ххреф. Реферат Строение и биологическая роль холестерина. Гиперхолестеринемия и атеросклероз (обзор литературы)
 Скачать 0.65 Mb. Скачать 0.65 Mb.
|

|
Федеральное государственное бюджетное образовательное учреждение высшего образования «Новосибирский государственный медицинский университет» Министерства здравоохранения Российской Федерации (ФГБОУ ВО НГМУ Минздрава России) Медико-профилактический факультет Кафедра медицинской химии Реферат Строение и биологическая роль холестерина. Гиперхолестеринемия и атеросклероз (обзор литературы) Выполнила: студентка 1 курса лечебного факультета специальности «Лечебное дело», 7 группы Сатторова Наргиз Нуриддиновна Проверила: преподаватель Дегтярева А.О Новосибирск – 2018 ОГЛАВЛЕНИЕ 1. Биомедицинское значение...3 2. Химическое строение...3 3. Кинетика оксигенирования гемоглобина...7 4. Конформационные изменения в окружении гемогруппы...9 5. Транспорт двуокиси углерода...10 6. Молекулярная основа эффекта Бора...12 7. Концентрация гемоглобина...13 8. Способы исследования...14 9. Метгемоглобин...15 10. Сульфогемоглобин...15 11. Типы гемоглобина...16 12. Методы дифференцировки видов гемоглобина...19 13. Гемоглобин при серповидноклеточной анемии...22 14. Талассемии.... 25 15. Список литературы...26 СТРОЕНИЕ И ФУНКЦИИ ГЕМОГЛОБИНА БИОМЕДИЦИНСКОЕ ЗНАЧЕНИЕ Гемсодержащие белки участвуют в процессах связывания и транспорта кислорода, в транспорте электронов в фотосинтезе. Детальное изучение гемоглобина выявляет ряд структурных аспектов, общих для многих белков. Говоря о большом биомедицинском значении этих белков, мы имеем в виду, что результаты, полученные при исследовании, наглядно иллюстрируют структурно-функциональные взаимосвязи. Кроме того, эти исследования выявляют молекулярную основу ряда генетических болезней, таких как серповидноклеточная анемия (возникающая в результате изменения свойств поверхности b-субъединицы гемоглобина) или талассемия (хроническое наследуемое гемолитическое заболевание, характеризующееся нарушениями процессов синтеза гемоглобина). Летальный эффект цианида и окиси углерода объясняется тем, что эти вещества блокируют физиологическую функцию гемопротеинов - цитохромоксидазы и гемоглобина соответственно. Наконец, стабилизация четвертичной структуры дезоксигемоглобина 2,3-бифосфоглицератом (ДФГ) занимает центральное место в исследовании механизмов кислородной недостаточности в условиях высокогорья и процессов адаптации к этим условиям. 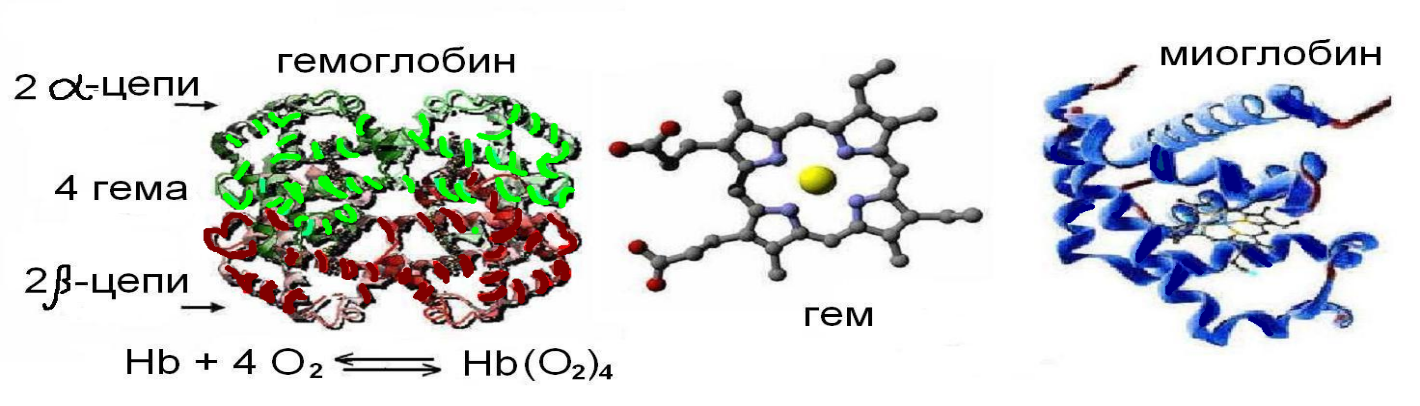 Рис.1 Структура гемоглобина ХИМИЧЕСКОЕ СТРОЕНИЕ Химически гемоглобин относится к группе хромопротеидов. Его простетическая группа представляет собой ферросоединение протопорфирина IХ, с молекулярным составом С34Н32О4N4Fe и носит название гем.Она придает соединению окраску. Белковый компонент гемоглобина называется глобином. Гемоглобиновая молекула содержит 4 гема и 1 глобин.(рис.1) Аминокислоты расположены в глобине в виде четырех полипептидных цепочек; две из них идентичны по структуре, и их обозначают как альфа-цепочки; две другие тоже идентичны между собой и их обозначают как бета-цепочки. Следовательно, формулу глобина можно выразить как альфа-альфа/бета-бета или альфа2бета2. альфа-полипептидная цепь состоит из 141, бета-полипептидная цепь - из 146 аминокислот.рис.2 Аминокислотный состав и последовательность (секвенция) альфа- и бета-цепей: альфа-полипептидная цепь заканчивается комбинацией аминокислот валина-лейцина, а бета-полипептидная цепь - комбинацией валина-гистидина-лейцина. альфа- и бета-полипептидные цепи в гемоглобиновой молекуле не расположены линейно, как это выглядит на первый взгляд из данных ("первичная структура") на рис.1 По причине существования интрамолекулярных сил, полипептидные цепи скручиваются в форме типичной для белков альфа-геликсовой спирали ("вторичная структура"). Сама альфа-геликсовая спираль на каждую альфа- и бета-полипептидную цепь огибается пространственно, образуя сплетения овоидной формы ("третичная структура"). На рис. 2 показано "третичное сгибание" полипептидных геликсовых спиралей в пространстве. Отдельные части альфа-геликсовых спиралей полипептидных цепей отмечают латинскими буквами от А до Н (рис. 2 и рис. 3) 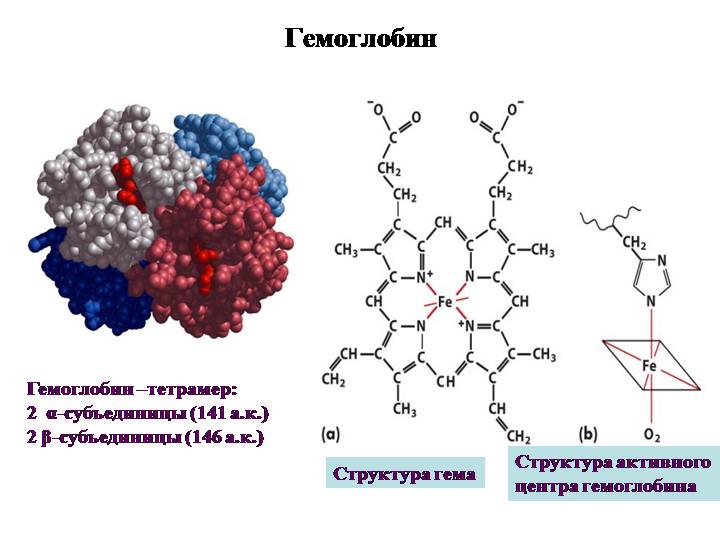 Рис.2 Все четыре третично изогнутые альфа- и бета-полипептидные цепи располагаются пространственно в определенном соотношении ("кватернерная структура"), что показано схематически на рис. 4. Они связаны между собой не настоящими химическими связями, а межмолекулярными силами. Четыре гема гемоглобиновой молекулы расположены в форме дисков меду складками четырех альфа-, соответственно бета-полипептидных цепей (рис. 3), причем каждый гем связан с одной полипептидной цепью посредством координационной связи между Fe++-атомом гема и гистидиновым остатком полипептидной цепи (рис. 5). Комплекс, составленный из одного гема и одной альфа-, респ. бета-полипептидной цепи, называется Сведберговой единицей. Очевидно гемоглобиновая молекула состоит из четырех Сведберговых единиц. В настоящее время принято считать, что молекулярный вес гемоглобина равен 64458, т.е. на один атом железа, соответственно приблизительно на Сведбергову единицу полагается по 16115. Кроме координационной связи, существующей между полипептидными цепями глобина, Fe++ атом гема располагает еще тремя координационными связями (рис. 5) Две из них связаны двумя азотными атомами порфиринового кольца, а третья, в среде с низким парциальным давлением кислорода (венозная кровь), связана с одной молекулой воды ( редуцированный гемоглобин ). В среде с высоким парциальным давлением кислорода (артериальная кровь), третья координационная связь соединена с одной молекулой кислорода, причем получается соединение - оксигемоглобин. Путем непрерывного превращения оксигемоглобина в редуцированный гемоглобин и обратно, осуществляется перенос кислорода из легких к тканям. [2] КИНЕТИКА ОКСИГЕНИРОВАНИЯ ГЕМОГЛОБИНА Гемоглобин связывает четыре молекулы кислорода на тетрамер (по одной на гем в каждой субъединице); особенно важным отличаем его от миоглобина является кривая насыщения кислородом, которая имеет сигмоидную форму (рис. 6). Таким образом, способность гемоглобина связывать кислород зависит от того, содержатся ли в данном тетрамере другие молекулы кислорода. Если да, то последующие молекулы кислорода присоединяются легче. Следовательно, для гемоглобина характерна кинетика кооперативного связывания, благодаря которой он связывает максимальное количество кислорода в легких и отдает максимальное количество кислорода при тех парциальных давлениях кислорода, которые имеют место в периферических тканях. Сродство гемоглобинов к кислороду характеризуется величиной Р50 - значением парциального давления кислорода, при котором наблюдается полунасыщение гемоглобина кислородом. Значение Р50 у разных организмов существенно различается, но во всех случаях оно превышает значение парциального давления кислорода в периферических тканях рассматриваемого организма. Это хорошо иллюстрирует фетальный гемоглобин человека (НВF). Для HbA Р50=26 мм. рт. ст., а для HbF Р50=20 мм. рт. ст. Благодаря этой разнице гемоглобин F отбирает кислород у HbA, находящегося в плацентарной крови. Однако после рождения ребенка HbF утрачивает свою функцию; обладая более высоким сродством к кислороду, он высвобождает меньшее его количество в тканях. ОКСИГЕНИРОВАНИЕ СОПРОВОЖДАЕТСЯ ЗНАЧИТЕЛЬНЫМИ КОНФОРМАЦИОННЫМИ ИЗМЕНЕНИЯМИ ВГЕМОГЛОБИНЕ Связывание кислорода сопровождается разрывом солевых связей, образованных концевыми карбоксильными группами субъединиц (рис.7) Это облегчает связывание следующих молекул кислорода, поскольку при этом требуется разрыв меньшего числа солевых связей. Указанные изменения заметно влияют на вторичную, третичную и особенно четвертичную структуру гемоглобина. При этом одна А/В-пара субъединиц поворачивается относительно другой А/В-пары, что приводит к компактизации тетрамера и повышению сродства гемов к кислороду (рис. 8 и 9). КОНФОРМАЦИОННЫЕ ИЗМЕНЕНИЯ В ОКРУЖЕНИЕ ГЕМОГРУППЫ Оксигенирование гемоглобина сопровождается структурными изменениями в окружении гемогруппы. При оксигенировании атом железа, который в дезоксигемоглобине выступал на 0,06 нм из плоскости гемового кольца, втягивает в эту плоскость (рис. 10). Вслед за атомом железа ближе к гему перемещается проксимальный гистидин (F8), а также связанные с ним соседние остатки. ТРАНСПОРТ ДВУОКИСИ УГЛЕРОДА Гемоглобин не только переносит кислород от легких к периферическим тканям, но и ускоряет транспорт углекислого газа от тканей к легким. Гемоглобин связывает углекислый газ сразу после высвобождения кислорода; примерно 15% углекислого газа, присутствующего в крови, переносится молекулами гемоглобина. Находящаяся в эритроцитах карбоангидраза катализирует превращение поступающего из тканей углекислого газа в угольную кислоту (рис.11). Угольная кислота быстро диссоциирует на бикарбонат-ион и протон, причем равновесие вдвинуто в сторону диссоциации. Для предотвращения опасного повышения кислотности крови должна существовать буферная система, способная поглощать избыток протонов. Гемоглобин связывает два протона на каждые четыре освободившиеся молекулы кислорода и определяет буферную емкость крови (рис. 12). В легких идет обратный процесс: присоединение кислорода к дезоксигемоглобину сопровождается высвобождением протонов, которые связываются с бикарбонат-ионами, переводя их в угольную кислоту. Далее эффективно действующая карбоангидраза катализирует превращение угольной кислоты в углекислый газ, выдыхаемый из легких. Таким образом, связывание кислорода тесно сопряжено с выдыханием углекислого газа. Это обратимое явление известно как эффект Бора. Эффект Бора является свойством тетрамерного гемоглобина и определяется гем-гемовым взаимодействием, лежащим в основе кооперативных эффектов. МОЛЕКУЛЯРНАЯ ОСНОВА ЭФФЕКТА БОРА Протоны, ответственные за эффект Бора, высвобождаются в результате разрушения солевых мостиков, которым сопровождается связывание кислорода с Т-структурой; они отсоединяются от атомов азота остатков гистидина (146) в бета-цепях. Эти протоны сдвигают равновесие в сторону образования угольной кислоты, которая расщепляется карбоангидразой с образованием углекислого газа (рис.13). Наоборот, при высвобождении кислорода вновь формируется Т-структура с присущими ей солевыми мостиками, при образовании которых происходит присоединение протонов к остаткам гистидина в бета-цепях. Таким образом, в периферических тканях протоны благоприятствуют образованию солевых мостиков путем протонирования (по атому азота) концевых остатков гистидина в бета- субъединицах. Образование солевых мостиков форсирует освобождение кислорода из оксигенированной R-формы гемоглобина. Итак, повышение концентрации протонов способствует освобождению кислорода, а повышение концентрации кислорода стимулирует высвобождение протонов. Первый из этих эффектов проявляется в сдвиге кривой диссоциации кислорода вправо при повышении концентрации ионов водорода (протонов).[3] КОНЦЕНТРАЦИЯ ГЕМОГЛОБИНА Нормальная концентрация гемоглобина у взрослого человека от 80 до 115% (условных процентов=13,0-18,5 г%). За среднюю величину принимают 100% (=16 г%). Нормальные величины у мужчин приблизительно на 10% выше (90-115%, соответственно 14,5-18,5 г% гемоглобина), чем у женщин (80-100%, соответственно 13-16 г% гемоглобина). Нормальная концентрация гемоглобина 1у ребенка 0существено отличается от норм у взрослого. Эти особенности показаны на рис.14 и табл. 1. [таблица 1] Средняя концентрация гемоглобина в крови в периоды детского возраста. Максимальные колебания средних величин/-12% Возраст Первые 4 дня 2 1/2 мес 1 год 2 года 4 года 8 лет 12 лет Конц Hb 19,5 11,5 12,0 12,1 12,5 13,0 13,4 У детей в раннем возрасте нет различия между мужским и женским полом.[4] Гемоглобин в плазме крови Нормальная плазма содержит следы гемоглобина, не превышающие 10 мг%. При интравитальном гемолизе концентрация гемоглобина в плазме повышается. Умеренные повышения (до 25мг%) встречаются при иммунных гемолитических анемиях, анемии Кули, гемоглобинозе С, дрепаноцитозе и др. Сильные увеличения (свыше 100 мг%) встречаются при всех гемоглобинуриях. [5] СПОСОБЫ ИССЛЕДОВАНИЯ Было предложено много методов определения концентрации гемоглобина. Важнейшие группы методов следующие: 1. Колориметрические методы 0. Гемоглобин колориметрируют как оксигемоглобин или редуцированный гемоглобин или же сперва превращают его в цветные производные (солянокислый гематин, щелочной гемоглобин, метгемоглобин, карбоксигемоглобин, циангемоглобин, азид-метгемоглобин и пр.). Сюда можно отнести и первый метод для определения гемоглобина, предложенный Велькером в 1854 году и модифицированный Тальквистом, при котором цвет капли крови на фильтровальной бумаге сравнивают с серией цветных бумажных стандартов. На основании превращения гемоглобина в солянокислый гематин и связанных с этим изменений в электрической проводимости, Неллер предложил электронный метод определения концентрации гемоглобина. 2. Газометрические методы. Гемоглобин насыщают газом, например кислородом, окисью углерода (СО). По количеству поглощенного газа судят о количестве гемоглобина. Количество кислорода устанавливают прибором ван-Слайка, прибором Баркрофта или каким-нибудь другим аппаратом для определения кислорода. 3. Методы, основанные на определении железа в гемоглобиновой молекуле. Так как гемоглобиновая молекула содержит точно определенное количество железа (0,0347%), по его количеству устанавливается и количество гемоглобина.[6] МЕТГЕМОГЛОБИН Метгемоглобин - производное гемоглобина, в котором двухвалентный атом железа переходит в трехвалентный. При процессах обмена в эритроцитах всегда образуются известные количества метгемоглобина, который, однако, восстанавливается обратно в гемоглобин под воздействием фермента метгемоглобинредуктазы, так что в цельной крови здорового человека метгемоглобин не превышает 2% общего содержания гемоглобина (0,03-0,3 г%).[7] СУЛЬФОГЕМОГЛОБИН Химическая структура сульфогемоглобина не выяснена. Вероятно, две виниловые группы гемоглобина соединяются, посредством SО2-мостиков, с соседними метиновыми связями. В норме, сульфогемоглобина в крови нет. Он появляется при отравлениях соединениями сурьмы, фенацитином, бромом, сульфонамидами, нитратами (колодезная вода), серными соединениями и пр. Определение сульфогемоглобина в крови можно произвести спектроскопически. Сульфогемоглобиновый спектр не изменяется от прибавления сульфида аммония, но исчезает от прибавления Na2S2О4 и 2 мл 10% едкого натра, или нескольких капель 3% перекиси водорода. ТИПЫ ГЕМОГЛОБИНА Недавно еще считалось, что гемоглобин взрослого человека представляет собой одно единственное соединение. Известно было только то, что в эмбриональной жизни имеется особенный тип гемоглобина, называемый HbF, в 155 раз более устойчивый к n/12 натриевой щелочи, чем нормальный гемоглобин. В последнее время, благодаря работам Полинга и его сотрудников и др., выяснилось, что гемоглобин взрослого человека и при нормальных, и при патологических состояниях не представляет собой гомогенного химического соединения. Открыто было много нормальных и патологических типов гемоглобина, которые представили в новом свете обмен гемоглобина и указали пути для исследования патогенеза некоторых анемий. Установлено было, что при некоторых заболеваниях наблюдаются особые типы гемоглобина, характерные для данной анемии. Типы гемоглобина имеют большое значение не только для диагноза, но и перемежают вопрос о патогенезе анемии из чисто морфологической области в биохимическую. Анемии, вызванные появлением патологического типа гемоглобина, называются гемоглобинопатиями или гемоглобинозами. Выяснилось, что у человека имеются три основных типа ормального гемоглобина: эмбриональный U, фетальный - F и гемоглобин взрослого человека - А. HbU (назван по начальной букве слова uterus) встречается в эмбрионе между 7 и 12 неделями жизни, затем он исчезает и появляется фетальный гемоглобин, который после третьего месяца является основным гемоглобином плода. Вслед за этим появляется постепенно обыкновенный гемоглобин взрослого человека, называемый Hb A, по начальной букве английского слова "adult". Количество фетального гемоглобина постепенно уменьшается, так что в момент рождения 80% гемоглобина представляет собой Hb A и только 20% - HbF. После рождения фетальный гемоглобин продолжает убывать и к 2-3 году жизни составляет всего 1-2% (рис.15). Тоже количество фетального гемоглобина и у взрослого. Количество HbF, превышающее 2% считается патологическим для взрослого человека и для детей старше 3 лет. Кроме нормальных типов гемоглобина в настоящее время известно свыше 50 его патологических вариантов. Они сначала были названы латинскими буквами. Буква В в обозначениях типов гемоглобина отсутствует, т.к. ею обозначен первоначально Hb S. Вскоре выяснилось, что букв азбуки не хватит для обозначения всех патологических типов гемоглобина. Поэтому стали применять для этого имена пациентов, больниц, лабораторий, названия мест и округов. Самой удобной является номенклатура по структурной формуле (см. ниже). Как нормальные, так и патологические типы гемоглобина различаются не по структуре протопорфиринового кольца, а построению глобина. Разница может заключаться в изменении целых пар полипептидных цепей в гемоглобиновой молекуле, или при сохранении тех же полипептидных цепей, замещаются на определенном месте в первичной структуре одна аминокислота другой. Первая возможность встречается у гемоглобинов H, F, Бартс, А2 и U. Вместо нормальной структуры гемоглобина А - альфа-альфа/бета-бета, соответственно альфа2/бета2 , гемоглобин Н имеет структуру бета-бета-бета-бета, соответственно бета4, что значит, что по обе альфа-полипептидные цепи замещены новыми двумя бета-полипептидными цепями. У гемоглобинов F, Бартс и А2 появляются две новые цепи, обозначаемые гамма и дельта, а у гемоглобина U новая цепь, обозначаемая ипсилон. Структура HbF альфа-альфа/гамма-гамма, соответственно альфа2/гамма2, структура гемоглобина Бартс гамма-гамма-гамма-гамма, соотв. гамма4, структура HbА2 альфа-альфа/дельта-дельта, соответственно альфа2/гамма2, структура гемоглобина U - альфа-альфа/ипсилон-ипсилон, соответственно альфа2/ипсилон2. Патологические гемоглобины, которые состоят из четырех одинаковых полипептидных цепей, обозначают тетрамерами. Тетрамеры альфа4 и дельта4 до сих пор in vivo не наблюдались. Вторая возможность встречается у большинства типов гемоглобина. Так например единственная разница между HbS и HbA состоит в том, что на 6-ом месте в бета-полипептидной цепи вместо глутамина находится валин, единственная разница между HbI и HbA в том, что на 16-ом месте в альфа-полипептидной цепи лизин замещен аспарагиновой кислотой. На рис. 16 даны рад других подобных примеров. Когда аномалия состоит в замещении аминокислоты в альфа-полипептидной цепи, то говорят об альфа-аномалии, когда состоит в бета-полипептидной цепи - о бета-цепной аномалии, когда в гамма-полипептидной цепи - о гамма-цепной аномалии (патологические варианты HbF) и когда в дельта-цепи - о дельта-цепной аномалии (патологические варианты HbA2). При изучении гемоглобиновых типов имеет большое значение вопрос о структуре глобинов. С одной стороны, структура является самым верным способом отдифференцирования отдельных типов гемоглобина один от другого, с другой стороны создается возможность для составления строго научной номенклатуры последних. МЕТОДЫ ДИФФЕРЕНЦИРОВКИ ВИДОВ ГЕМОГЛОБИНА 1. Для разграничения отдельных типов человеческого гемоглобина пользуются электрофорезом на блоке крахмала, на крахмальном геле, на геле агара, на целлюлозно-ацетатных листах, на акриламидном геле, на карбоксиметилцеллюлозном геле, электрофорезом при высоком напряжении тока. 2. Вторым по значению методом, которым пользуются в настоящее время для дифференциации отдельных видов гемоглобина, является хроматография. Особенно хорошие результаты получается при употреблении в качестве адсорбирующего вещества ионообменной смолы амберлита и ионообменный декстрановый гель. На рис. 17 даны характеристики различных типов гемоглобина, полученных при применении амберлита при рН=6,0. 3. Для разграничения некоторых видов гемоглобина пользуются также их растворимостью в некоторых растворителях. Наиболее известным тестом этой группы является проба Итано для доказательства наличия HbS. При этой пробе нам служит то обстоятельство, что редуцированный HbS осаждается в 2,24 m буфере, в противоположность другим типам гемоглобина (рис. 18). Проба эта имеет значение в особенности для дифференцирования HbS и HbD, потому что, как видно из рис. 18, HbS и HbD обладают одинаковой электрофоретической и хроматографической подвижностью. 4. Для отличия HbA от HbF пользуются, как было подчеркнуто выше, устойчивостью при денатурации растворами натриевой щелочи. Это известный в истории метод, которым Кербер в 1886 году дифференцировал HbA и HbF. 5. Гемоглобины группы F (HbF, Hb Феллас, Hb Александра и Hb Бартс) отличается от других гемоглобиновых типов и по своей характерной триптофановой полосе при 289,8 нм ультрафиолетового спектра. Гемоглобины, обладающие группой М, не имеют абсорбционной полосы при длине волны 630 нм, но зато показывают увеличенную абсорбцию при 600 нм. 6. "Отпечатковый метод" .Дело касается важнейшего метода установления "первичной структуры" гемоглобина при различных гемоглобиновых типах. Исследуемый гемоглобин гидролизуют трипсином, при чем полипептидные цепи глобиновой молекулы распадаются на большое число пептидов. Пептидную смесь подвергают электрохроматографии на бумаге, т.е. в одном направлении проводится электрофоретическое, в другом хроматографическое разделение. Получаются характерные для отдельных типов гемоглобинов электрохроматограммы, по которым их можно точно различить (рис. 19). Определение аминокислотного состава отдельных пептидов дает возможность первичную структуру глобина соответствующего гемоглобинового типа. Делая аналогию с соответствующей по сложности и точности криминалистической техникой для изучения отпечатков пальцев рук, он был назван "пальцеотпечатковым" ("fingerprint") методом. 7. Для определения состава полипептидных цепей в каком-нибудь гемоглобиновом типе можно воспользоваться и так называемым "рекомбинационным" или "гибридизационным" методом. Если смешать известный и неизвестный гемоглобин при рН 4,3, они диссоциируют полумолекулами, состоящими из соответствующих пар полипептидных цепей. После нейтрализации раствора полипептидные пары снова комбинируются в целые гемоглобиновые молекулы, при чем могут получится и новые "гибридные" гемоглобиновые молекулы. Их идентифицирование электрофоретическим способом или хроматографией позволит сделать заключение о полипептидной структуре неизвестного гемоглобинового типа. Этот метод предназначен также преимущественно для научных исследовательских целей. 8. Иммунологические методы. 9. Кроме вышеуказанных методов при дифференциации отдельных типов гемоглобина пользуются также различиями в кристаллическом строении, изоэлектрической точке и т.д. 10. Разработаны также методы цитологического определения типа гемоглобина в эритроцитах на мазке крови. Так наличие HbF в эритроцитах можно доказать путем обработки кровяного мазка лимоннокислой буферной смесью с рН 3,2-3,6. При этих условиях HbA извлекается и эритроциты, в которых он преобладал, остаются только в виде эритроцитных теней, тогда как HbF сохраняется и эритроциты, содержащие преимущественно этот тип гемоглобина, сохраняют свое содержание.[8] ГЕМОГЛОБИН ПРИ СЕРПОВИДНОКЛЕТОЧНОЙ АНЕМИИ В гемоглобине S остаток Glu А2(6)бета замещен на Val. Остаток А2 (Glu или Val) располагается на поверхности молекулы гемоглобина и контактирует в водой, и замещение полярного остатка Glu на неполярный Val приводит к появлению на поверхности бета-субъеденицы "липкого участка". Этот липкий участок присутствует как в оксигенированном, так и в дезоксигенированном гемоглобине S (в гемоглобине А отсутствует). На поверхности дезоксигенированного гемоглобина существует комплементарный участок, способный прочно связываться с липким участком бета-субъединицы, тогда как в оксигенированном гемоглобине этот участок маскируется другими группами (рис. 20). Когда гемоглобин S переходит в дезоксигенированное состояние, его липкий участок связывается с комплементарным участком на другой молекуле дезоксигенированного гемоглобина. Происходит полимеризация дезоксигемоглобина S и его осаждение в виде длинных волокон. Волокна дезоксигемоглобина S механически деформируют эритроцит, предавая ему серповидную форму, что приводит к лизису клеток и множеству вторичных клинических проявлений. Таким образом, если бы можно было можно поддерживать гемоглобин S в оксигенированном состоянии или по крайней мере свести к минимуму концентрацию дезоксигенированного гемоглобина S, то нам удалось бы предотвратить полимеризацию дезоксигенированного гемоглобина S и образование "серповидных" клеток. Ясно, что полимеризации подвержена Т-форма гемоглобина S. Интересно отметить (хотя в практическом плане это малосущественно), что ферри-ион метгемоглобина А остается в плоскости порфиринового кольца и тем самым стабилизирует R-форму гемоглобина. То же относится и к гемоглобину при серповидноклеточной анемии: гемоглобин S в ферри-состоянии (метгемоглобин S) не подвержен полимеризации, поскольку он стабилизирован в R-форме. В дезоксигемоглобине А тоже имеется рецепторный участок, способный взаимодействовать с липким участком оксигенированного или дезоксигенированного гемоглобина S (рис.20), но присоединения "липкого" гемоглобина S к к дезоксигемоглобину А недостаточно для образования полимера, поскольку сам дезоксигемоглобин А липкого участка не содержит и не может связывать следующую молекулу гемоглобина. Следовательно, связывание дезоксигемоглобина А с R- или Т-формой гемоглобина S перекрывает полимеризацию. В результате полимеризации дезоксигемоглобина S образуются спиральные фибрилярные структуры. При этом каждая молекула гемоглобина контактирует с четырьмя соседними молекулами (рис. 21). Образование подобных трубчатых волокон ответственно за механические нарушения в содержащем их эритроците: он приобретает серповидную форму (рис. 22), становится подверженным лизису в момент прохождения им щелей в синусоидах селезенки. ТАЛАССЕМИИ Другая важная группа нарушений, связанных с аномалиями гемоглобина - талассемии. Для них характерна пониженная скорость синтеза альфа-цепей гемоглобина (альфа-талассемия) или бета-цепей (бета-талассемия). Это приводит к анемии, которая может принимать очень тяжелую форму. В последние годы достигнут ощутимый прогресс в выяснении молекулярных механизмов, ответственных за развитие талассемии.[9] СПИСОК ЛИТЕРАТУРЫ I. Основная литература: 1. Р.Марри, Д.Греннер, П.Мейес, В.Родуэлл, Биохимия человека, том 1, "Мир", Москва 1993г., стр.52 2. И.Тодоров, Клинические лабораторные исследования в педиатрии, "Медицина и физкультура",София 1968г., стр. 278-281 3. Р.Марри, Д.Греннер, П.Мейес, В.Родуэлл, Биохимия человека, том 1, "Мир", Москва 1993г., стр.56-59 4. И.Тодоров, Клинические лабораторные исследования в педиатрии, "Медицина и физкультура",София 1968г., стр.283-284 5. тоже стр.293 6. тоже стр.285-286 7. тоже стр.293-304 8. Р.Марри, Д.Греннер, П.Мейес, В.Родуэлл, Биохимия человека, том 1, "Мир", Москва 1993 г.6 стр. 60-62 II. Дополнительная литература Dean J., Schechter A.N. Sickle-cell anemia: Molekular and lubar basis of therapeutic approaches. (3 parts), N.E.Med., 1978, 299, 752, 804, 863. Klotz I.M., Haney D.N., King L.C. Ritional approaches chemotherapy: Antisickling agents, Sience, 1981, 219 |