Биология и патология свиньи. Алексеева Ирина 4 курс 4 гр ФВМ очно-заочно, Особенности системы. Реферат Лист 29 Реферат По дисциплине Биология и патология свиньи Тема Особенности системы лактации у свиней студентка 4 курса 4 группы Факультета ветеринарной медицины
 Скачать 1.82 Mb. Скачать 1.82 Mb.
|

Реферат По дисциплине «Биология и патология свиньи» Тема: «Особенности системы лактации у свиней» Выполнила: студентка 4 курса 4 группы Факультета ветеринарной медицины очно-заочной формы обучения Алексеева И.А. Преподаватель: доктор биологических наук, профессор кафедры физиологии, фармакологии и токсикологии им. А.Н. Голикова и И.Е. Мозгова Максимов В.И. Москва 2021 г. ОглавлениеВведение 3 Биологические особенности свиней 7 Морфофизиология молочной железы свиней 13 Биологические особенности лактации у свиней 17 Регуляция молокообразования и молокоотдачи 21 Молочность, состав молозива и молока кормящих свиноматок 26 Заключение 29 ВведениеСвинья домашняя (Sus scrofa domesticus) — крупное парнокопытное, подвид кабана, одомашненная человеком около 7 тыс. лет назад. Длина тела составляет от 0,9 до 1,8 м, взрослая особь весит от 50 до 150 кг. По сравнению с другими парнокопытными, которые чаще бывают растительноядными, домашняя свинья всеядна, как и её предок, дикий кабан. Современная домашняя свинья является продуктом многовековой эволюции, результатом развития вида под воздействием естественного отбора и деятельности человека. Огромный полиморфизм (многообразие форм) этого вида свидетельствует о множестве исходных передовых форм, с одной стороны, и разной интенсивности и направлении изменения свиней под влиянием доместикации селекции – с другой. Благодаря работе селекционеров в мире насчитывается около 100 пород свиней. При всем разнообразии пород ведущее положение для производства товарной продукции сохраняют породы, используемые в большинстве стран: крупная белая или йоркшир, ландрас, дюрок, гемпшир и пьетрен (рис.1-5). Большинство пород, созданных в последние десятилетия, в том числе скороспелая мясная, получены в результате разных вариантов воспроизводительного скрещивания в первую очередь этих пород. Остальные породы имеют преимущественно региональное значение, но они хорошо приспособлены к местным условиям и еще долго будут сохранять свое значение, в том числе как генетический материал для создания новых пород.  В решении вопросов интенсификации свиноводства большая роль отводится совершенствованию и разработке новых технологий. В системе зоотехнических мероприятий, направленных на увеличение производства мяса и повышение рентабельности производства большое значение имеет дальнейшее совершенствование организации воспроизводства стада. Успешное развитие отрасли определяется рациональным использованием маточного поголовья, повышением его продуктивных качеств и выращиванием молодняка. Проблема выращивания и сохранения молодняка на ранних стадиях его роста и развития принадлежит к числу наиболее важных проблем зоотехнии и находится в связи с молочной продуктивностью свиноматок (рис.6). Уровень молочности свиней является определяющим фактором роста и развития поросят, достижения необходимой живой массы при раннем и сверхраннем отъеме. 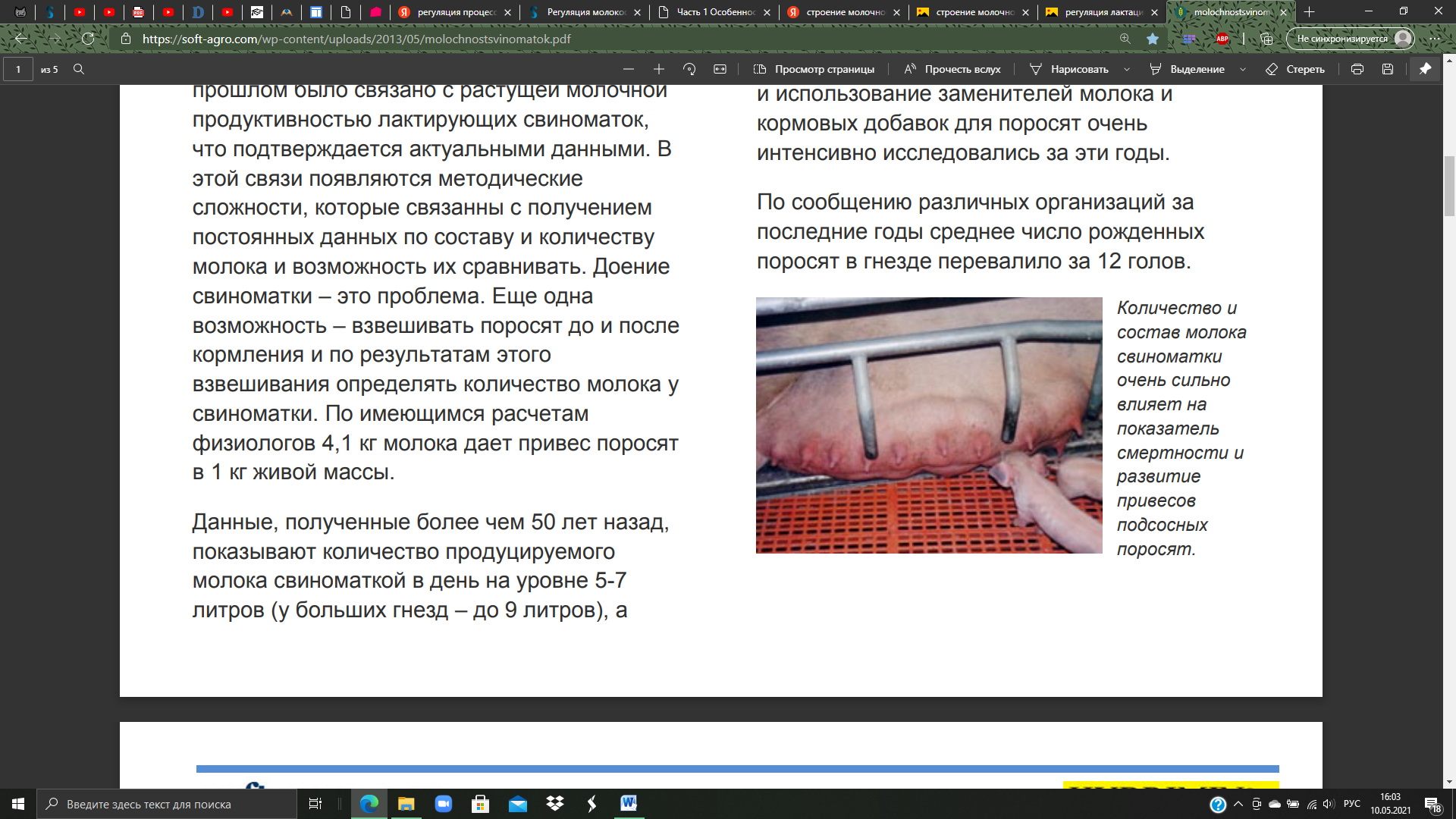 Рисунок 6. Количество и состав молока свиноматки очень сильно влияет на показатель смертности и развитие привесов подсосных поросят. Правильная организация воспроизводства стада в использовании маточного поголовья и выращивания поросят должна базироваться на знании биологических особенностей лактации животных. Несмотря на большие успехи, достигнутые в этой области, многие вопросы остаются неразрешенными. В частности, далеко не полно изучены особенности и закономерности лактации свиней, влияние молочной продуктивности на изменение роста поросят, динамика молочности в связи с хозяйственно-биологическими признаками (возрастом, живой массой, многоплодием), от показателей которых зависит формирование и использование маточного стада. Неразработанным разделом технологии промышленного свиноводства является режим кормления поросят в послеотьемный период, что служит причиной возникновения желудочно-кишечных заболеваний. Цель данной работы – изучить морфофизиологию множественного вымени свиньи домашней в период относительного физиологического покоя (поздний период супоросности) и интенсивного функционирования в период лактации, а также изучить биологические основы лактации свиней. Исходя из указанной цели, перед нами были поставлены следующие Задачи: изучить морфофизиологию множественного вымени свиньи домашней в период относительного покоя и интенсивного функционирования в период лактации; изучить закономерности и особенности лактации в связи с другими биологическими признаками и хозяйственно-полезнымп качествами; определить молочность, состав молозива и молока кормящих свиноматок. Биологические особенности свинейМолочные железы (glandulae lactiferae) – симметричные производные кожи, расположенные у свиней в области живота (рис.7). Они представляющие собой отдельные доли (lobi lactiferae), которые вместе образуют множественное вымя (ubera). 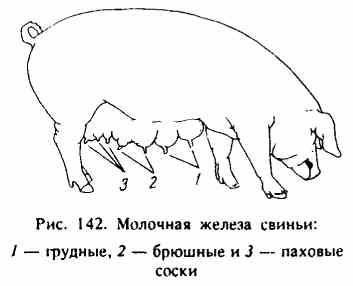 Рисунок 7. Молочная железа свиньи: 1-грудные, 2-брюшные, 3-паховые Молочные железы имеют сложное трубчато-альвеолярное строение. Молочная железа (mamma) состоит из тела (corpus mammae) и сосков (papillae). Каждая доля молочной железы имеет паренхиму и соединительнотканную строму. Паренхима железы состоит из долек (lobuli lactiferae). В образовании каждой принимают участие молочные альвеолы (alveolae lactiferae), молочные трубочки (tubulae lactiferae), молочные каналы (canales lactiferes) и молочные протоки (ductus lactiferi). Последние из указанных выше структур открываются в молочную цистерну (sinus lactiferi). В молочных альвеолах и трубочках осуществляется секреция молока, а во время усиленной лактации оно может секретироваться и в молочных каналах. Молочные каналы и молочные ходы служат для выведения молока в молочную цистерну. Развитие молочной железы у всеядных домашних млекопитающих начинается на ранних этапах эмбриогенеза. В ходе роста и развития молокообразующего аппарата и подготовки к секреции существенно изменяются структура и функция тканей множественного вымени. Морфогенез молочной железы – “маммогенез” – начинается в эмбриональный период с закладки молочных холмов. У эмбрионов по обеим сторонам вдоль белой линии живота они закладываются в виде узких длинных полосок утолщенного эпителия. Это так называемые млечные (молочные) тяжи. Млечные тяжи уменьшаются по длине, становятся прерывистыми и образуют серию расширенных и утолщенных эктодермальных млечных бугорков. Развивающиеся млечные бугорки погружаются в мезенхиму и переформатируются в млечные почки – первичную структуру будущего множественного вымени. Разрастаясь, мезенхима даёт начало будущей соединительной ткани и адипоцитам молочных холмов. Свинья домашняя характеризуется высоким многоплодием, коротким периодом пренатального развития, быстрым физиологическим и хозяйственным созреванием, высоким убойным выходом мясной продукции. В сравнении с другими видами сельскохозяйственных животных у свиноматок самый короткий срок вынашивания плодов (супоросности) - в среднем 114,6 дня. Это позволяет получать от них два, а при организации раннего отъема поросят 2,1-2,5 опоросов в год. Для свиней характерны интенсивные процессы пренатального развития органов и организма в целом. Масса плодов у них нарастает значительно быстрее, чем у крупного и мелкого рогатого скота. Уже во второй половине супоросности начинают определяться породные особенности плодов. У свиней молочная железа – множественное вымя. Оно располагается в области груди и живота, состоит из 5-8 парных молочных желез (молочных холмов) в виде отдельных долек с сосками. В отличие от вымени коров, овец и лошадей, у свиней нет выраженных молочных цистерн. У хороших свиноматок соски располагаются равномерно, а отдельные холмы вымени одинаково развиты. От молочных альвеол простирается сеть тончайших молочных проточков, которые по ходу многократно сливаются в более крупные. К вершине соска они оканчиваются двумя-тремя крупными в диаметре протоками (рис.8). На вершине каждого соска имеется от одного до четырех сосковых каналов с отверстиями для выхода молока. После опороса свиноматка кормит поросят не менее 25 раз в сутки, а в дальнейшем — 12-14 раз. Продолжительность лактации (периода с момента родов до прекращения молокоотдачи) зависит от породы, кормления и содержания животных, срока наступления новой беременности и т. д. У свиней она составляет два месяца после родов. В период эмбрионального развития свиней крупной белой породы «молочная полоска» закладывается на 20-22 день при длине зародыша в 13- 14 мм. Молочные соски становятся заметными на 28-30 день эмбрионального развития. При переходе от эмбрионального к постнатальному состоянию в гистоструктуре молочной железы свиней не наблюдается резких различий. До семимесячного возраста в постнатальном онтогенезе молочная железа развита слабо, в ней выявляются малоразвитые молочные ходы, на концах которых имеются альвеоло-трубки в состоянии формирования.  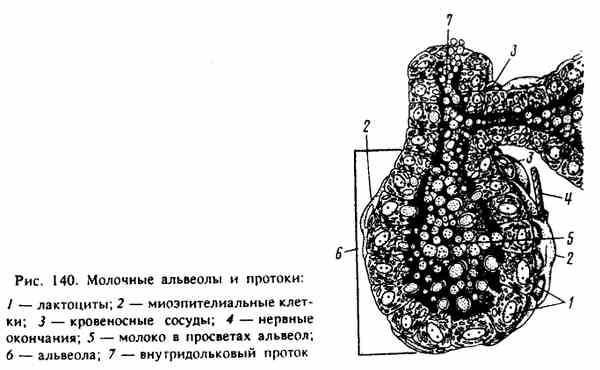 Рисунок 8. Молочные альвеолы и протоки К 12 месяцам молочная железа по уровню паренхиматозно-стромального соотношения и степени зрелости структур достигает морфологической, а к 18 – морфофункциональной зрелости. От центральных каналов молочной железы однодневных поросят отходят отростки, которые простираются довольно глубоко в соединительную ткань и в подкожную клетчатку, как сосковый канал, так и его ответвления имеют узенькие просветы. В дальнейшем в молочной железе 15-дневных поросят, увеличиваются размеры сосков и вся система разветвляющихся протоков молочной железы. Сосковые каналы сильнее разветвляются по сравнению с таковыми у однодневных поросят. От них отходят горизонтальные отростки, идущие параллельно поверхности кожи. Первичные отростки, в свою очередь, разветвляются на ещё более мелкие отростки у месячных поросят размеры протоков увеличены по сравнению с протоками у 15 дневных поросят. Центральные каналы и вся система ветвящихся протоков, обогащаются новыми ответвлениями. В месячном возрасте система разветвлений молочных протоков уже довольно сложная. Концы некоторых протоков чуть-чуть расширены в виде очень, мелких шариков, но эти расширения ещё не имеют никаких признаков просвета. По мере роста животного увеличиваются размеры имеющейся системы протоков молочной железы, их длина и разветвлённость. Увеличиваются немного и просветы протоков. Крупные выводные протоки покрыты многослойным эпителием. По мере уменьшения размеров протоков уменьшается и количество слоёв эпителиального покрова. Самые мелкие из них покрыты 2-слойным эпителием. Нарастает и количество ветвей, но альвеол с полостью не обнаруживается. Такое состояние молочной железы сохраняется у 2 и 4 месячных поросят. Увеличиваются размеры протоков и число разветвлений. В результате образуются не только вторичные, но и третичные, ответвления, которые в свою очередь, ветвятся так, что к 4 месяцам получается довольно сложная система протоков. У 2 и 4 месячных эмбрионов концы протоков несколько расширены в виде мелких шариков, но без просветов. Развитие молочной железы начинается на ранних этапах эмбриогенеза, причём в ходе роста и развития молокообразующего аппарата и подготовки к секреции существенно изменяются структура и функция тканей молочной железы. В литературе также накоплены данные о регуляторной роли иммунных реакций в лактогенезе и лактопоэзе. Важное значение в регуляции синтеза молока и молокоотдачи имеют клетки лейкоцитарного ряда. Достаточно подробно изучена роль сегментоядерных лейкоцитов в период лактации у человека и некоторых животных. Однако в литературе отсутствуют данные, достаточно полно характеризующие иммунный ответ и касающиеся функции моноцитов и макрофагов в период развития молочной железы и её функционирования. Моноцитарная активность важна для понимания восстановительных и иммунных процессов, протекающих в молочной железе в период лактации. Морфофизиология молочной железы свинейНаступление беременности у животных сопровождается в молочной железе интенсивными пролиферативными процессами, которые обеспечивают дальнейший рост её протоков и образование ампулообразных альвеолярных зачатков. Образование альвеолярных клеточных скоплений происходит в условиях перехода клеток от недифференцированного состояния к клеточной специализации. В конце беременности дифференциация клеток значительно преобладает над пролиферативными процессами. Сосуществование обоих процессов позволяет наблюдать многочисленные митозы, включая морфологические признаки наступающего секретообразования. Первичными эффектами пробуждения синтетической активности секреторные клетки обязаны индуцирующим влияниям мезенхимы. Исследование механизмов гормональной регуляции пролиферации и дифференциации выявило последовательность молекулярных процессов, приводящих к активации специфических генов, определяющих деятельность секреторных клеток. Инсулин, СТГ способны увеличивать скорость синтеза ДНК, что призвано, очевидно, стимулировать митотическую активность, благодаря которой наращивается клеточная масса развивающегося органа. При сочетании эстрадиола с инсулином обнаруживается увеличение митотической активности, связанное с изменением скорости перехода клетки из фазы G в фазу S клеточного цикла. Наряду с гормональными влияниями на развитие молочной железы вполне очевидна регулирующая роль эфферентной нервной системы. Денервация молочной железы значительно тормозит развитие структур альвеолярного отдела органа. Вместе с развитием структурных компонентов, характерных для дифференцированной секреторной клетки, образуется структура альвеолы. Вместе с дифференциацией секреторных клеток в альвеоле происходит становление функциональной активности миоэпителиоцитов. Характерные очертания миоэпителиальных клеток появляются с наступлением половой зрелости, однако формирование сократительного аппарата происходит во время беременности. Затем в начальный период лактации сократительная реакция миоэпителиоцитов претерпевает дальнейшее становление. Процессы прекращения лактационной функции осуществляются благодаря центральным регуляторным влияниям, о чем можно судить, рассматривая результаты гормональных и фармакологических воздействий. Пролактин, ОК, СТГ, АКТГ и резерпин, участвующий в освобождении некоторых гипофизарных гормонов, способны на некоторое время отодвинуть сроки инволюции органа. Применение эстрогенов ускоряет прекращение секретообразования. Необходимо учитывать, что в молочной железе процессы регенерации клеток могут проходить одновременно с процессом секретообразования, и в этом проявляется возможность смены клеточных поколений. В конце лактационного периода, с прекращением секреции, наступает период “старения” клеток, и они в большинстве подвергаются деструктивным процессам, отторгаются в просвет альвеолы, обнажая миоэпителий и базальную мембрану. В разрушении клеток принимают участие ферментативные процессы, связанные с образованием и функционированием специализированных органелл – лизосом. Мигрирующие в интерстициальную ткань лимфоциты, лейкоциты, макрофаги и моноциты выстраиваются рядом с разрушающимися альвеолами и довершают деструкцию клеточного материала. Наряду с разрушением клеток проходит дедифференциация ряда клеток с естественной потерей клеточной специализации. До наступления беременности дольчато-альвеолярный генез молочной железы выражен очень слабо и только во время беременности система протоков и дольчато-альвеолярная система достигают своего максимального развития. Наряду с изменениями клеточного состава в период, предшествующий лактации, становления структуры альвеолярного отдела молочной железы существенно изменяется цитоархитектоника ткани органа. Эстрогены стимулируют рост протоков, а прогестерон совместно с эстрогенами ответственен за рост и развитие альвеол. Важно отметить, что процессы развития миоэпителиальных клеток находятся под контролем эстрогенов, сопровождающих период развития молочной железы. При действии прогестерона развитие миоэпителиоцитов не наблюдается. Прогестерон оказывает тормозящее влияние и на деятельность зрелой миоэпительальной клетки в лактационный период, увеличивая латентный период молоковыделительной реакции альвеол на окситоцин и ацетилхолин. До второй половины беременности молочная железа заполнена главным образом жировыми клетками – липоцитами с оттесненными к периферии тонким слоем цитоплазмы и овально-удлиненными ядрами. Эти клетки формируют отдельные дольки, разделенные между собой и поддерживаемые тяжами стромы, сформированной из рыхлой соединительной ткани. В ней имеются транспортные кровеносные сосуды и элементы гемомикроциркуляторного русла. В последней трети беременности жировые клетки молочной железы постепенно замещаются соединительнотканной стромой. Этот процесс характеризовался значительным выведением триглицеридов из жировой клетки и использованием их в пластическом и энергетическом обеспечении развивающегося органа. Впоследствии замещаются все жировые клетки, на месте которых начиналась интенсивная пролиферация недифференцированных клеток галактофорных каналов с мелкими темноокрашенными ядрами, которые прилегали друг к другу плотными скоплениями. К моменту начала лактации боковые поверхности плазматической мембраны формируют структуры замыкательного комплекса. Основой его является плотный контакт, обусловленный истинным слиянием наружных слоев и соседних мембран в апикальной зоне эпителиальных клеток. Биологические особенности лактации у свинейПоказано, что передние соски снабжаются кровью обильнее, чем задние. Этим подтверждается и общеизвестное наблюдение свиноводов, что передние соски вымени богаче молоком, чем задние. Снабжение кровью передних молочных желез осуществляется через межреберные и наружную грудную артерии. Задние железы питаются в основном через наружную половую (срамную) артерию. Передняя группа молочных желез свиньи (с 1-й по 5-ю) питается главным образом ветвями надчревной артерии, являющейся продолжением внутренней грудной артерии. В кровоснабжении этой группы желез некоторое участие принимают также межреберные артерии. В кровоснабжении других желез, начиная с 6-7-й, участвуют ветви наружной половой артерии. К 5-й железе подходят последняя межреберная артерия и концевые ветви наружной половой артерии. Задняя группа молочных желез питается ветвями наружной половой и частично поясничных артерий. У домашних свиней половое созревание наступает значительно раньше, чем у диких (самки становятся половозрелыми в 7-8 месяцев). Интенсивность роста молодняка у них также более высокая. Однако продолжительность лактации у домашних свиней по сравнению с дикими изменилась весьма незначительно. Подсосный период у самок крупной белой породы продолжается в среднем 93 дня (и дикие свиньи лактируют около 3 месяцев). Однако в хозяйствах отъем поросят от матки обычно практикуют в 2-месячном возрасте, а в ряде стран при специальных видах подкормки и еще раньше. В зоотехнической практике молочность свиноматок обычно определяют по изменению веса поросят. Было показано, что свиньи за лактацию дают довольно много молока - в среднем 3-4 л в сутки, а иногда и до 8-9 л. Молочная продуктивность их зависит от породы, возраста, индивидуальных особенностей, характера кормления, числа поросят в помете, сезона опороса и других факторов. Повышенная молочная продуктивность обычно свойственна свиньям высокопродуктивных многоплодных пород (крупная белая, миргородская, украинская степная). В процессе лактации суточные удои свиноматок закономерно изменяются. Высшая точка лактации у большинства животных наблюдается между второй и четвертой неделями лактации, или во второй декаде подсосного периода. В целом в первый месяц лактации свиньи дают больше молока, чем во второй. В третьем месяце лактации количество выделяемого молока резко падает. Активная секреция в молочной железе свиньи начинается за 1-2 дня до опороса. В первые дни после опороса она настолько интенсивна, что во время кормления поросят молоко самопроизвольно струйками вытекает из незанятых сосков. На этой стадии лактации достаточно лишь легкого прикосновения поросенка к соску, чтобы началось молоковыделение, сопровождающееся выпрыскиванием молока наружу. Это явление особенно выражено у обильномолочных маток. С течением лактации быстрота и сила этой ответной реакции на прикосновение к соскам заметно уменьшается, снижается и молокопродукция. Если в начале лактации рефлекс молоковыделения, возникающий от прикосновения поросенка к одному соску, распространяется на другие соски, то позднее выделение молока происходит только из сосков, непосредственно раздражаемых поросятами, да и то далеко не сразу. В этом и состоит своеобразие выделения молока у свиней. Уже через неделю после рождения поросята, прежде чем приступить к сосанию, длительное время массажируют молочные железы (рис.9). Предварительный массаж молочных желез совершенно необходим для реализации молоковыделительного рефлекса у свиньи в этот период.  Рисунок 9. Массаж молочной железы Во время этого массажа поросята, захватив сосок ртом, начинают энергично разминать молочную железу, что продолжается от 2 до 5, а иногда до 10 мин и более. При отсутствии систематического раздражения молочной железы сосанием и выведения секрета молочная железа очень быстро подвергается инволюции. Следовательно, массаж вымени у свиней имеет очень важное значение для нормального осуществления мо- локовыведения. Свиноматки подпускают поросят к вымени в начале лактации всего лишь на 3-4 мин. В начале лактации длительность выделения молока из сосков составляет 1-2 мин, а к концу лактации этот период измеряется секундами. При этом поросята сначала энергично массажируют вымя, после чего затихают и начинается акт сосания. Он длится, как считают многие ученые, 40-50 сек. По другим данным, выделение молока у свиноматок продолжается 13-18 сек и реже 30-32 сек. В течение суток поросята сосут свинью в среднем 10-14 раз и иногда до 17 и даже 28 раз. Свиноматка кормит поросят круглосуточно с более или менее равномерными интервалами. Чем больше интервалы между кормлениями, тем ниже молочная продуктивность свиней. Поэтому, увеличивая частоту кормления поросят, можно таким путем повысить молочность свиноматок. При часовых интервалах продуктивность свиноматок достигает в 8,87 кг молока, при двухчасовых только 6,1 кг. Интервалы между кормлениями обычно увеличиваются по мере развития лактации. В начале ее свиньи кормят поросят чаще (через 55 мин), чем в конце (через 90 мин и более). Новорожденный получает за одно активное сосание 25-45 г молозива. Общее количество молозива, выделяемое маткой за одно сосание, колеблется от 140 до 545 г в первые и от 80 до 405 г во вторые сутки. На одного поросенка в начале лактации (первая пятидневка) приходится в среднем 1375 г молока в сутки, а в конце лактации (12-я пятидневка) - 1254 г. Максимальное количество молока на поросенка приходится с четвертой по восьмую пятидневку лактации (2457-2608 г). Между числом поросят в помете и общим их весом существует зависимость. Однако вес приплода не всегда правильный показатель наследственной молочности. Регуляция молокообразования и молокоотдачиРегуляция молокообразования осуществляется в результате взаимодействия нервной, эндокринной и сосудистой систем (рис.10).  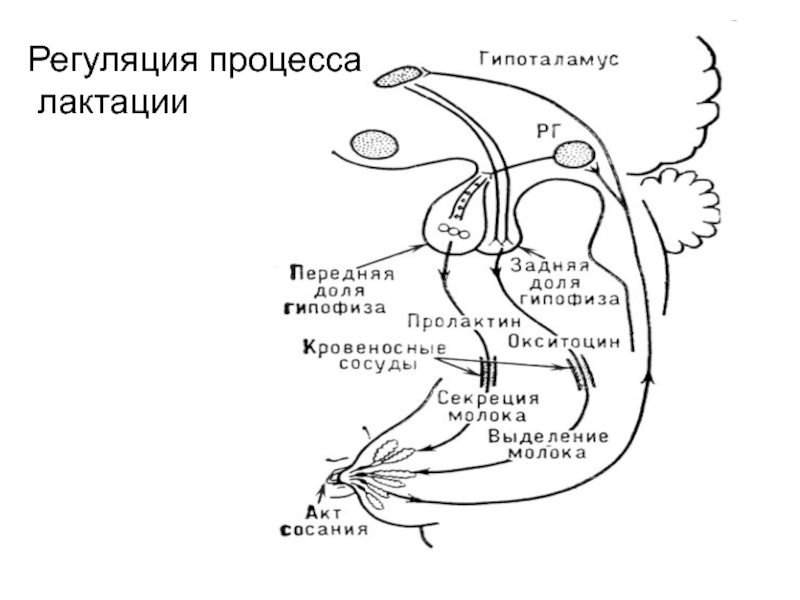 Рисунок 10. Схема регуляции лактации Нервная регуляция. Сложную функцию молокообразования молочная железа выполняет благодаря совершенству рефлекторной регуляции. Процесс молокообразования осуществляется при участии коры полушарий мозга и ряда отделов центральной нервной системы, строго согласованных в своей деятельности и образующих единую морфофункциональную структуру, которую можно назвать центром, регулирующим секрецию и выведение молока, или лактационным центром (И.И. Грачев, 1970). Этот центр обусловливает подготовку молочной железы к лактации, пуск в ход секреторного процесса, выведение молока. Он находится во взаимодействии с пищевым, дыхательным, сосудодвигательным, половым и другими центрами. Каждый из отделов центра лактации выполняет свою функцию: спинной мозг обеспечивает «грубую» регуляцию двигательной функции молочной железы; продолговатый мозг регулирует кровоснабжение различных её частей; гипоталамо-гипофизарная система – обеспечивает продукцию рилизинг-факторов (либеринов) и ингибирующих факторов (статинов) определяющих продукцию лактогенных гормонов. Супраоптическое ядро гипоталамуса секретирует преимущественно вазопрессин, а паравентрикулярное – вазопрессин и в большей степени окситоцин. Аденогипофиз выделяет пролактин. В гипоталамусе осуществляется координация деятельности молочной железы с другими системами организма; кора полушарий мозга. От гипоталамуса возбуждение идёт к коре больших полушарий, где и формируется общность нервных центров, осуществляющих контроль за интенсивностью лактационного процесса и создается особое состояние организма «лактационная доминанта». Рефлекс молокоотдачи. От молочной железы (при доении, сосании) по центростремительным нервам афферентные импульсы передаются в афферентные нейроны спинномозговых ганглиев и дорсальные столбы серого вещества спинного мозга. Отсюда через вставочные нейроны серого вещества сигнал поступает на эфферентные нейроны вентральных столбов серого вещества и по двигательным аксонам передается в молочную железу. Это обеспечивает снижение тонуса гладкой мускулатуры молочной железы и раскрытие сфинктеров протоков и сосков. В результате наступает припуск и легко удаляется цистернальная порция молока, составляющая 15-17% удоя. Центральная часть дуги рефлекса находится в коре полушарий мозга в супраоптических и паравентрикулярных ядрах гипоталамуса. Эфферентные пути от гипоталамуса идут к задней доле гипофиза в специальном пучке, насчитывающем до 1000 аксонов, который называют супраоптико-гипофизарным трактом. Сюда поступают импульсы из вымени. Под их влиянием синтезируется окситоцин в нейронах гипоталамуса и накапливается в задней доле гипофиза. Отсюда гормон поступает в воротную систему гипофиза и в кровь и по кровеносной системе в молочную железу. Альвеолярное молоко выводится из молочной железы только при участии гормонов гипоталамо-гипофизарной системы. Она возбуждается поступающими к ней импульсами от рецепторов молочной железы. Выделение гормона окситоцина и поступление его с кровью в молочную железу происходит не сразу, латентный период 30-40 с и более, который может варьировать у разных животных. Окситоцин оказывает стимулирующее влияние на миоэпителий альвеол, который начинает быстро сокращаться и сжимать снаружи альвеолы, в результате секрет выводится в протоки альвеол. В рефлексе молокоотдачи различают две фазы: нервная, начинается через 2-6 с после раздражения рецепторов, характеризуется выделением молока из крупных протоков вследствие раздражения рецепторов молочной железы и передачи импульсов в центры и обратно по короткой рефлекторной дуге; нейро-гуморальная, характеризуется поступлением окситоцина в кровь, а затем в молочную железу и выведением молока из альвеол и мелких протоков вследствие сокращения миоэпителия под воздействием нейро-гуморального механизма. Основное звено в этом механизме – влияние окситоцина на миоэпителий альвеолярного аппарата (на звёздчатые клетки). На регуляторные механизмы лактации влияет характер системы содержания свиноматок. Гуморальная регуляция В срединном возвышении гипоталамуса образуется пролактин-рилизинг-фактор (PRF) вызывающий секрецию пролактина в аденогипофизе. Пролактин непосредственно воздействует на эпителиальные клетки молочной железы, стимулирует синтез белков, в том числе казеина, лактоальбумина, а также липидов и углеводов молока. Пролактин постоянно присутствует в крови лактирующих животных, но уровень содержания гормона непостоянен. Во время доения/сосания его содержание резко увеличивается, особенно в начальный период, затем происходит постепенное уменьшение. Концентрация пролактина в крови повышается уже в момент преддоильной подготовки (условнорефлекторная фаза), в конце доения его уровень снижается почти до нулевых значений. Снижение уровня секреции пролактина обусловлены действием на аденогипофиз, образующегося в преоптической области гипоталамуса пролактин-ингибирующего фактора (PIF). Тормозящее действие на гипоталамус, а следовательно, на синтез и выделение пролактина из гипофиза оказывают катехоламины, эстрогены. Стимулом для выделения гормонов гипофиза является снижение их уровня из циркулирующей крови в результате расхода данных гормонов на синтез и выведение молока. В этом заключается принцип обратной связи между центром лактации и молочной железой. Гормоны щитовидной железы усиливают лактацию. Тиреоидные гормоны стимулируют морфогенез молочной железы, способствуют наступлению лактации и повышают секреторную способность клеток молочной железы. Тироксин усиливает активность ферментных систем переноса электронов и фосфорилирование в митохондриях. Надпочечники также принимают участие в регуляции секреции молока. Кортизол непосредственно воздействует на секреторный эпителий (ускорение транскрипции мРНК). Удаление надпочечников ведёт к прекращению секреции. Гормон желтого тела прогестерон оказывает тормозящее влияние на лактацию. После родов и удаления из родовых путей плаценты содержание эстрогенов и прогестерона в крови резко падает, а их тормозящее влияние на гипофиз прекращается, поэтому последний начинает продуцировать значительное количество пролактина. Секреция молока (вначале молозива) начинается после родов и продолжается на протяжении всего периода лактации. Гормон поджелудочной железы инсулин способствует депонированию углеводов и жиров, что обеспечивает надежное снабжение молочной железы метаболитами в промежутках между приёмами корма, а также активирует перенос сахаров и аминокислот через клеточные мембраны, что сказывается на скорости и характере секреторных процессов в молочной железе. Роль околощитовидных желёз в обеспечении активно секретирующей молочной железы кальцием ярко проявляется при их удалении – наступает резкое угнетение лактации. Торможение молокоотдачи осуществляется через эфферентные нервы молочной железы и надпочечники вследствие рефлекторного выделения адреналина. Молочность, состав молозива и молока кормящих свиноматокДля оценки молочности маток определяют вес гнезда поросят в месячном возрасте. При обычной подкормке поросят в племенных стадах он должен составлять не менее 70 кг, в пользовательных – 50 – 60 кг. Учитывают также и материнские качества свиноматок, способность их хорошо выкармливать сосунов. Молоко свиней существенно отличается по своему составу от молока других сельскохозяйственных животных. Оно представляет собой белую, клейкую, сладковатую на вкус жидкость с характерным запахом и может долго сохраняться в свежем виде. Сухого вещества в молоке свиней содержится от 16 до 20% (в среднем 18%). Содержание жира в молоке свиней очень велико. Его концентрация колеблется от 3 до 16% и зависит от возраста животных, числа опоросов, месяца лактации, расположения сосков и других факторов. Белка в молоке свиней содержится от 4,9% до 7,6%. В его состав входит 57-58% казеина, 7-8% альбуминов, 10% глобулинов, 17-18% протеозо-пептона и 7-8% небелкового азота. Интересно, что между белками молока и сывороткой крови свиней имеется определенная взаимосвязь. Содержание лактозы в молоке колеблется от 3,6 до 6,64%. Минеральный состав молока свиней изучен еще недостаточно. Однако известно, что на 100 г сухого вещества в молоке свиней приходится 0,068 г фосфора, 0,26 г кальция, 0,202 г хлоридов и 1,39 г золы. Молоко свиней очень богато витаминами. В нем обнаружен витамин А, тиамин, рибофлавин, пантотеновая, никотиновая и аскорбиновая кислоты. Удельный вес молока равен в среднем 1,032. В процессе лактации происходят существенные изменения ряда составных частей молока. Из органических компонентов молока наибольшим изменениям в процессе лактации подвержено содержание жира. Жирность молока к концу лактации обычно увеличивается. Содержание белка убывает ко второй пятидневке и затем с восьмой пятидневки вновь увеличивается (до 6,4%); хотя и не достигает первоначального уровня. Закономерных изменений концентрации лактозы в молоке в зависимости от стадии лактации не обнаружено. Динамика концентрации минеральных веществ в ходе лактации изменяется: содержание кальция увеличивается к концу лактации с 0,9 до 1,56 г/кг, а фосфора - с 1,1 до 1,7 r/кг. Содержание в молоке свиноматок калия и натрия уменьшается к концу лактации: калия - с 1,06 до 0,425 г/кг, натрия - с 0,794 до 0,450 г/кг. Общее содержание золы по ходу лактации имеет тенденцию к увеличению. Установлено, что на содержание сухого вещества, золы, кальция и фосфора в молоке существенное влияние оказывает комплекс микроэлементов - железа, цинка, меди и кобальта. Существенные различия наблюдаются в составе молока, полученного из разных сосков одного и того же животного. Особенно отличается молоко передней и задней пар сосков. В передних сосках молоко несколько беднее сухим веществом, жиром. Свиноматки-первоопороски имеют несколько большую жирность молока, чем свиньи старшего возраста. В отличие от других видов сельскохозяйственных животных у свиноматок в порциях молока разового удоя жир распределен относительно равномерно. Свойства молока зависят и от сезона лактации. Летнее молоко обычно более питательное. Как и у других видов животных, молозиво свиней заметно отличается по своему составу от зрелого молока. В молочных железах свиноматок секреторная активность наступает уже в середине супоросного периода, однако выделение молозива начинается лишь за несколько часов до опороса или спустя 5-10 ч после него. Сразу же после опороса молозиво имеет светло-коричневый цвет, по консистенции напоминает сметану, вкус его приторный, сладковатый. На второй день молозиво имеет уже желтокоричневый цвет, оно более тягучее, клейкое и слаще молозива первого дня. Секрет молочных желез свиней в начале лактации характеризуется высоким содержанием протеина и низким содержанием жира и лактозы. Содержание жира в молозиве в начале увеличивается с 7,2 до 12%, а уже на 4-6-й день снова снижается до 8,8%. В целом содержание жира в молозиве колеблется от 6,43 до 9,20%. Содержание белка в молозиве у свиней колеблется от 13,27 до 14,43%, в зависимости от опороса и сезона года. Его концентрация уже через 24 ч резко уменьшается. Лактозы в молозиве свиней в первый день содержится 3,22%, на второй день - 4,49, на третий - 4,81, на четвертый - 5,46 и на пятый - 5,96% . Количество сухого вещества на второй день уменьшается с 24,31 до 17,8%, а затем вновь постепенно увеличивается и на пятый день составляет 19,52%. Содержание золы уменьшается до четвертого дня (с 1,96 до 0,85%). Уменьшается и содержание витамина С (с 29,59 мг/100 мл в первый день до 14,33 мг/100 мл на пятый день). Калорийность молозива свиней выше калорийности молока. В первый день она составляет 155 к калорийности молока, во второй день - 106,59, в третий - 113,02, в четвертый - 110,63 и в пятый - 120%. ЗаключениеЛактация свиней — важнейшая деятельность животного организма, посредством которой осуществляется вскармливание поросят молоком и обеспечение развивающегося приплода полноценным питанием. Благодаря этому организм новорожденного животного на самых ранних стадиях своего развития, т. е. в наиболее критический период жизни, находится в зависимости от секреторной деятельности молочной железы и выведения молока во время сосания. Интенсивный рост, сохранение и последующее развитие молодняка обусловлены молочной продуктивностью свиноматок. По опубликованным в 1971 году результатам, свиноматка в среднем производила во время лактации 8 кг молока (6-11) кг в день, а период отъема уже был сокращен до 4-6 недель. Во время одного кормления поросята потребляли около 30 г молока на голову; в день – 700- 1100 г. За лактацию свиноматка при правильном кормлении производила от 300 до 450 кг молока. С тех пор интерес к повышению продуктивности свиноматок очень сильно возрос. Переход на ранний отъем, разработка метода выращивания поросят без свиноматки и использование заменителей молока и кормовых добавок для поросят очень интенсивно исследовались за эти годы Увеличение многоплодности и уровня привесов в свиноводстве повышает требования к молочности лактирующих свиноматок. Чтобы иметь возможность оценить количество и течение лактации во время подсосного периода, подходит оценка привесов подсосных поросят в гнезде, определяемых регулярными взвешиваниями. Задача племенной работы – развитие не только параметров размера гнезда и здоровья рожденных поросят, но и функциональных признаков. У свиноматок это материнский инстинкт, молочная продуктивность и охотность поедания корма (потребление корма) во время периода лактации. Список литературы Бажов, Г. М. Свиноводство / Г. М. Бажов, В. А. Погодаев. Ставрополь: Сервисшкола, 2009. 528 с. Бекенев, В. А. Технология разведения и содержания свиней / В. А. Бекенев, - СПб. Издательство «Лань». – 2012. – 416 с. Гончарова, В. М. Морфологические основы лактогенеза у свиней крупной белой породы в первую половину супоросности / В. М. Гончарова // Актуальные проблемы ветеринарии и животноводства: ГНУ СамНИВС Россельхозакадемии. – Самара, 2010. Гончарова, В. М. Особенности гистофизиологии молочной железы свиней в периоды репродуктивного цикла: автореф. дис. … канд. биол. наук / Гончарова В. М. – Оренбург, 2011. Гончарова, В. М., Абрамова, Л. Л. Морфология молочной железы свиней в период эструса во время полового созревания / В. М. Гончарова, Л. Л. Абрамова // Известия Оренбургского государственного аграрного университета, Оренбург, 2008. Т. 4, № 20-1. Зеленевский, Н. В., Зеленевский, К. Н. Анатомия животных – СПб, Лань, 2014, 844 с. Исламов, Р. Р. Особенности белкового состава молозива и молока у крупного рогатого скота и свиньи / Исламов Р. Р., Хаертдинов Р. Р. // Сельскохозяйственная биология. Серия: Биология растений. Серия: Биология животных. – 2009. - № 6. Понд, У. Д. Биология свиньи: Пер. с англ. и предисл. В. В. Попова / У. Д. Понд, К. А. Хаунт. – М. : Колос, 1983. – 334 с. Рахматов, Л. А. Молочная продуктивность свиноматок /Л.А. Рахматов // Ученые записки КГАВМ. – Казань, 2010. – Т. 204. |
