем а — q. Частоты этих аллелей в популяции выражаются формулой p +q = 1 (или 100%). Поскольку в панмиктической популяции встречаемость гамет равновероятна, можно определить и частоты генотипов.
Х
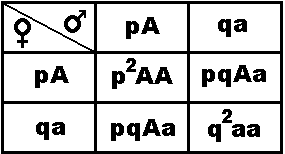
арди и Вайнберг, суммируя данные о частоте генотипов, образующихся в результате равновероятной встречаемости гамет, вывели формулу частоты генотипов в панмиктической популяции:
P2 + 2pq + q2 = 1.
АА + 2Аа + аа = 1
Пользуясь этими формулами, можно рассчитать частоты аллелей и генотипов в конкретной панмиктической популяции.
Однако действие этого закона выполняется при соблюдении следующих условий:
неограниченно большая численность популяции;
все особи могут свободно скрещиваться друг с другом;
все генотипы одинаково жизнеспособны, плодовиты и не подвергаются отбору;
прямые и обратные мутации возникают с одинаковой частотой или настолько редко, что ими можно пренебречь;
отток или приток новых генотипов в популяцию отсутствует.
В реально существующих популяциях выполнение этих условий невозможно, поэтому закон справедлив только для идеальной популяции. Несмотря на это, закон Харди-Вайнберга является основой для анализа некоторых генетических явлений, происходящих в природных популяциях. Например, если известно, что фенилкетонурия встречается с частотой 1:10000 и наследуется по аутосомно-рецессивному типу, можно посчитать частоту встречаемости гетерозигот и гомозигот по доминантному признаку. Больные фенилкетонурией имеют генотип q2(аа) = 0,0001. Отсюда q = 0,01. p = 1 — 0,01 = 0,99. Частота встречаемости гетерозигот равна 2pq, равна 2 х 0,99 х 0,01 ≈ 0,02 или около 2%. Частота встречаемости гомозигот по доминантному и рецессивному признакам: АА = p2 = 0,992 ≈ 98%, аа = 0,01%.
Факторы, изменяющие
генетическую структуру популяции
Изменение равновесия генотипов и аллелей в панмиктической популяции происходит под влиянием постоянно действующих факторов, к которым относятся:
мутационный процесс;
популяционные волны;
изоляция;
естественный отбор;
дрейф генов и другие.
Именно благодаря этим явлениям возникает элементарное эволюционное явление — изменение генетического состава популяции, являющееся начальным этапом процесса видообразования.
40.9. Изменчивость
Генетика изучает не только наследственность, но и изменчивость организмов. Изменчивостью называют способность живых организмов приобретать новые признаки и свойства. Благодаря изменчивости, организмы могут приспосабливаться к изменяющимся условиям среды обитания.
Различают два типа изменчивости:
наследственную, или генотипическую, — изменения признаков организма, обусловленные изменением генотипа; она бывает:
комбинативной — возникающей в результате перекомбинации хромосом в процессе полового размножения и участков хромосом в процессе кроссинговера;
мутационной — возникающей в результате внезапного изменения состояния генов;
ненаследственную, или фенотипическую, — изменчивость, при которой изменений генотипа не происходит.
Мутационная изменчивость
Наследственные изменения генетического материала теперь называют мутациями. Мутации — внезапные изменения генетического материала, приводящие к изменению тех или иных признаков организмов.Термин "мутация" впервые ввел в науку голландский генетик Г. де-Фриз. Проводя опыты с энотерой (декоративное растение), он случайно обнаружил экземпляры, отличающиеся рядом признаков от остальных (большой рост, гладкие, узкие и длинные листья, красные жилки листьев и широкая красная полоса на чашечке цветка и т.д.). Причем при семенном размножении растения из поколения в поколение стойко сохраняли эти признаки. В результате обобщения своих наблюдений де-Фриз создал мутационную теорию, основные положения которой не утратили своего значения и по сей день:
мутации возникают внезапно, скачкообразно, без всяких переходов;
мутации наследственны, т.е. стойко передаются из поколения в поколение;
мутации не образуют непрерывных рядов, не группируются вокруг среднего типа (как при модификационной изменчивости), они являются качественными изменениями;
мутации ненаправленны — мутировать может любой локус, вызывая изменения как незначительных, так и жизненно важных признаков в любом направлении;
одни и те же мутации могут возникать повторно;
мутации индивидуальны, то есть возникают у отдельных особей.
Процесс возникновения мутаций называют мутагенез, организмы, у которых произошли мутации, — мутантами, а факторы среды, вызывающие появление мутаций, — мутагенными.
Способность к мутированию — одно из свойств гена. Каждая отдельная мутация вызывается какой-то причиной, как правило, связанной с изменениями во внешней среде.
Классификация мутаций
Существует несколько классификаций мутаций:
Мутации по месту их возникновения:
Генеративные — возникшие в половых клетках. Они не влияют на признаки данного организма, а проявляются только в следующем поколении.
Соматические — возникающие в соматических клетках. Эти мутации проявляются у данного организма и не передаются потомству при половом размножении (черное пятно на фоне коричневой окраски шерсти у каракулевых овец). Сохранить соматические мутации можно только путем бесполого размножения (прежде всего вегетативного).
Мутации по адаптивному значению:
Полезные — повышающие жизнеспособность особей.
Вредные:
летальные — вызывающие гибель особей;
полулетальные — снижающие жизнеспособность особи (у мужчин рецессивный ген гемофилии носит полулетальный характер, а гомозиготные женщины оказываются нежизнеспособными).
Нейтральные — не влияющие на жизнеспособность особей.
Эта классификация весьма условна, так как одна и та же мутация в одних условиях может быть полезной, а в других — вредной.
Мутации по характеру проявления:
доминантные, которые могут делать обладателей этих мутаций нежизнеспособными и вызывать их гибель на ранних этапах онтогенеза (если мутации являются вредными);
рецессивные — мутации, не проявляющиеся у гетерозигот, поэтому длительное время сохраняющиеся в популяции и образующие резерв наследственной изменчивости (при изменении условий среды обитания носители таких мутаций могут получить преимущество в борьбе за существование).
Мутации по степени фенотипического проявления:
крупные — хорошо заметные мутации, сильно изменяющие фенотип (махровость у цветков);
малые — мутации, практически не дающие фенотипического проявления (незначительное удлинение остей у колоса).
Мутации по изменению состояния гена:
прямые — переход гена от дикого типа к новому состоянию29;
обратные — переход гена от мутантного состояния к дикому типу.
Мутации по характеру их появления:
спонтанные — мутации, возникшие естественным путем под действием факторов среды обитания;
индуцированные — мутации, искусственно вызванные действием мутагенных факторов.
Мутации по характеру изменения генотипа:
генны;
хромосомные;
геномные.
Мутации по характеру изменения генотипа
Мутации могут вызывать различные изменения генотипа, затрагивая отдельно взятые гены, целые хромосомы или весь геном.Генные мутации
Генными мутациями называют изменения структуры молекулы ДНК на участке определенного гена, кодирующего структуру определенной молекулы белка. Эти мутации влекут за собой изменение строения белков, то есть появляется новая последовательность аминокислот в полипептидной цепи, в результате чего происходит изменение функциональной активности белковой молекулы. Благодаря генным мутациям происходит возникновение серии множественных аллелей одного и того же гена. Чаще всего генные мутации происходят в результате:
замены одного или нескольких нуклеотидов на другие;
вставки нуклеотидов;
потери нуклеотидов;
удвоения нуклеотидов;
изменения порядка чередования нуклеотидов.
Хромосомные мутации
Хромосомные мутации — мутации, вызывающие изменения структуры хромосом. Они возникают в результате разрыва хромосом с образованием "липких" концов, "Липкие" концы — это одноцепочечные фрагменты на концах двухцепочечной молекулы ДНК. Эти фрагменты способны соединяться с другими фрагментами хромосом, также имеющих "липкие" концы. Перестройки могут осуществляться как в пределах одной хромосомы — внутрихромосомные мутации, так и между негомологичными хромосомами — межхромосомные мутации.
Внутрихромосомные мутации:
делеция — утрата части хромосомы (АВСD AB);
инверсия — поворот участка хромосомы на 180˚(ABCD ACBD);
дупликация — удвоение одного и того же участка хромосомы; (ABCD ABCBCD);
Межхромосомные мутации:
транслокация — обмен участками между негомологичными хромосомами (АВCD AB34).
Геномные мутации
Геномными называют мутации, в результате которых происходит изменение в клетке числа хромосом. Геномные мутации возникают в результате нарушения митоза или мейоза, приводящих либо к неравномерному расхождению хромосом к полюсам клетки, либо к удвоению хромосом, но без деления цитоплазмы.В зависимости от характера изменения числа хромосом, различают:
Гаплоидию — уменьшение числа полных гаплоидных наборов хромосом.
Полиплоидию — увеличение числа полных гаплоидных наборов хромосом. Полиплоидия чаще наблюдается у простейших и у растений. В зависимости от числа гаплоидных наборов хромосом, содержащихся в клетках, различают: триплоиды (3n), тетраплоиды (4n) и т.д. Они могут быть:
автополиплоидами — полиплоидами, возникающими в результате умножения геномов одного вида;
аллополиплоидами — полиплоидами, возникающими в результате умножения геномов разных видов (характерно для межвидовых гибридов).
Гетероплоидию (анеуплоидия) — некратное увеличение или уменьшение числа хромосом. Чаще всего наблюдается уменьшение или увеличение числа хромосом на одну (реже две и более). Вследствие нерасхождения какой-либо пары гомологичных хромосом в мейозе одна из образовавшихся гамет содержит на одну хромосому меньше, а другая — на одну больше. Слияние таких гамет с нормальной гаплоидной гаметой при оплодотворении приводит к образованию зиготы с меньшим или большим числом хромосом по сравнению с диплоидным набором, характерным для данного вида. Среди анеуплоидов встречаются:
трисомики — организмы с набором хромосом 2n+1;
моносомики — организмы с набором хромосом 2n -1;
нулесомики — организмы с набором хромосом 2n –2.
Например, болезнь Дауна у человека возникает в результате трисомии по 21-й паре хромосом.
Закон гомологических рядов в наследственной изменчивости
Н.И. Вавилов, изучая наследственную изменчивость у культурных растений и их предков, обнаружил ряд закономерностей, которые позволили сформулировать закон гомологических рядов наследственной изменчивости: «Виды и роды, генетически близкие, характеризуются сходными рядами наследственной изменчивости с такой правильностью, что, зная ряд форм в пределах одного вида, можно предвидеть нахождение параллельных форм у других видов и родов. Чем ближе генетически расположены в общей системе роды и виды, тем полнее сходство в рядах их изменчивости. Целые семейства растений в общем характеризуются определенным циклом изменчивости, проходящей через все роды и виды, составляющие семейство30».Этот закон можно проиллюстрировать на примере семейства Мятликовые, к которому относятся пшеница, рожь, ячмень, овес, просо и т.д. Так, черная окраска зерновки обнаружена у ржи, пшеницы, ячменя, кукурузы и других растений, удлиненная форма зерновки — у всех изученных видов семейства. Закон гомологических рядов в наследственной изменчивости позволили самому Н.И.Вавилову найти ряд форм ржи, ранее не известных, опираясь на наличие этих признаков у пшеницы. К ним относятся: остистые и безостые колосья, зерновки красной, белой, черной и фиолетовой окраски, мучнистое и стекловидное зерно и т.д.
Открытый Н.И.Вавиловым закон справедлив не только для растений, но и для животных. Так, альбинизм встречается не только в разных группах млекопитающих, но и птиц, и других животных. Короткопалость наблюдается у человека, крупного рогатого скота, овец, собак, птиц, отсутствие перьев у птиц, чешуи у рыб, шерсти у млекопитающих и т.д.
Закон гомологических рядов наследственной изменчивости имеет огромное значение для селекционной практики. Он позволяет предугадать наличие форм, не обнаруженных у данного вида, но характерного для близкородственных видов, то есть закон указывает направление поисков. Причем искомая форма может быть обнаружена в дикой природе или получена путем искусственного мутагенеза. Например, в 1927 г. немецкий генетик Э.Баур, исходя из закона гомологических рядов, высказал предположение о возможном существовании безалкалоидной формы люпина, которую можно было бы использовать на корм животным. Однако такие формы не были известны. Было высказано предположение, что безалкалоидные мутанты менее устойчивы к вредителям, чем растения горького люпина, и большая их часть погибает еще до цветения.
Опираясь на эти предположения, Р.Зенгбуш начал поиски безалкалоидных мутантов. Он исследовал 2,5 млн. растений люпина и выявил среди них 5 растений с низким содержанием алкалоидов, которые явились родоначальниками кормового люпина.
Более поздние исследования показали действие закона гомологических рядов на уровне изменчивости морфологических, физиологических и биохимических признаков самых разных организмов — от бактерий до человека.
Искусственное получение мутаций
В природе постоянно идет спонтанный мутагенез. Однако спонтанные мутации — редкое явление. Например, у дрозофилы мутация белых глаз образуется с частотой 1:100000 гамет, у человека многие гены мутируют с частотой 1:200000 гамет.В 1925 г. Г.А.Надсон и Г.С.Филиппов открыли мутагенный эффект лучей радия на наследственную изменчивость у клеток дрожжей. Особое значение для развития искусственного мутагенеза имели работы Г.Меллера (1927), которые не только подтвердили мутагенный эффект лучей радия в опытах на дрозофилах, но и показали, что облучение увеличивает частоту мутаций в сотни раз. В 1928 г. Л.Стадлер использовал для получения мутаций рентгеновские лучи. Позже был доказан и мутагенный эффект химических веществ. Эти и другие эксперименты показали существование большого количества факторов, называемых мутагенными, способных вызывать мутации у различных организмов.
Все применяемые для получения мутаций мутагены делятся на две группы:
физические — радиация, высокая и низкая температура, механическое воздействие, ультразвук;
химические — различные органические и неорганические соединения: кофеин, иприт, соли тяжелых металлов, азотистая кислота и т.д.
Индуцированный мутагенез имеет большое значение. Он дает возможность создания ценного исходного материала для селекции, сотен высокопродуктивных сортов растений и пород животных, повышения в 10-20 раз продуктивности ряда продуцентов биологически активных веществ, а также раскрывает пути создания средств защиты человека от действия мутагенных факторов.
Модификационная изменчивость
Большую роль в формировании признаков организмов играет среда его обитания. Каждый организм развивается и обитает в определенной среде, испытывая на себе действие ее факторов, способных изменять морфологические и физиологические свойства организмов, т.е. их фенотип.Классическим примером изменчивости признаков под действием факторов внешней среды является разнолистность у стрелолиста: погруженные в воду листья имеют лентовидную форму, листья, плавающие на поверхности воды, — округлую, а находящиеся в воздушной среде, — стреловидные. Если же все растение оказывается полностью погруженным в воду, его листья только лентовидные. Некоторые виды саламандр темнеют на темном грунте и светлеют на светлом. Под действием ультрафиолетовых лучей у людей (если они не альбиносы) возникает загар в результате накопления в коже меланина, причем у разных людей интенсивность окраски кожи различна. Если же человек лишен действия ультрафиолетовых лучей, изменение окраски кожи у него не происходит.
Таким образом, изменения ряда признаков организмов вызывается действием факторов внешней среды. Причем эти изменения не наследуются. Так, если получить потомство от тритонов, выращенных на темном грунте, и поместить их на светлый, то все они будут иметь светлую окраску, а не темную, как их родители. Или, собрав семена со стрелолиста, выросшего в условиях полного погружения в воду, и высадив их в мелком водоеме, мы получим растения, листья которых будут иметь форму в зависимости от условий среды (лентовидные, округлые, стреловидные). То есть, данный вид изменчивости не затрагивает генотип и поэтому не передается потомкам.
Изменчивость организмов, возникающая под влиянием факторов внешней среды и не затрагивающая генотипа, называется модификационной.
Модификационная изменчивость носит групповой характер, то есть все особи одного вида, помещенные в одинаковые условия, приобретают сходные признаки. Например, если сосуд с эвгленами зелеными поместить в темноту, то все они утратят зеленую окраску, если же вновь выставить на свет — все опять станут зелеными.
Модификационная изменчивость является определенной, то есть всегда соответствует факторам, которые ее вызывают. Так, ультрафиолетовые лучи изменяют окраску кожи человека (так как усиливается синтез пигмента), но не изменяют пропорций тела, а усиленные физические нагрузки влияют на степень развития мышц, а не на цвет кожи.
Однако не следует забывать, что развитие любого признака определяется прежде всего генотипом. Вместе с тем, гены определяют возможность развития признака, а его появление и степень выраженности во много м определяется условиями среды. Так, зеленая окраска растений зависит не только от генов, контролирующих синтез хлорофилла, но и от наличия света. При отсутствии света хлорофилл не синтезируется.
Несмотря на то, что под влиянием условий внешней среды признаки могут изменяться, эта изменчивость не беспредельна. Даже в случае нормального развития признака степень его выраженности различна. Так, на поле пшеницы можно обнаружить растения с крупными колосьями (20 см и более) и очень мелкими (3-4 см). Это объясняется тем, что генотип определяет определенные границы, в пределах которых может происходить изменение признака. Степень варьирования признака, или пределы модификационной изменчивости, называют нормой реакции. Норма реакции выражается в совокупности фенотипов организмов, формирующихся на основе определенного генотипа под влиянием различных факторов среды. Как правило, количественные признаки (высота растений, урожайность, размер листьев, удойность коров, яйценоскость кур) имеют более широкую норму реакции, то есть могут изменяться в широких пределах, нежели качественные признаки (цвет шерсти, жирность молока, строение цветка, группа крови).
Знание нормы реакции имеет большое значение для практики сельского хозяйства
Таким образом, модификационная изменчивость характеризуется следующими основными свойствами:
ненаследуемость;
групповой характер изменений;
соответствие изменений действию фактора среды;
Статистические закономерности модификационной изменчивости
зависимость пределов изменчивости от генотипа.
Модификационная изменчивость многих признаков растений, животных и человека подчиняется общим закономерностям. Эти закономерности выявляются на основании анализа проявления признака у группы особей (n). Степень выраженности изучаемого признака у членов выборочной совокупности различна.
Каждое конкретное значение изучаемого признака называют вариантой и обозначают буквой v.
При изучении изменчивости признака в выборочной совокупности составляется вариационный ряд, в котором особи располагаются по возрастанию показателя изучаемого признака.
Частота встречаемости отдельных вариант обозначается буквой p.
Н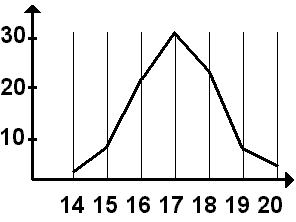
Рис. 338. Вариационная кривая.
а основании вариационного ряда строится вариационная кривая — графическое отображение частоты встречаемости каждой варианты (рис. 338).
Например, если взять 100 колосьев пшеницы (n) и подсчитать число колосков в колосе, то это количество будет от 14 до 20 — это численное значение вариант (v).
Вариационный ряд:
v = 14 15 16 17 18 19 20
Частота встречаемости каждой варианты
p= 2 7 22 32 24 8 5
Среднее значение признака встречается чаще, а вариации, значительно отличающиеся от него, — значительно реже. Это называется нормальным распределением. Кривая на графике бывает, как правило, симметричной. Вариации, как большие, чем средние, так и меньшие, встречаются одинаково часто.
Легко посчитать и среднее значение данного признака. Для этого используют формулу: (vּp)
М
 = n
= nгде М — средняя величина признака, в числителе сумма произведений вариант на их частоту встречаемости, в знаменателе — количество вариант. Для данного признака среднее значение равно 17,13.
Знание закономерностей модификационной изменчивости имеет большое практическое значение, поскольку позволяет предвидеть и заранее планировать степень выраженности многих признаков организмов в зависимости от условий внешней среды.